
Genetic Loci Underlying Awn Morphology in Barley



Abstract
1. Awnness or Awnlessness
2. Biological Function and Anatomy of Awns
3. Morphological Diversity of Barley Awns
4. Cloned Awnness Genes in Barley
5. Mapped Genetic Loci for Awns in Barley
6. Interactions of Awnness Genes
7. Pleiotropism of Awn Genes
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ntakirutimana, F.; Xie, W. Morphological and genetic mechanisms underlying awn development in monocotyledonous grasses. Genes 2019, 10, 573. [Google Scholar] [CrossRef]
- Saisho, D.; Takeda, K. Barley: Emergence as a new research material of crop science. Plant Cell Physiol. 2011, 52, 724–727. [Google Scholar] [CrossRef] [PubMed]
- Druka, A.; Franckowiak, J.; Lundqvist, U.; Bonar, N.; Alexander, J.; Houston, K.; Radovic, S.; Shahinnia, F.; Vendramin, V.; Morgante, M.; et al. Genetic dissection of barley morphology and development. Plant Physiol. 2011, 155, 617–627. [Google Scholar] [CrossRef] [PubMed]
- The International Barley Genome Sequencing Consortium. A physical, genetic and functional sequence assembly of the barley genome. Nature 2012, 491, 711–717. [Google Scholar] [CrossRef]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. Chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Monat, C.; Padmarasu, S.; Lux, T.; Wicker, T.; Gundlach, H.; Himmelbach, A.; Ens, J.; Li, C.; Muehlbauer, G.J.; Schulman, A.H.; et al. TRITEX: Chromosome-scale sequence assembly of Triticeae genomes with open-source tools. Genome Biol. 2019, 20, 284. [Google Scholar] [CrossRef]
- van Esse, G.W.; Walla, A.; Finke, A.; Koornneef, M.; Pecinka, A.; von Korff, M. Six-rowed spike3 (VRS3) is a histone demethylase that controls lateral spikelet development in barley. Plant Physiol. 2017, 174, 2397–2408. [Google Scholar] [CrossRef]
- Bull, H.; Casao, M.C.; Zwirek, M.; Flavell, A.J.; Thomas, W.T.B.; Guo, W.; Zhang, R.; Rapazote-Flores, P.; Kyriakidis, S.; Russell, J.; et al. Barley SIX-ROWED SPIKE3 encodes a putative Jumonji C-type H3K9me2/me3 demethylase that represses lateral spikelet fertility. Nat. Commun. 2017, 8, 936. [Google Scholar] [CrossRef]
- Schrager-Lavelle, A.; Klein, H.; Fisher, A.; Bartlett, M. Grass flowers: An untapped resource for floral evo-devo. J. Syst. Evol. 2017, 55, 525–541. [Google Scholar] [CrossRef]
- Lombardo, F.; Hitoshi, Y. Interpreting lemma and palea homologies: A point of view from rice floral mutants. Front. Plant Sci. 2015, 6, 61. [Google Scholar] [CrossRef]
- Guo, Z.; Schnurbusch, T. Costs and benefits of awns. J. Exp. Bot. 2016, 67, 2533–2535. [Google Scholar] [CrossRef]
- Yuo, T.; Yamashita, Y.; Kanamori, H.; Matsumoto, T.; Lundqvist, U.; Sato, K.; Ichii, M.; Jobling, S.A.; Taketa, S. A SHORT INTERNODES (SHI) family transcription factor gene regulates awn elongation and pistil morphology in barley. J. Exp. Bot. 2012, 63, 5223–5232. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Bonnett, D.G.; Reynolds, M.P. Awns reduce grain number to increase grain size and harvestable yield in irrigated and rainfed spring wheat. J. Exp. Bot. 2016, 67, 2573–2586. [Google Scholar] [CrossRef]
- Terzi, V.; Tumino, G.; Pagani, D.; Rizza, F.; Ghizzoni, R.; Morcia, C.; Stanca, A.M. Barley developmental mutants: The high road to understand the cereal spike morphology. Diversity 2017, 9, 21. [Google Scholar] [CrossRef]
- Franckowiak, J.D.; Kleinhofs, A.; Lundqvist, U. Descriptions of barley genetic stocks for 2016. Barley Genet. Newsl. 2016, 46, 1–47. [Google Scholar]
- Schulte, D.; Close, T.J.; Graner, A.; Langridge, P.; Matsumoto, T.; Muehlbauer, G.; Sato, K.; Schulman, A.H.; Waugh, R.; Wise, R.P.; et al. The International Barley Sequencing Consortium–at the threshold of efficient access to the barley genome. Plant Physiol. 2009, 149, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Stanca, A.M.; Tumino, G.; Pagani, D.; Rizza, F.; Alberici, R.; Lundqvist, U.; Morcia, C.; Tondelli, A.; Terzi, V. The “Italian” barley genetics mutant collection: Conservation, development of new mutants and use. In Advance in Barley Sciences; Zhang, G., Li, C., Liu, X., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 30–35. [Google Scholar]
- Stebbins, G.L.; Yagil, E. The morphogenetic effects of the hooded gene in barley. I. The course of development in hooded and awned genotypes. Genetics 1966, 54, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, C.; Faccioli, P.; Terzi, V.; Stanca, A.M.; Cerioli, S.; Castiglioni, P.; Fink, R.; Capone, R.; Müller, K.J.; Bossinger, G.; et al. Genetics of mutations affecting the development of a barley floral bract. Genetics 2000, 154, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Xu, T. Classification and identification of varieties of Chinese cultivated barley. Sci. Agric. Sin. 1982, 6, 39–47. [Google Scholar]
- Sun, L. Varieties of cultivated barley and their distribution in China. Sci. Agric. Sin. 1988, 21, 25–31. [Google Scholar]
- Müller, K.J.; Romano, N.; Gerstner, O.; Garcia-Maroto, F.; Pozzi, C.; Salamini, F.; Rohde, W. The barley hooded mutation caused by a duplication in a homeobox gene intron. Nature 1995, 374, 727–730. [Google Scholar] [CrossRef]
- Williams-Carrier, R.E.; Lie, Y.S.; Hake, S.; Lemaux, P.G. Ectopic expression of the maize kn1 gene phenocopies the Hooded mutant of barley. Development 1997, 124, 3737–3745. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, C.; di Pietro, D.; Halas, G.; Roig, C.; Salamini, F. Integration of a barley (H. vulgare) molecular linkage map with the position of genetic loci hosting 29 developmental mutants. Heredity 2003, 90, 390–396. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Komatsuda, T.; Pourkheirandish, M.; He, C.F.; Azhaguvel, P.; Kanamori, H.; Perovic, D.; Stein, N.; Graner, A.; Wicker, T.; Tagiri, A.; et al. Six-rowed barley originated from a mutation in a homeodomain leucine zipper I-class homeobox gene. Proc. Natl. Acad. Sci. USA 2007, 104, 1424–1429. [Google Scholar] [CrossRef]
- Leonard, W.H. Inheritance of reduced lateral spikelet appendages in the Nudihaxtoni variety of barley. J. Am. Soc. Agron. 1942, 34, 211–221. [Google Scholar] [CrossRef]
- Taketa, S.; Yuo, T.; Sakurai, Y.; Miyake, S.; Ichii, M. Molecular mapping of the short awn 2 (lks2) and dense spike 1 (dsp1) genes on barley chromosome 7H. Breed. Sci. 2011, 61, 80–85. [Google Scholar] [CrossRef]
- Robertson, D.W.; Tsuchiya, T.; Haus, T.E. Description of genetic stocks in the Barley Genetic Stock Center at Fort Collins, Colorado. BGS Reduced lateral spikelet appendage on the lemma lr. Barley Genet. Newsl. 1971, 1, 92–193. [Google Scholar]
- Takahashi, R.; Hayashi, J. Studies on classification and inheritance of barley varieties having awnless or shorter-awned lateral spikelets (Bozu barley). II. Mode of inheritance of spike characteristics of Bozu barley-1. Nogaku Kenkyu 1982, 60, 25–37. [Google Scholar]
- Kuusk, S.; Sohlberg, J.J.; Eklund, D.M.; Sundberg, E. Functionally redundant SHI family genes regulate Arabidopsis gynoecium development in a dose-dependent manner. Plant J. 2006, 47, 99–111. [Google Scholar] [CrossRef]
- Takahashi, R. Genetic features of East Asian barleys. In Barley Genetics V; Yasuda, S., Konishi, T., Eds.; Sanyo Press Co.: Okayama, Japan, 1987; pp. 7–20. [Google Scholar]
- Woodward, R.W.; Rasmussen, D.C. Hood and awn development in barley determined by two gene pairs. Agron. J. 1957, 49, 92–94. [Google Scholar] [CrossRef]
- Gruszka, D.; Szarejko, I.; Maluszynski, M. New allele of HvBRI1 gene encoding brassinosteroid receptor in barley. J. Appl. Genet. 2011, 52, 257–268. [Google Scholar] [CrossRef]
- Chono, M.; Honda, I.; Zeniya, H.; Yoneyama, K.; Saisho, D.; Takeda, K.; Takatsuto, S.; Hoshino, T.; Watanabe, Y. A semidwarf phenotype of barley uzu results from a nucleotide substitution in the gene encoding a putative brassinosteroid receptor. Plant Physiol. 2003, 133, 1209–1219. [Google Scholar] [CrossRef]
- Klein, H.; Xiao, Y.; Conklin, P.A.; Govindarajulu, R.; Kelly, J.A.; Scanlon, M.J.; Whipple, C.J.; Bartlett, M. Bulked-segregant analysis coupled to whole genome sequencing (BSA-Seq) for rapid gene cloning in maize. G3 Genes Genomes Genet. 2018, 8, 3583–3592. [Google Scholar] [CrossRef]
- Burnham, C.R.; Hagberg, A. Cytogenetic notes on chromosomal interchanges in barley. Hereditas 1956, 42, 467–482. [Google Scholar] [CrossRef]
- Gustafsson, Å.; Hagberg, A.; Lundqvist, U.; Persson, G. A proposed system of symbols for the collection of barley mutants at Svalöv. Hereditas 1969, 62, 409–414. [Google Scholar] [CrossRef]
- Graner, A.; Jahoor, A.; Schondelmaier, J.; Siedler, H.; Pillen, K.; Fischbeck, G.; Wenzel, G.; Herrmann, R.G. Construction of an RFLP map of barley. Theor. Appl. Genet. 1991, 83, 250–256. [Google Scholar] [CrossRef]
- Rossini, L.; Vecchietti, A.; Nicoloso, L.; Stein, N.; Franzago, S.; Salamini, F.; Pozzi, C. Candidate genes for barley mutants involved in plant architecture: An in silico approach. Theor. Appl. Genet. 2006, 112, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Konishi, T.; Hayashi, J.; Myriya, I.; Takahashi, R. Inheritance and linkage studies in barley. VII. Location of six new mutant genes on chromosome 3. Ber. Ohara Inst. Landwirtsch. Biol. Okayama Univ. 1984, 18, 251–264. [Google Scholar]
- Kucera, J.; Lundqvist, U.; Gustafsson, A. Induction of breviaristatum mutants in barley. Hereditas 1975, 80, 263–278. [Google Scholar] [CrossRef]
- Persson, G. An attempt to find suitable genetic markers for the dense ear loci in barley II. Hereditas 1969, 63, 1–28. [Google Scholar] [CrossRef]
- Robertson, D.W.; Immer, F.R.; WLebe, G.A.; Stevens, H.L. The location of two genes for mature plant characters in barley in linkage group I. J. Am. Soc. Agron. 1944, 36, 66–72. [Google Scholar] [CrossRef]
- Huang, B.; Huang, D.; Hong, Z.; Owie, S.O.; Wu, W. Genetic analysis reveals four interacting loci underlying awn trait diversity in barley (Hordeum vulgare). Sci. Rep. 2020, 10, 12535. [Google Scholar] [CrossRef] [PubMed]
- Kleinhofs, A.; Franckowiak, J.D. BGS 724, Short awn 6, lks6. Barley Genet. Newsl. 2013, 43, 219. [Google Scholar]
- Yoshioka, M.; Iehisa, J.C.M.; Ohno, R.; Kimura, T.; Enoki, H.; Nishimura, S.; Nasuda, S.; Takumi, S. Three dominant awnless genes in common wheat: Fine mapping, interaction and contribution to diversity in awn shape and length. PLoS ONE 2017, 12, e0176148. [Google Scholar] [CrossRef] [PubMed]
- Mulki, M.A.; Bi, X.; von Korff, M. FLOWERING LOCUS T3 controls spikelet initiation but not floral development. Plant Physiol. 2018, 178, 1170–1186. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Zheng, Q.; Melchkart, T.; Bekkaoui, Y.; Konkin, D.J.F.; Kagale, S.; Martucci, M.; You, F.M.; Clarke, M.; Adamski, N.M.; et al. Dominant inhibition of awn development by a putative zinc-finger transcriptional repressor expressed at the B1 locus in wheat. New Phytol. 2020, 225, 340–355. [Google Scholar] [CrossRef]
- DeWitt, N.; Guedira, M.; Lauer, E.; Sarinelli, M.; Tyagi, P.; Fu, D.; Hao, Q.; Murphy, J.P.; Marshall, D.; Akhunova, A.; et al. Sequence-based mapping identifies a candidate transcription repressor underlying awn suppression at the B1 locus in wheat. New Phytol. 2020, 225, 326–339. [Google Scholar] [CrossRef]
- Würschum, T.; Jähne, F.; Phillips, A.L.; Langer, S.M.; Longin, C.F.H.; Tucker, M.R.; Leiser, W.L. Misexpression of a transcriptional repressor candidate provides a molecular mechanism for the suppression of awns by Tipped 1 in wheat. J. Exp. Bot. 2020, 71, 3428–3436. [Google Scholar] [CrossRef]
- Wang, D.; Yu, K.; Jin, D.; Sun, L.; Chu, J.; Wu, W.; Xin, P.; Gregova, E.; Li, X.; Sun, J.; et al. Natural variations in the promoter of Awn Length Inhibitor 1 (ALI-1) are associated with awn elongation and grain length in common wheat. Plant J. 2020, 101, 1075–1090. [Google Scholar] [CrossRef]
- Niu, J.; Zheng, S.; Shi, X.; Si, Y.; Tian, S.; He, Y.; Ling, H. Fine mapping and characterization of the awn inhibitor B1 locus in common wheat (Triticum aestivum L.). Crop J. 2020, 8, 613–622. [Google Scholar] [CrossRef]
- Myler, J.L. Awn inheritance in barley. J. Agric. Res. 1942, 65, 405–412. [Google Scholar]
- Litzenberger, S.C.; Green, J.M. Inheritance of awns in barley. Agron. J. 1951, 43, 117–123. [Google Scholar] [CrossRef]
- Liller, C.B.; Walla, A.; Boer, M.P. Fine mapping of a major QTL for awn length in barley using a multiparent mapping population. Theor. Appl. Genet. 2017, 130, 269–281. [Google Scholar] [CrossRef]
- Sun, D.F.; Ren, W.B.; Sun, G.L.; Peng, J.H. Molecular diversity and association mapping of quantitative traits in Tibetan wild and worldwide originated barley (Hordeum vulgare L.) germplasm. Euphytica 2011, 178, 31–43. [Google Scholar] [CrossRef]
- Chen, G.D.; Li, H.B.; Zheng, Z.; Wei, Y.M.; Zheng, Y.L.; McIntyre, C.L.; Zhou, M.X.; Liu, C.J. Characterization of a QTL affecting spike morphology on the long arm of chromosome 3H in barley (Hordeum vulgare L.) based on near isogenic lines and a NIL-derived population. Theor. Appl. Genet. 2012, 125, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Yang, J.M.; McNeil, D.L.; Zho, M.X. Identification and molecular mapping of a dwarfing gene in barley (Hordeum vulgare L.) and its correlation with other agronomic traits. Euphytica 2010, 175, 331–342. [Google Scholar] [CrossRef]
- Gyenis, L.; Yun, S.J.; Smith, K.P.; Steffenson, B.J.; Bossolini, E.; Sanguineti, M.C.; Muehlbauer, G.J. Genetic architecture of quantitative trait loci associated with morphological and agronomic trait differences in a wild by cultivated barley cross. Genome 2007, 50, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Roig, C.; Pozzi, C.; Santi, L.; Müller, J.; Wang, Y.; Stile, M.R.; Rossini, L.; Stanca, M.; Salamini, F. Genetics of barley hooded suppression. Genetics 2004, 167, 439–448. [Google Scholar] [CrossRef]
- Gustaffson, A. Mutations in agricultural plants. Hereditas 1947, 33, 1–100. [Google Scholar] [CrossRef]
- Dockter, C.; Gruszka, D.; Braumann, I.; Druka, A.; Druka, I.; Franckowiak, J.; Gough, S.P.; Janeczko, A.; Kurowska, M.; Lundqvist, J.; et al. Induced variations in brassinosteroid genes define barley height and sturdiness, and expand the green revolution genetic toolkit. Plant Physiol. 2014, 166, 1912–1927. [Google Scholar] [CrossRef]
- Huang, B.; Wu, W.; Hong, Z. Genetic Interactions of Awnness Genes in Barley. Genes 2021, 12, 606. [Google Scholar] [CrossRef]
- Franckowiak, J.D. Revised linkage maps for morphological markers in barley, Hordeum vulgare. Barley Genet. Newsl. 1997, 26, 9–21. [Google Scholar]
- Sameri, M.; Takeda, K.; Komatsuda, T. Quantitative trait loci controlling agronomic traits in recombinant inbred lines from a cross of oriental- and occidental-type barley cultivars. Breed. Sci. 2006, 56, 243–252. [Google Scholar] [CrossRef][Green Version]
- Tsuchiya, T.; Singh, R.J. Another case of paracentric inversion in a genetic stock, Engleawnless. Barley Genet. Newsl. 1972, 2, 110–111. [Google Scholar]
- Ramage, R.T. Alleles at the short-awned loci lk2 on chromosome 1 and lk5 on chromosome 4. Barley Genet. Newsl. 1986, 16, 22–23. [Google Scholar]
- Kjack, J.L.; Witters, R.E. Physiological activity of awns in isolines of Atlas barley 1. Crop Sci. 1974, 14, 243–248. [Google Scholar]
- Maydup, M.L.; Antonietta, M.; Graciano, C.; Guiamet, J.J.; Tambussi, E.A. The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: Responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. Field Crop. Res. 2014, 167, 102–111. [Google Scholar] [CrossRef]
- Qualset, C.Q.; Schaller, C.W.; Williams, J.C. Performance of isogenic lines of barley as influenced by awn length, linkage blocks, and environment. Crop Sci. 1965, 5, 489–494. [Google Scholar] [CrossRef]
- Haas, M.; Schreiber, M.; Mascher, M. Domestication and crop evolution of wheat and barley: Genes, genomics, and future directions. J. Integr. Plant Biol. 2019, 61, 204–225. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
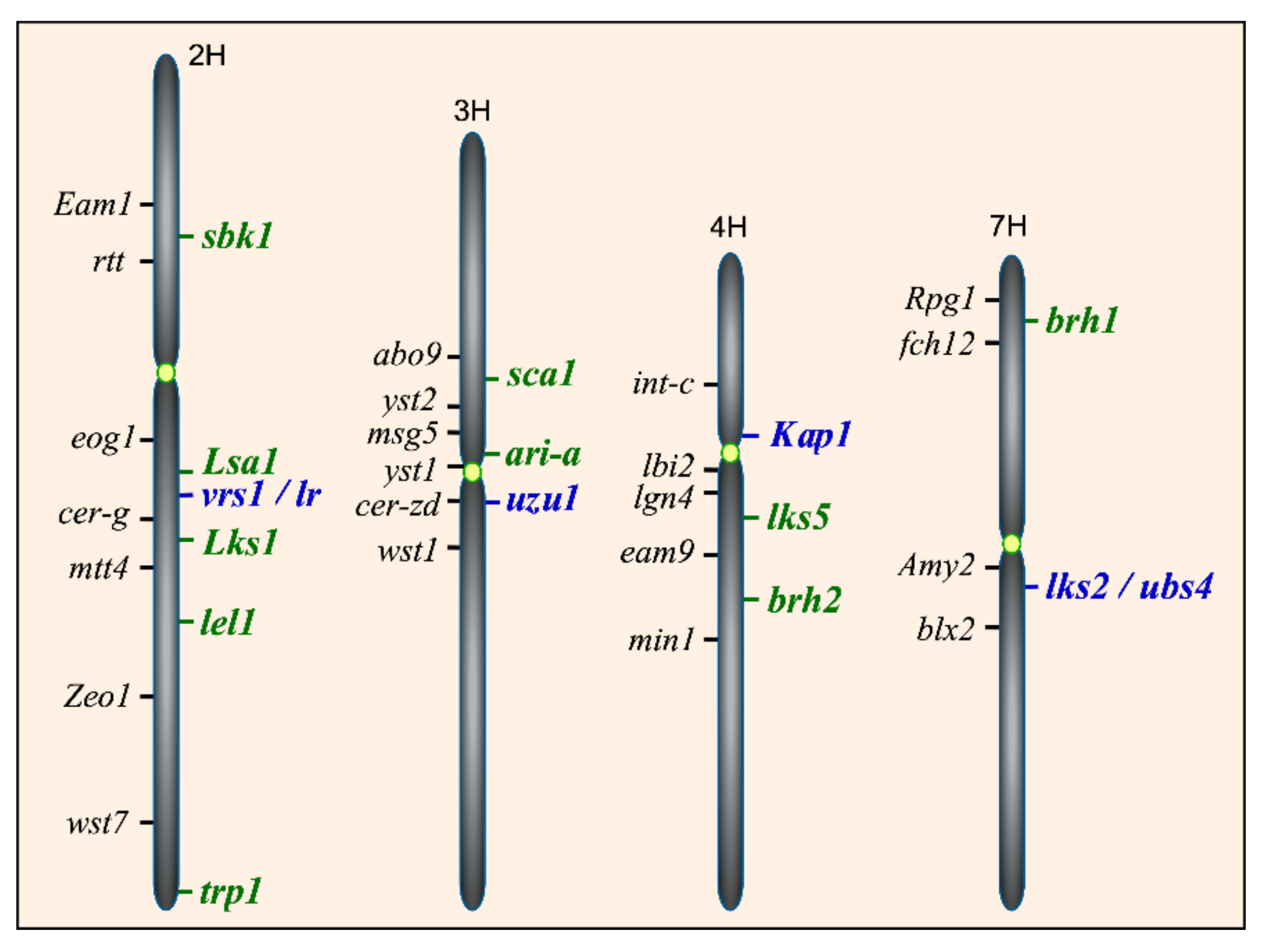{kind=link}
| Gene Names | Kap1 (hooded lemma 1) | vrs1 (six-rowed spike 1) | lks2 (short awn 2) |
| Other Names | K, HvKnox3 | lr, HvHox1 | lk2, lk4, ari-d, ubs4 |
| Awn Phenotype | Hooded lemma | Reduced lateral spikelet appendage (lr) | Short awn |
| Chromosome | 4HS | 2HL | 7HL |
| Cloning Method | Homologous cloning | Positional cloning | Positional cloning |
| Mutation Types | Insertion in intron 4 | Missense, nonsense, and splicing site changes | Missense mutation |
| Protein Length a | 364 aa | 222 aa | 344 aa |
| Protein Type | KNOX family transcription factor | HD-ZIP family transcription factor | SRS family transcription factor |
| Orthologs | Rice OSH15 and maize KNOTTED-1 | OsHox12 and OsHox14 in rice | Os06g0712600 in rice |
| References | [22,24] | [25,26] | [12,27] |
| Locus | Other Names | Awn Phenotypes | Chr a | Ref. b |
|---|---|---|---|---|
| sbk1 | sk, cal-a | Subjacent hood | 2HS | [19,37] |
| lel1 | lel | Leafy lemma | 2HL | [3,38] |
| trp1 | tr | Triple awned lemma | 2HL | [24,39] |
| sca1 | sca | Short, crooked awn | 3HS | [40] |
| lks5 | lk5, ari-c, lel2 | Short awn | 4HS | [27,39] |
| ari-a | lk7 | Short awn | 3HS | [41,42] |
| lks6 | Lks.q | Short awn | ND c | [3] |
| Lks1 | Lk | Awnless | 2HL | [43] |
| Lsa1 | Lsa | Awnless on lateral rows | 2H | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, B.; Wu, W.; Hong, Z. Genetic Loci Underlying Awn Morphology in Barley. Genes 2021, 12, 1613. https://doi.org/10.3390/genes12101613
Huang B, Wu W, Hong Z. Genetic Loci Underlying Awn Morphology in Barley. Genes. 2021; 12(10):1613. https://doi.org/10.3390/genes12101613
Chicago/Turabian StyleHuang, Biguang, Weiren Wu, and Zonglie Hong. 2021. "Genetic Loci Underlying Awn Morphology in Barley" Genes 12, no. 10: 1613. https://doi.org/10.3390/genes12101613
APA StyleHuang, B., Wu, W., & Hong, Z. (2021). Genetic Loci Underlying Awn Morphology in Barley. Genes, 12(10), 1613. https://doi.org/10.3390/genes12101613