
Context Fear Conditioning in Down Syndrome Mouse Models: Effects of Trisomic Gene Content, Age, Sex and Genetic Background

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Context Fear Conditioning
2.3. Statistics
3. Results
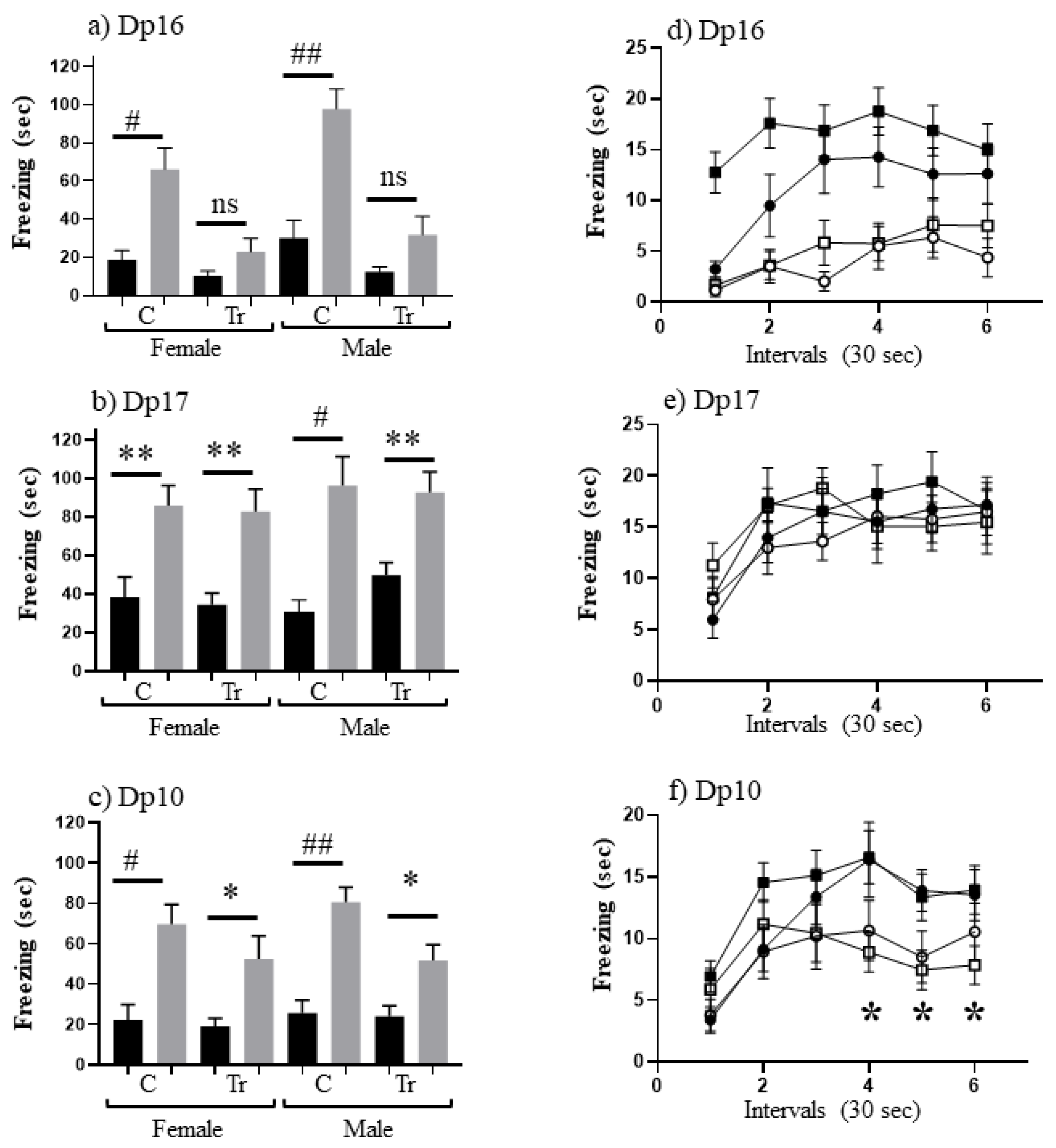
3.1. Performance in CFC at 3 Months of Age (Experiment 1)
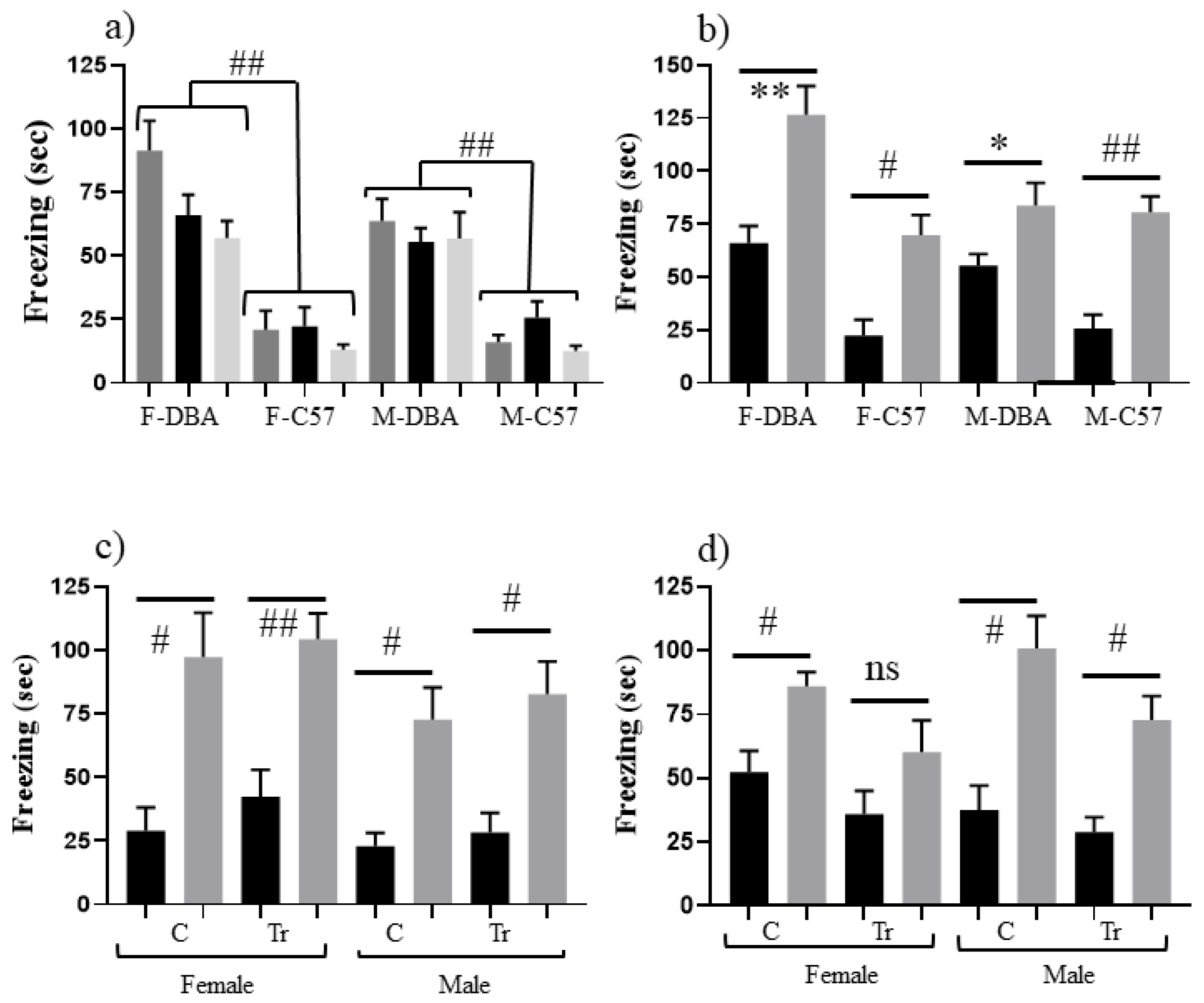
3.2. Effects of Genomic Background on CFC Performance (Experiment 2)
3.3. Performance in CFC at ~18 Months of Age (Experiment 3)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapman, R.S.; Hesketh, L.J. Behavioral phenotype of individuals with Down syndrome. Ment. Retard. Dev. Disabil. Res. Rev. 2000, 6, 84–95. [Google Scholar] [CrossRef]
- Silverman, W. Down syndrome: Cognitive phenotype. Ment. Retard. Dev. Disabil. Res. Rev. 2007, 13, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Improved National Prevalence Estimates for 18 Major Birth Defects. In Morbidity and Mortality Weekly Report; CDC: Atlanta, GA, USA, 2006; Volume 54, pp. 6–12. [Google Scholar]
- Irving, C.; Basu, A.; Richmond, S.; Burn, J.; Wren, C. Twenty-year trends in prevalence and survival of Down syndrome. Eur. J. Hum. Genet. 2008, 16, 1336–1340. [Google Scholar] [CrossRef] [PubMed]
- Parker, S.; Mai, C.T.; Canfield, M.A.; Rickard, R.; Wang, Y.; Meyer, R.E.; Anderson, P.; Mason, C.; Collins, J.S.; Kirby, R.S.; et al. Updated national birth prevalence estimates for selected birth defects in the United States, 2004–2006. Birth Defects Res. Part A Clin. Mol. Teratol. 2010, 88, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Glasson, E.J.; Sullivan, S.G.; Hussain, R.; Petterson, B.A.; Montgomery, P.D.; Bittles, A.H. The changing survival profile of people with Down’s syndrome: Implications for genetic counselling. Clin. Genet. 2002, 62, 390–393. [Google Scholar] [CrossRef]
- Bittles, A.H.; Glasson, E.J. Clinical, social, and ethical implications of changing life expectancy in Down syndrome. Dev. Med. Child. Neurol. 2007, 46, 282–286. [Google Scholar] [CrossRef]
- Bittles, A.H.; Bower, C.; Hussain, R.; Glasson, E.J. The four ages of Down syndrome. Eur. J. Public Heal. 2006, 17, 221–225. [Google Scholar] [CrossRef]
- Kucik, J.E.; Shin, M.; Siffel, C.; Marengo, L.; Correa, A. Congenital Anomaly Multistate Prevalence and Survival Collaborative Trends in survival among children with Down syndrome in 10 regions of the United States. Pediatrics 2013, 131, e27–e36. [Google Scholar] [CrossRef]
- Glasson, E.J.; Jacques, A.; Wong, K.; Bourke, J.; Leonard, H. Improved Survival in Down Syndrome over the Last 60 Years and the Impact of Perinatal Factors in Recent Decades. J. Pediatr. 2016, 169, 214–220. [Google Scholar] [CrossRef]
- De Graaf, G.; Buckley, F.; Skotko, B.G. Estimation of the number of people with Down syndrome in the United States. Genet. Med. 2017, 19, 439–447. [Google Scholar] [CrossRef]
- Hart, S.J.; Visootsak, J.; Tamburri, P.; Phuong, P.; Baumer, N.; Hernandez, M.-C.; Skotko, B.G.; Ochoa-Lubinoff, C.; D’Ardhuy, X.L.; Kishnani, P.S.; et al. Pharmacological interventions to improve cognition and adaptive functioning in Down syndrome: Strides to date. Am. J. Med. Genet. Part A 2017, 173, 3029–3041. [Google Scholar] [CrossRef] [PubMed]
- Antonarakis, S.E.; Skotko, B.G.; Rafii, M.S.; Strydom, A.; Pape, S.E.; Bianchi, D.W.; Sherman, S.L.; Reeves, R.H. Down syndrome. Nat. Rev. Dis. Primers 2020, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- de la Torre, R.; de Sola, S.; Hernandez, G.; Farré, M.; Pujol, J.; Rodriguez, J.; Espadaler, J.M.; Langohr, K.; Cuenca-Royo, A.; Principe, A. TESDAD study group. Safety and efficacy of cognitive training plus epigallocatechin-3-gallate in young adults with Down’s syndrome (TESDAD): A double-blind, randomised, placebo-controlled, phase 2 trial. Lancet Neurol. 2016, 15, 801–810. [Google Scholar] [CrossRef]
- Boada, R.; Hutaff-Lee, C.; Schrader, A.; Weitzenkamp, D.; Benke, T.A.; Goldson, E.J.; Costa, A.C.S. Antagonism of NMDA receptors as a potential treatment for Down syndrome: A pilot randomized controlled trial. Transl. Psychiatry 2012, 2, e141. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, K.J. Pharmacological approaches to improving cognitive function in Down syndrome: Current status and con-siderations. Drug Des. Dev. Ther. 2014, 9, 103–125. [Google Scholar] [CrossRef]
- Rahmani, Z.; Blouin, J.L.; Creau-Goldberg, N.; Watkins, P.C.; Mattei, J.F.; Poissonnier, M.; Prieur, M.; Chettouh, Z.; Nicole, A.; Aurias, A. Critical role of the D21S55 region on chromosome 21 in the pathogenesis of Down syndrome. Proc. Natl. Acad. Sci. USA 1989, 86, 5958–5962. [Google Scholar] [CrossRef]
- Delabar, J.-M.; Theophile, D.; Rahmani, Z.; Chettouh, Z.; Blouin, J.-L.; Prieur, M.; Noel, B.; Sinet, P.-M. Molecular Mapping of Twenty-Four Features of Down Syndrome on Chromosome 21. Eur. J. Hum. Genet. 1993, 1, 114–124. [Google Scholar] [CrossRef]
- Korenberg, J.R.; Chen, X.N.; Schipper, R.; Sun, Z.; Gonsky, R.; Gerwehr, S.; Carpenter, N.; Daumer, C.; Dignan, P.; Disteche, C. Down syndrome phenotypes: The consequences of chromosomal imbalance. Proc. Natl. Acad. Sci. USA 1994, 91, 4997–5001. [Google Scholar] [CrossRef]
- Korbel, J.O.; Tirosh-Wagner, T.; Urban, A.E.; Chen, X.-N.; Kasowski, M.; Dai, L.; Grubert, F.; Erdman, C.; Gao, M.C.; Lange, K.; et al. The genetic architecture of Down syndrome phenotypes revealed by high-resolution analysis of human segmental trisomies. Proc. Natl. Acad. Sci. USA 2009, 106, 12031–12036. [Google Scholar] [CrossRef]
- Gupta, M.; Dhanasekaran, A.R.; Gardiner, K.J. Mouse models of Down syndrome: Gene content and consequences. Mamm. Genome 2016, 27, 538–555. [Google Scholar] [CrossRef] [PubMed]
- Moyer, A.J.; Gardiner, K.; Reeves, R.H. All Creatures Great and Small: New Approaches for Understanding Down Syndrome Genetics. Trends Genet. 2020, 37, 444–459. [Google Scholar] [CrossRef]
- Davisson, M.T.; Schmidt, C.; Akeson, E.C. Segmental trisomy of murine chromosome 16: A new model system for studying Down syndrome. Prog. Clin. Boil. Res. 1990, 360, 263–280. [Google Scholar]
- Choong, X.Y.; Tosh, J.L.; Pulford, L.J.; Fisher, E.M. Dissecting Alzheimer disease in Down syndrome using mouse models. Front. Behav. Neurosci. 2015, 9, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Rueda, N.; Flórez, J.; Martínez-Cué, C. Mouse models of Down syndrome as a tool to unravel the causes of mental disabilities. Neural Plast. 2012, 584071. [Google Scholar] [CrossRef]
- Herault, Y.; Delabar, J.M.; Fisher, E.M.C.; Tybulewicz, V.L.J.; Yu, E.; Brault, V. Rodent models in Down syndrome research: Impact and future opportunities. Dis. Model. Mech. 2017, 10, 1165–1186. [Google Scholar] [CrossRef]
- Moreno, M.D.M.M.; Brault, V.; Birling, M.-C.; Pavlovic, G.; Herault, Y. Modeling Down syndrome in animals from the early stage to the 4.0 models and next. Prog Brain Res. 2019, 251, 91–143. [Google Scholar] [CrossRef]
- Kazuki, Y.; Gao, F.J.; Li, Y.; Moyer, A.J.; Devenney, B.; Hiramatsu, K.; Miyagawa-Tomita, S.; Abe, S.; Kazuki, K.; Kajitani, N.; et al. A non-mosaic transchromosomic mouse model of down syn-drome carrying the long arm of human chromosome 21. Elife 2020, 9, e56223. [Google Scholar] [CrossRef]
- Reeves, R.H.; Irving, N.G.; Moran, T.H.; Wohn, A.; Kitt, C.; Sisodia, S.S.; Schmidt, C.; Bronson, R.T.; Davisson, M.T. A mouse model for Down syndrome exhibits learning and behaviour deficits. Nat. Genet. 1995, 11, 177–184. [Google Scholar] [CrossRef]
- Davisson, M.T.; Schmidt, C.; Reeves, R.H.; Irving, N.G.; Akeson, E.C.; Harris, B.S.; Bronson, R.T. Segmental trisomy as a mouse model for Down syndrome. Prog. Clin. Boil. Res. 1993, 384, 117–133. [Google Scholar]
- Duchon, A.; Raveau, M.; Chevalier, C.; Nalesso, V.; Sharp, A.J.; Herault, Y. Identification of the translocation breakpoints in the Ts65Dn and Ts1Cje mouse lines: Relevance for modeling down syndrome. Mamm. Genome 2011, 22, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cué, C.; Martínez, P.; Rueda, N.; Vidal, R.; García, S.; Vidal, V.; Corrales, A.; Montero, J.A.; Pazos, Á.; Flórez, J.; et al. Reducing GABAA α5 receptor-mediated inhibition rescues functional and neuromorphological deficits in a mouse model of down syndrome. J. Neurosci. 2013, 33, 3953–3966. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.S.; Hawkins, C.; Franca, A.; Lawler, A.; Devenney, B.; Das, I.; Reeves, R.H. Increased male reproductive success in Ts65Dn “Down syndrome” mice. Mamm. Genome 2010, 21, 543–549. [Google Scholar] [CrossRef]
- Yu, T.; Liu, C.; Belichenko, P.; Clapcote, S.J.; Li, S.; Pao, A.; Kleschevnikov, A.; Bechard, A.R.; Asrar, S.; Chen, R.; et al. Effects of individual segmental trisomies of human chromosome 21 syntenic regions on hippocampal long-term potentiation and cognitive behaviors in mice. Brain Res. 2010, 1366, 162–171. [Google Scholar] [CrossRef]
- Yu, T.; Li, Z.; Jia, Z.; Clapcote, S.J.; Liu, C.; Li, S.; Asrar, S.; Pao, A.; Chen, R.; Fan, N.; et al. A mouse model of Down syndrome trisomic for all human chromosome 21 syntenic regions. Hum. Mol. Genet. 2010, 19, 2780–2791. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.M.; Guedj, F.; Pennings, J.L.A.; Olmos-Serrano, J.L.; Siegel, A.; Haydar, T.F.; Bianchi, D.W. Lifespan analysis of brain development, gene expression and behavioral phenotypes in the Ts1Cje, Ts65Dn and Dp(16)1/Yey mouse models of Down syndrome. Dis. Model. Mech. 2018, 11. [Google Scholar] [CrossRef]
- Chang, P.; Bush, D.; Schorge, S.; Good, M.; Canonica, T.; Shing, N.; Noy, S.; Wiseman, F.K.; Burgess, N.; Tybulewicz, V.L.; et al. Altered Hippocampal-Prefrontal Neural Dynamics in Mouse Models of Down Syndrome. Cell Rep. 2020, 30, 1152–1163. [Google Scholar] [CrossRef]
- Belichenko, P.V.; Kleschevnikov, A.M.; Becker, A.; Wagner, G.E.; Lysenko, L.V.; Yu, Y.E.; Mobley, W.C. Down Syndrome Cognitive Phenotypes Modeled in Mice Trisomic for All HSA 21 Homologues. PLoS ONE 2015, 10, e0134861. [Google Scholar] [CrossRef]
- Goodliffe, J.W.; Olmos-Serrano, J.L.; Aziz, N.M.; Pennings, J.L.; Guedj, F.; Bianchi, D.W.; Haydar, T.F. Absence of Prenatal Forebrain Defects in the Dp(16)1Yey/+ Mouse Model of Down Syndrome. J. Neurosci. 2016, 36, 2926–2944. [Google Scholar] [CrossRef]
- Radulovic, J.; Kammermeier, J.; Spiess, J. Generalization of fear responses in C57BL/6N mice subjected to one-trial fore-ground contextual fear conditioning. Behav. Brain Res. 1998, 95, 179–189. [Google Scholar] [CrossRef]
- Ahmed, M.M.; Dhanasekaran, A.R.; Block, A.; Tong, S.; Costa, A.C.; Gardiner, K.J. Protein Profiles Associated with Context Fear Conditioning and Their Modulation by Memantine. Mol. Cell. Proteom. 2014, 13, 919–937. [Google Scholar] [CrossRef]
- Paylor, R.; Tracy, R.; Wehner, J.; Rudy, J.W. DBA/2 and C57BL/6 mice differ in contextual fear but not auditory fear condi-tioning. Behav. Neurosci. 1994, 108, 810–817. [Google Scholar] [CrossRef]
- Davisson, M.T.; Bechtel, L.J.; Akeson, E.C.; Fortna, A.; Slavov, D.; Gardiner, K. Evolutionary breakpoints on human chro-mosome 21. Genomics 2001, 78, 99–106. [Google Scholar] [CrossRef]
- Atkins, C.M.; Selcher, J.C.; Petraitis, J.J.; Trzaskos, J.M.; Sweatt, J.D. The MAPK cascade is required for mammalian associa-tive learning. Nat. Neurosci. 1998, 1, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.M.; Dhanasekaran, A.R.; Block, A.; Tong, S.; Costa, A.C.; Stasko, M.; Gardiner, K.J. Protein Dynamics Associated with Failed and Rescued Learning in the Ts65Dn Mouse Model of Down Syndrome. PLoS ONE 2015, 10, e0119491. [Google Scholar] [CrossRef] [PubMed]
- Franconi, F.; Campesi, I. Pharmacogenomics, pharmacokinetics and pharmacodynamics: Interaction with biological differ-ences between men and women. Br. J. Pharmacol. 2014, 171, 580–594. [Google Scholar] [CrossRef] [PubMed]
- Koss, W.A.; Frick, K.M. Sex differences in hippocampal function. J. Neurosci. Res. 2017, 95, 539–562. [Google Scholar] [CrossRef] [PubMed]
- Moyer, A.M.; Matey, E.T.; Miller, V.M. Individualized medicine: Sex, hormones, genetics, and adverse drug reactions. Pharmacol. Res. Perspect. 2019, 7, e00541. [Google Scholar] [CrossRef]
- Hornung, J.; Lewis, C.A.; Derntl, B. Sex hormones and human brain function. Handb. Clin. Neurol. 2020, 175, 195–207. [Google Scholar] [CrossRef]
- Gall, C.M.; Le, A.A.; Lynch, G. Sex differences in synaptic plasticity underlying learning. J. Neurosci. Res. 2021. [Google Scholar] [CrossRef]
- Dalla, C.; Shors, T.J. Sex differences in learning processes of classical and operant conditioning. Physiol. Behav. 2009, 97, 229–238. [Google Scholar] [CrossRef]
- McEwen, B.S.; Milner, T.A. Understanding the broad influence of sex hormones and sex differences in the brain. J. Neurosci. Res. 2017, 95, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Brandt, N.; Rune, G.M. Sex-dependency of oestrogen-induced structural synaptic plasticity: Inhibition of aromatase versus application of estradiol in rodents. Eur. J. Neurosci. 2020, 52, 2548–2559. [Google Scholar] [CrossRef] [PubMed]
- Gozzo, S.; Renzi, P.; D’Udine, B. Morphological Differences in Cerebral Cortex and Corpus Callosum Are Genetically Determined in Two Different Strains of Mice. Int. J. Neurosci. 1979, 9, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Crusio, W.E.; Schwegler, H.; van Abeelen, J.H. Behavioural and neuroanatomical divergence between two sublines of C57BL/6J inbred mice. Behav. Brain Res. 1991, 42, 93–97. [Google Scholar] [CrossRef]
- Kittler, P.; Krinsky-McHale, S.J.; Devenny, D.A. Sex differences in performance over 7 years on the Wechsler Intelligence Scale for Children—Revised among adults with intellectual disability. J. Intellect. Disabil. Res. 2004, 48, 114–122. [Google Scholar] [CrossRef]
- de Sola, S.; The TESDAD Study Group; de la Torre, R.; Sã¡nchez-Benavides, G.; Benejam, B.; Cuenca-Royo, A.; Del Hoyo, L.; Rodrãguez, J.; Catuara-Solarz, S.; Sanchez-Gutierrez, J.; et al. A new cognitive evaluation battery for Down syndrome and its relevance for clinical trials. Front. Psychol. 2015, 6, 708. [Google Scholar] [CrossRef] [PubMed]
- Aoki, S.; Yamauchi, Y.; Hashimoto, K. Developmental trend of children with Down’s syndrome—How do sex and neonatal conditions influence their developmental patterns? Brain Dev. 2018, 40, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Block, A.; Ahmed, M.M.; Dhanasekaran, A.R.; Tong, S.; Gardiner, K.J. Sex differences in protein expression in the mouse brain and their perturbations in a model of Down syndrome. Biol. Sex. Differ. 2015, 6, 24. [Google Scholar] [CrossRef]
- Meyer, R.; Wolf, S.S.; Obendorf, M. PRMT2, a member of the protein arginine methyltransferase family, is a coactivator of the androgen receptor. J. Steroid Biochem. Mol. Biol. 2007, 107, 1–14. [Google Scholar] [CrossRef]
- Qi, C.; Chang, J.; Zhu, Y.; Yeldandi, A.V.; Rao, S.M.; Zhu, Y.-J. Identification of Protein Arginine Methyltransferase 2 as a Coactivator for Estrogen Receptor α. J. Biol. Chem. 2002, 277, 28624–28630. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Cai, C.; Omwancha, J.; Chen, S.-Y.; Baslan, T.; Shemshedini, L. SUMO-3 enhances androgen receptor transcrip-tional activity through a sumoylation-independent mechanism in prostate cancer cells. J. Biol. Chem. 2006, 281, 4002–4012. [Google Scholar] [CrossRef] [PubMed]
- Rytinki, M.; Kaikkonen, S.; Sutinen, P.; Paakinaho, V.; Rahkama, V.; Palvimo, J.J. Dynamic SUMOylation Is Linked to the Activity Cycles of Androgen Receptor in the Cell Nucleus. Mol. Cell. Biol. 2012, 32, 4195–4205. [Google Scholar] [CrossRef]
- Yang, N.; Liu, S.; Qin, T.; Liu, X.; Watanabe, N.; Mayo, K.H.; Li, J.; Li, X. SUMO3 modification by PIAS1 modulates androgen receptor cellular distribution and stability. Cell Commun. Signal. 2019, 17, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hood, J.L.; Emeson, R.B. Editing of Neurotransmitter Receptor and Ion Channel RNAs in the Nervous System. In Current Topics in Microbiology and Immunology; Samuel, C.E., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 353, pp. 61–90. [Google Scholar] [CrossRef]
- Horsch, M.; Seeburg, P.H.; Adler, T.; Aguilar-Pimentel, J.A.; Becker, L.; Calzada, J.; Garrett, L.; Götz, A.; Hans, W.; Higuchi, M.; et al. Requirement of the RNA-editing Enzyme ADAR2 for Normal Physiology in Mice. J. Biol. Chem. 2011, 286, 18614–18622. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Verma, S.; Nakayama, S.; Quillinan, N.; Grafe, M.R.; Hurn, P.D.; Herson, P.S. Sex Differences in Neuroprotection Provided by Inhibition of TRPM2 Channels following Experimental Stroke. J. Cereb. Blood Flow Metab. 2011, 31, 2160–2168. [Google Scholar] [CrossRef]
- Shimizu, T.; Macey, T.A.; Quillinan, N.; Klawitter, J.; Perraud, A.-L.L.; Traystman, R.J.; Herson, P.S. Androgen and PARP-1 Regulation of TRPM2 Channels after Ischemic Injury. J. Cereb. Blood Flow Metab. 2013, 33, 1549–1555. [Google Scholar] [CrossRef]
- Dhanasekaran, A.R.; Block, A.; Ahmed, M.; Gardiner, K.J. Abnormal Protein Profiles in Hippocampus of Mouse Models of Down Syndrome: Similarities with Alzheimer’s Disease. J. Alzheimer’s Dis. Park. 2014, 4, 138–144. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Female Control | Female Trisomic | Male Control | Male Trisomic | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SC | CS | NS | SC | CS | NS | SC | CS | NS | SC | CS | NS | |
| Experiment 1 | ||||||||||||
| Dp16 | 12 | 11 | 9 | 10 | 13 | 16 | 10 | 10 | ||||
| Dp17 | 12 | 13 | 13 | 16 | 11 | 12 | 10 | 10 | ||||
| Dp10 | 11 | 13 | 12 | 16 | 12 | 20 | 10 | 19 | ||||
| Experiment 2 | ||||||||||||
| DBA | 8 | 9 | 11 | 11 | ||||||||
| DBA X Dp17 | 7 | 7 | 10 | 10 | 6 | 9 | 7 | 9 | ||||
| DBA X Dp10 | 15 | 16 | 9 | 9 | 9 | 9 | 9 | 8 | ||||
| Experiment 3 | ||||||||||||
| Dp17 | 7 | 8 | 7 | 6 | 9 | 6 | 5 | 10 | 11 | 5 | ||
| Dp10 | 10 | 11 | 5 | 10 | 10 | 5 | 8 | 10 | 7 | 9 | 13 | 7 |
| SC | SC-t | CS | CS-t | |||
|---|---|---|---|---|---|---|
| Dp16 | Female | C | 13 | 18 | 13 | 66 |
| Tr | 8 | 10 | 10 | 23 | ||
| Male | C | 14 | 30 | 7 | 98 | |
| Tr | 7 | 12 | 7 | 32 | ||
| Dp17 | Female | C | 29 | 39 | 15 | 86 |
| Tr | 35 | 35 | 11 | 83 | ||
| Male | C | 18 | 31 | 11 | 96 | |
| Tr | 19 | 50 | 31 | 93 | ||
| Dp10 | Female | C | 19 | 22 | 13 | 70 |
| Tr | 17 | 19 | 11 | 53 | ||
| Male | C | 16 | 26 | 13 | 81 | |
| Tr | 17 | 24 | 14 | 52 |
| SC | SC-t | CS | CS-t | |||
|---|---|---|---|---|---|---|
| DBA/2J | Female | C | 91 | 66 | 60 | 127 |
| Male | C | 64 | 55 | 57 | 84 | |
| DBA X Dp17 | Female | C | 15 | 29 | 14 | 97 |
| Tr | 21 | 22 | 11 | 104 | ||
| Male | C | 16 | 23 | 11 | 73 | |
| Tr | 21 | 28 | 14 | 83 | ||
| DBA X Dp10 | Female | C | 37 | 52 | 13 | 86 |
| Tr | 23 | 37 | 13 | 60 | ||
| Male | C | 35 | 38 | 15 | 101 | |
| Tr | 26 | 29 | 17 | 73 |
| SC | SC-t | CS | CS-t | NS | NS-t | |||
|---|---|---|---|---|---|---|---|---|
| Dp17 | Female | C | 25 | 55 | 10 | 56 | NA | NA |
| Tr | 15 | 26 | 11 | 46 | NA | NA | ||
| Male | C | 28 | 61 | 7 | 88 | 11 | 39 | |
| Tr | 53 | 83 | 11 | 107 | 9 | 12 | ||
| Dp10 | Female | C | 24 | 38 | 13 | 31 | 9 | 21 |
| Tr | 20 | 38 | 8 | 24 | 6 | 17 | ||
| Male | C | 44 | 62 | 11 | 73 | 13 | 25 | |
| Tr | 19 | 60 | 9 | 88 | 8 | 23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, M.M.; Block, A.; Busquet, N.; Gardiner, K.J. Context Fear Conditioning in Down Syndrome Mouse Models: Effects of Trisomic Gene Content, Age, Sex and Genetic Background. Genes 2021, 12, 1528. https://doi.org/10.3390/genes12101528
Ahmed MM, Block A, Busquet N, Gardiner KJ. Context Fear Conditioning in Down Syndrome Mouse Models: Effects of Trisomic Gene Content, Age, Sex and Genetic Background. Genes. 2021; 12(10):1528. https://doi.org/10.3390/genes12101528
Chicago/Turabian StyleAhmed, Md. Mahiuddin, Aaron Block, Nicolas Busquet, and Katheleen J. Gardiner. 2021. "Context Fear Conditioning in Down Syndrome Mouse Models: Effects of Trisomic Gene Content, Age, Sex and Genetic Background" Genes 12, no. 10: 1528. https://doi.org/10.3390/genes12101528
APA StyleAhmed, M. M., Block, A., Busquet, N., & Gardiner, K. J. (2021). Context Fear Conditioning in Down Syndrome Mouse Models: Effects of Trisomic Gene Content, Age, Sex and Genetic Background. Genes, 12(10), 1528. https://doi.org/10.3390/genes12101528