Confirmation of Paternity despite Three Genetic Incompatibilities at Chromosome 2

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. DNA Samples
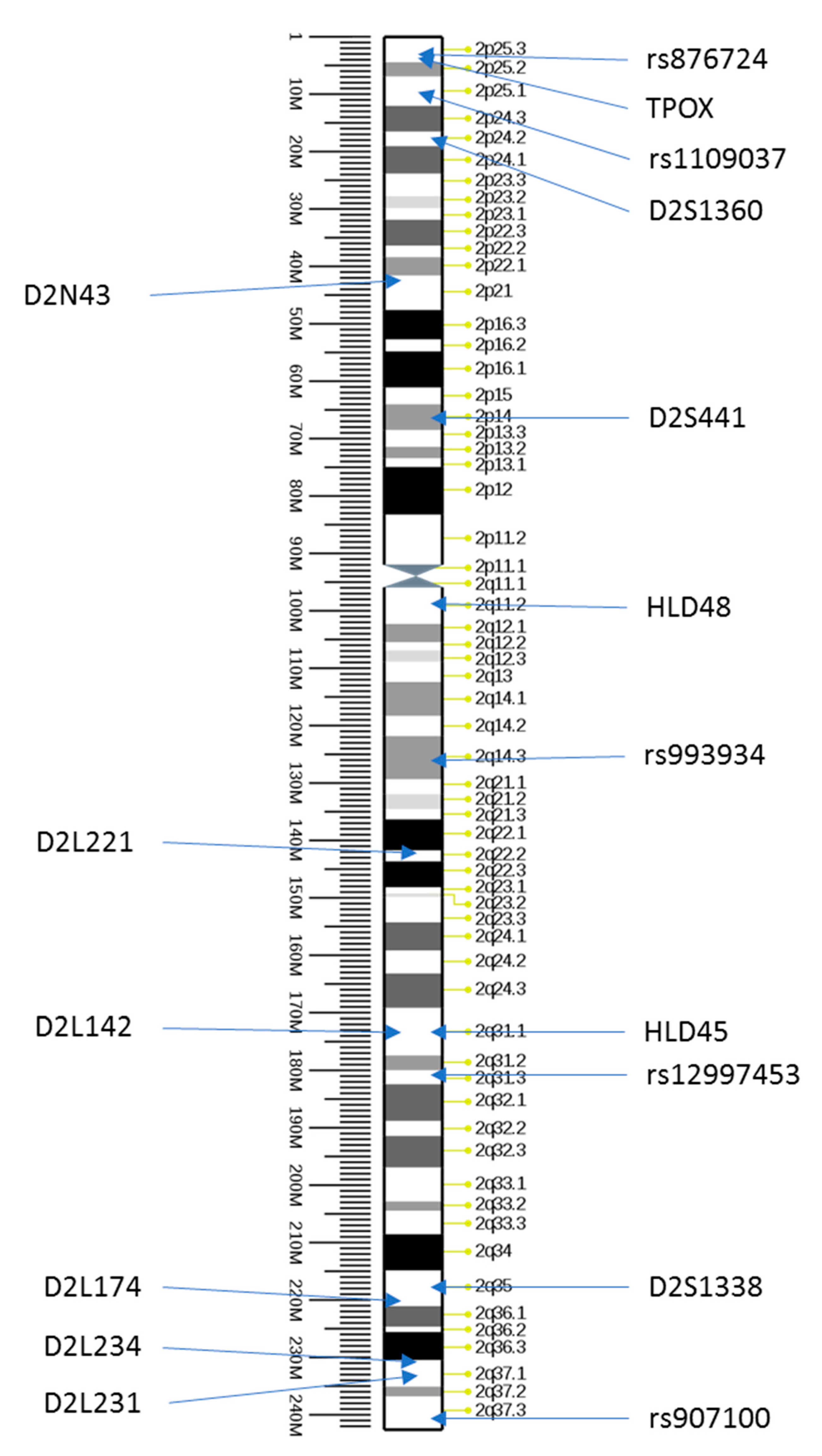
2.2. Selection and Analysis of New STR Markers on Chromosome 2
- D2N43—FP: 6FAM-5′TTAA TAAA TGCA CTCA CACT CTAG ATAG, RP: 5′GTTC CAGG AGCA TCTC CATC C3′;
- D2L221—FP: 6FAM-5′CATC CAGC AGGA TTTC TTTC T, RP: TGTG AGCT ATGA TTAT GCCA TTG3′;
- D2L142—FP: 6FAM-5′CATT GAAA TAAT TAAC CTCA CATT TTC 3′, RP: 5′GAAA CTAA ATGT CAGT TGTT TTC 3′;
- D2L174—FP: TAMRA-5′CGGA TACA CACC ACTG TTGG AC3′, RP: 5′AGCC AGAA AAGC ATAC CCGT3′;
- D2L114—FP: HEX-5′GATA AATT CCAC TCTT GGTC ATAT AC3′, RP: 5′GTCA GTGC TAGA GGCA TATT TACA3′;
- D2L234—FP: ROX-5′AAAG TGCA AGGT TTGA AGCC3′, RP: 5′AGCT GTGG TTGG CGAT CATT3′;
- D2L231—FP: 6-FAM-5′AGGC TGAT CATT TGAC TTTC TTTG T3′, RP; 5′CACA TGCT CTCA CTCA TAAG TGGA3′.
2.2.1. PCR Reaction
2.2.2. Electrophoresis
2.3. Quantification and PCR-CE Analysis with Commercially Available STR and In-Del Kits
2.4. Next Generation Sequencing
2.5. Statistical Analysis
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| Locus | PF | Daughter | Mother |
|---|---|---|---|
| Amelogenin | X,Y | X,X | X,X |
| D1S1656 | 16, 19.3 | 16, 17.3 | 16, 17.3 |
| TPOX | 8, 8 | 9, 11 | 9, 11 |
| D2S441 | 11, 15 | 11.3, 11.3 | 11.3, 14 |
| D2S1338 | 18, 18 | 21, 23 | 21, 23 |
| D3S1358 | 18, 18 | 17,18 | 17, 18 |
| D4S2408 | 8, 9 | 8, 9 | 8, 12 |
| FGA | 20, 21 | 20, 21 | 21, 26 |
| D5S818 | 12, 12 | 9, 12 | 9, 10 |
| CSF1PO | 10, 10 | 10, 10 | 10, 10 |
| D6S1043 | 11, 19 | 11, 19 | 11, 11 |
| D7S820 | 11, 12 | 9, 12 | 9, 10 |
| D8S1179 | 10, 14 | 10, 14 | 8, 10 |
| D9S1122 | 13, 13 | 12, 13 | 12, 12 |
| D10S1248 | 13, 17 | 13, 13 | 13, 14 |
| TH01 | 6, 7 | 6, 9.3 | 9, 9.3 |
| vWA | 16, 16 | 16, 18 | 17, 18 |
| D12S391 | 18, 19 | 19, 22 | 21, 22 |
| D13S317 | 8, 8 | 8, 8 | 8, 12 |
| PentaE | 11, 17 | 14, 17 | 12, 14 |
| D16S539 | 12, 13 | 11, 12 | 11, 11 |
| D17S1301 | 11, 13 | 12, 13 | 12, 13 |
| D18S51 | 15, 18 | 17, 18 | 13, 17 |
| D19S433 | 13, 14 | 13, 14 | 14, 15 |
| D20S482 | 11, 14 | 11, 13 | 13, 15 |
| D21S11 | 29, 31.2 | 28, 29 | 28, 32.2 |
| PentaD | 12, 13 | 12, 16 | 10, 16 |
| D22S1045 | 11, 16 | 16, 16 | 15, 16 |
| DYS391 | 10 | - | - |
| SE33 (actB) | 26.2, 27.2 | 18, 27.2 | 18, 27.2 |
| DXS10135 | 26 | 26 | 22, 26 |
| DXS8378 | 13 | 13 | 11, 13 |
| DXS7132 | 15 | 12, 15 | 12, 14 |
| DXS10074 | 19 | 8, 19 | 8, 17 |
| DXS10103 | 19 | 19, 20 | 20, 21 |
| HPRTB | 12 | 12 | 12 |
| DXS7423 | 15 | 15 | 14, 15 |
| Penta E | 11, 17 | 14, 17 | 12, 14 |
| D2S1360 | 22 | 22 | 22 |
| D3S1744 | 14, 18 | 18 | 16, 18 |
| D4S2366 | 13, 14 | 9,14 | 9, 11 |
| D5S2500 | 12 | 12, 16 | 16 |
| D6S474 | 13 | 13, 14 | 14, 15 |
| D7S1517 | 24, 25 | 24, 25 | 25 |
| D8S1132 | 20, 21 | 20, 22 | 22, 23 |
| D10S2325 | 7, 13 | 7, 13 | 8, 13 |
| D21S2055 | 26, 29, 33 | 19.1, 29 | 19.1, 33 |
| Locus | AF | Daughter | Mother |
|---|---|---|---|
| rs1490413 | G,A | G,G | G,G |
| rs560681 | A,G | A,A | A,A |
| rs1294331 | G,A | A,A | G,A |
| rs10495407 | G,A | A,A | G,A |
| rs891700 | G,G | A,G | A,G |
| rs1413212 | G,G | A,G | A,A |
| rs876724 | C,C | C,C | C,C |
| rs1109037 | A,A | A,A | G,A |
| rs993934 | T,T | T,T | T,C |
| rs12997453 | G,G | A,G | A,G |
| rs907100 | G,C | C,C | C,C |
| rs1357617 | A,A | T,A | T,T |
| rs4364205 | G,G | T,G | T,G |
| rs2399332 | A,A | A,C | C,C |
| rs1355366 | A,G | A,G | A,G |
| rs6444724 | T,C | C,C | T,C |
| rs2046361 | A,T | A,T | T,T |
| rs279844 | A,A | A,A | A,A |
| rs6811238 | G,G | T,G | T,G |
| rs1979255 | G,C | G,C | C,C |
| rs717302 | G,A | A,A | A,A |
| rs159606 | A,G | G,G | G,G |
| rs13182883 | G,A | G,A | G,G |
| rs251934 | T,C | C,C | T,C |
| rs338882 | C,T | C,T | T,T |
| rs13218440 | G,A | G,G | G,G |
| rs1336071 | G,G | G,G | G,G |
| rs214955 | G,A | A,A | A,A |
| rs727811 | A,A | C,A | C,C |
| rs6955448 | C,C | C,T | T,T |
| rs917118 | C,T | C,C | C,C |
| rs321198 | T,T | T,T | T,C |
| rs737681 | C,C | C,C | C,C |
| rs763869 | C,T | C,C | C,C |
| rs10092491 | T,T | T,C | C,C |
| rs2056277 | C,T | T,T | C,T |
| rs4606077 | T,C | C,C | T,C |
| rs1015250 | G,G | G,G | G,C |
| rs7041158 | C,T | C,C | C,T |
| rs1463729 | A,A | A,A | A,A |
| rs1360288 | C,C | C,C | C,T |
| rs10776839 | T,T | T,T | T,T |
| rs826472 | C,C | T,C | T,T |
| rs735155 | G,G | G,A | G,A |
| rs3780962 | T,C | T,C | T,C |
| rs740598 | A,A | A,A | A,A |
| rs964681 | T,C | T,T | T,T |
| rs1498553 | C,T | T,T | T,T |
| rs901398 | T,T | T,T | C,T |
| rs10488710 | C,C | C,C | G,C |
| rs2076848 | T,T | T,A | T,A |
| rs2107612 | A,A | A,A | A,A |
| rs2269355 | C,G | C,G | G,G |
| rs2920816 | C,C | T,C | T,C |
| rs2111980 | G,G | G,G | G,G |
| rs10773760 | A,A | A,A | A,G |
| rs1335873 | T,T | A,T | A,T |
| rs1886510 | C,T | C,T | T,T |
| rs1058083 | A,G | G,G | G,G |
| rs354439 | T,T | T,T | T,T |
| rs1454361 | A,A | A,A | A,A |
| rs722290 | C,C | C,G | C,G |
| rs873196 | C,T | T,T | C,T |
| rs4530059 | G,G | G,A | G,A |
| rs1821380 | G,C | G,C | C,C |
| rs8037429 | C,T | C,T | C,T |
| rs1528460 | C,T | C,T | T,T |
| rs729172 | C,C | C,C | C,C |
| rs2342747 | G,G | A,G | A,G |
| rs430046 | C,C | C,C | C,T |
| rs1382387 | G,T | T,T | T,T |
| rs9905977 | A,G | G,G | G,G |
| rs740910 | A,A | A,A | A,A |
| rs938283 | T,C | T,T | T,T |
| rs8078417 | C,C | C,C | C,C |
| rs1493232 | A,A | A,A | A,A |
| rs9951171 | A,A | A,A | G,A |
| rs1736442 | A,G | A,A | A,A |
| rs1024116 | G,A | A,A | G,A |
| rs719366 | T,T | C,T | C,C |
| rs576261 | A,A | A,C | A,C |
| rs1031825 | C,C | C,C | C,C |
| rs445251 | C,G | G,G | C,G |
| rs1005533 | G,A | G,A | G,A |
| rs1523537 | T,C | T,T | T,C |
| rs722098 | A,A | A,A | A,A |
| rs2830795 | A,A | A,A | A,G |
| rs2831700 | A,G | G,G | A,G |
| rs914165 | A,A | G,A | G,A |
| rs221956 | C,C | C,C | T,C |
| rs733164 | G,G | G,G | G,G |
| rs987640 | T,T | T,A | A,A |
| rs2040411 | A,A | A,A | A,A |
| rs1028528 | A,G | A,G | A,G |
| Locus | AF | Daughter | Mother |
|---|---|---|---|
| Amel. X | X | X | X |
| Amel. Y | Y | − | − |
| HLD77 | + | + | + |
| HLD45 | −/+ | + | + |
| HLD131 | −/+ | −/+ | − |
| HLD70 | + | + | + |
| HLD6 | −/+ | + | + |
| HLD111 | −/+ | −/+ | −/+ |
| HLD58 | −/+ | + | + |
| HLD56 | −/+ | −/+ | + |
| HLD118 | + | −/+ | −/+ |
| HLD92 | + | −/+ | −/+ |
| HLD93 | + | + | + |
| HLD99 | + | −/+ | −/+ |
| HLD88 | − | − | − |
| HLD101 | −/+ | −/+ | −/+ |
| HLD67 | + | −/+ | − |
| HLD83 | + | −/+ | − |
| HLD114 | + | + | + |
| HLD48 | − | − | − |
| HLD124 | −/+ | −/+ | + |
| HLD122 | −/+ | − | − |
| HLD125 | - | −/+ | + |
| HLD64 | - | − | − |
| HLD81 | - | −/+ | −/+ |
| HLD136 | + | + | −/+ |
| HLD133 | + | −/+ | −/+ |
| HLD97 | −/+ | + | + |
| HLD40 | + | + | + |
| HLD128 | −/+ | −/+ | −/+ |
| HLD39 | − | − | − |
| HLD84 | + | −/+ | − |
References
- Wojtas, M.; Piniewska, D.; Polańska, N.; Stawowiak, A.; Sanak, M. Mutations of microsatellite autosomal loci in paternity investigations of the Southern Poland population. Forensic Sci. Int. Genet. 2013, 7, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, K.; Ochiai, E.; Tamura, T.; Osawa, M. Kinship analysis based on SNP data from the HID-Ion AmpliSeq identity panel. J. Forensic. Investig. 2015, 3, 6. [Google Scholar]
- Krawczak, M.; Budowle, B.; Weber-Lehmann, J.; Rolf, B. Distinguishing genetically between the germlines of male monozygotic twins. PLoS Genet. 2018, 14, e1007756. [Google Scholar] [CrossRef]
- Brinkmann, B.; Klintschar, M.; Neuhuber, F.; Hühne, J.; Rolf, B. Mutation rate in human microsatellites: Influence of the structure and length of the tandem repeat. Am. J. Hum. Genet. 1998, 62, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.M. Forensic DNA Typing, 2nd ed.; Elsevier Academic Press: Burlington, VT, Canada, 2005; p. 140. [Google Scholar]
- Ayadi, I.; Mahfoudh-Lahiani, N.; Makni, H.; Ammar-Keskes, L.; Rebaï, A. Combining autosomal and y-chromosomal short tandem repeat data in paternity testing with male child: Methods and application. J. Forensic Sci. 2007, 52, 1068–1072. [Google Scholar] [CrossRef]
- Liehr, T. Cytogenetic contribution to uniparental disomy (UPD). Mol. Cytogenet. 2010, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Engel, E. A new genetic concept: Uniparental disomy and its potential effect, isodisomy. Am. J. Med. Genet. 1980, 6, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Créau-Goldberg, N.; Gegonne, A.; Delabar, J.; Cochet, C.; Cabanis, M.O.; Stehelin, D.; Turleau, C.; de Grouchy, J. Maternal origin of a de novo balanced t (21q21q) identified by ets-2 polymorphism. Hum. Genet. 1987, 76, 396–398. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Yue, S.; Wang, L.; Zhang, T.; Guo, C.; Hu, W.; Kagan, K.-O.; Wu, Q. Uniparental disomy and prenatal phenotype: Two case reports and review. Medicine 2017, 96. [Google Scholar] [CrossRef]
- Novroski, N.M.; Woerner, A.E.; Budowle, B. Potential highly polymorphic short tandem repeat markers for enhanced forensic identity testing. Forensic Sci. Int. Genet. 2018, 37, 162–171. [Google Scholar] [CrossRef]
- Ossowski, A.; Diepenbroek, M.; Szargut, M.; Zielińska, G.; Jędrzejczyk, M.; Berent, J.; Jacewicz, R. Population analysis and forensic evaluation of 21 autosomal loci included in GlobalFiler™ PCR Kit in Poland. Forensic Sci. Int. Genetics 2017, 29, e38–e39. [Google Scholar] [CrossRef] [PubMed]
- Wróbel, M.; Parys-Proszek, A.; Marcińska, M.; Ba, G.; Sekuła, A.; Kowalczyk, M.; Januła, M.; Doniec, A.; Kupiec, T. Analysis of the frequency of occurrence in the polish population of alleles of 21 genetic markers in the Globalfiler kit. Probl. Forensic Sci. 2019, 117, 49–61. [Google Scholar]
- Gjertson, D.W.; Brenner, C.H.; Baur, M.P.; Carracedo, A.; Guidet, F.; Luque, J.A.; Lessig, R.; Mayr, W.R.; Pascali, V.L.; Prinz, M.; et al. ISFG: Recommendations on biostatistics in paternity testing. Forensic Sci. Int. Genetics 2007, 1, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Robinson, W.P. Mechanisms leading to uniparental disomy and their clinical consequences. Bioessays 2000, 22, 452–459. [Google Scholar] [CrossRef]
- Stratakis, C.A.; Taymans, S.E.; Schteingart, D.; Haddad, B.R. Segmental uniparental isodisomy (UPD) for 2p16 without clinical symptoms: Implications for UPD and other genetic studies of chromosome 2. J. Med. Genet. 2001, 38, 106–109. [Google Scholar] [CrossRef]
- Ting, J.C.; Roberson, E.D.; Miller, N.D.; Lysholm-Bernacchi, A.; Stephan, D.A.; Capone, G.T.; Ruczinski, I.; Thomas, G.H.; Pevsner, J. Visualization of uniparental inheritance, Mendelian inconsistencies, deletions, and parent of origin effects in single nucleotide polymorphism trio data with SNPtrio. Hum. Mutat. 2007, 28, 1225–1235. [Google Scholar] [CrossRef]
- Zhang, X.; Ding, Z.; He, R.; Qi, J.; Zhang, Z.; Cui, B. Complete paternal uniparental disomy of chromosome 2 in an Asian female identified by short tandem repeats and whole genome sequencing. Cytogenet. Genome Res. 2019, 157, 197–202. [Google Scholar] [CrossRef]
- Guzmán-Alberto, J.C.; Martínez-Cortes, G.; Rangel-Villalobos, H. Inference of maternal uniparental disomy of the entire chromosome 2 from a paternity test. Int. J. Leg. Med. 2019, 133, 71–75. [Google Scholar] [CrossRef]
- Su, H.; Sun, T.; Chen, M.; Liu, J.; Wang, X.; Chen, Y.; Ren, W.; Zhang, G.; Yan, J.; Yun, K. Multiple methods used for type detection of uniparental disomy in paternity testing. Int. J. Leg. Med. 2019, 1–9. [Google Scholar] [CrossRef]
- Bein, G.; Driller, B.; Schürmann, M.; Schneider, P.M.; Kirchner, H. Pseudo-exclusion from paternity due to maternal uniparental disomy 16. Int. J. Leg. Med. 1998, 111, 328–330. [Google Scholar] [CrossRef]
- Mansuet-Lupo, A.; Henke, J.; Henke, L.; Blank, C.; Ernsting, A.; Kozlowski, P.; Rouger, P.; Van Huffel, V. A paternity case with three genetic incompatibilities between father and child due to maternal uniparental disomy 21 and a mutation at the Y chromosome. Forensic Sci. Int. Genet. 2009, 3, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Heide, E.; Heide, K.G.; Rodewald, A. Maternal uniparental disomy (UPD) for chromosome 2 discovered by exclusion of paternity. Am. J. Med. Gen. 2000, 92, 260–263. [Google Scholar] [CrossRef]

| Name | Repeated Motif | Heterozygosity | Location | AF | Child | Mother |
|---|---|---|---|---|---|---|
| D2N43 * | (AGAT) | n/a | 42,072,419—42,072,494 | 458; 470 | 498 | 498 |
| D2L221 | (AAAG) | 0.864 | 142,567,661—142,567,756 | 259; 279 | 259 | 239; 259 |
| D2L142 | (AAAG) | 0.887 | 174,327,360—174,327,425 | 410; 430 | 424; 438 | 424; 438 |
| D2L174 | (AAAG) | 0.911 | 221,218,006—221,218,073 | 175; 189 | 193 | 193 |
| D2L234 | (AAAG) | 0.878 | 231,153,171—231,153,239 | 244; 253 | 244; 248 | 244; 248 |
| D2L231 | (AAAG) | 0.883 | 234,570,270—234,570,328 | 141; 162 | 159 | 159; 154 |
| Kit | Maker Type | Name | Location | AF | Child | Mother |
|---|---|---|---|---|---|---|
| ForenSeq | SNP | rs876724 | 2p25.3 | C,C | C,C | C,C |
| ForenSeq | SNP | rs1109037 | 2p25.1 | A,A | A,A | G,A |
| ForenSeq | SNP | rs993934 | 2q14.3 | T,T | T,T | T,C |
| ForenSeq | SNP | rs12997453 | 2q31.3 | G,G | A,G | A,G |
| ForenSeq | SNP | rs907100 | 2q37.3 | G,C | C,C | C,C |
| DIPplex | indel | HLD45 | 2q31.1 | −,+ | +,+ | +,+ |
| DIPplex | indel | HLD48 | 2q11.2 | −,− | −,− | −,− |
| GlobalFiler | STR | TPOX | 2p25.3 | 8,8 | 9,11 | 9,11 |
| HDplex | STR | D2S1360 | 2p24.2 | 22,22 | 22,22 | 22,22 |
| GlobalFiler | STR | D2S441 | 2p14 | 11,15 | 11.3,11.3 | 11.3,14 |
| GlobalFiler | STR | D2S1338 | 2q35 | 18,18 | 21,23 | 21,23 |
| POI | Allele | Nucleotide Sequence of Allele |
|---|---|---|
| Daughter | 11.3 | (TCTA)4 TC A (TCTA)7 A (TCTA)7 |
| 11.3 | (TCTA)4 TCA (TCTA)7 | |
| AF | 11 | (TCTA)11 |
| 15 | (TCTA)15 | |
| Mother | 11.3 | (TCTA)4 TCA (TCTA)7 |
| 14 | (TCTA)14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doniec, A.; Łuczak, W.; Wróbel, M.; Januła, M.; Ossowski, A.; Grzmil, P.; Kupiec, T. Confirmation of Paternity despite Three Genetic Incompatibilities at Chromosome 2. Genes 2021, 12, 62. https://doi.org/10.3390/genes12010062
Doniec A, Łuczak W, Wróbel M, Januła M, Ossowski A, Grzmil P, Kupiec T. Confirmation of Paternity despite Three Genetic Incompatibilities at Chromosome 2. Genes. 2021; 12(1):62. https://doi.org/10.3390/genes12010062
Chicago/Turabian StyleDoniec, Andrzej, Wojciech Łuczak, Maria Wróbel, Miłosz Januła, Andrzej Ossowski, Paweł Grzmil, and Tomasz Kupiec. 2021. "Confirmation of Paternity despite Three Genetic Incompatibilities at Chromosome 2" Genes 12, no. 1: 62. https://doi.org/10.3390/genes12010062
APA StyleDoniec, A., Łuczak, W., Wróbel, M., Januła, M., Ossowski, A., Grzmil, P., & Kupiec, T. (2021). Confirmation of Paternity despite Three Genetic Incompatibilities at Chromosome 2. Genes, 12(1), 62. https://doi.org/10.3390/genes12010062