Genetic Hypothesis and Pharmacogenetics Side of Renin-Angiotensin-System in COVID-19



Abstract
1. Introduction
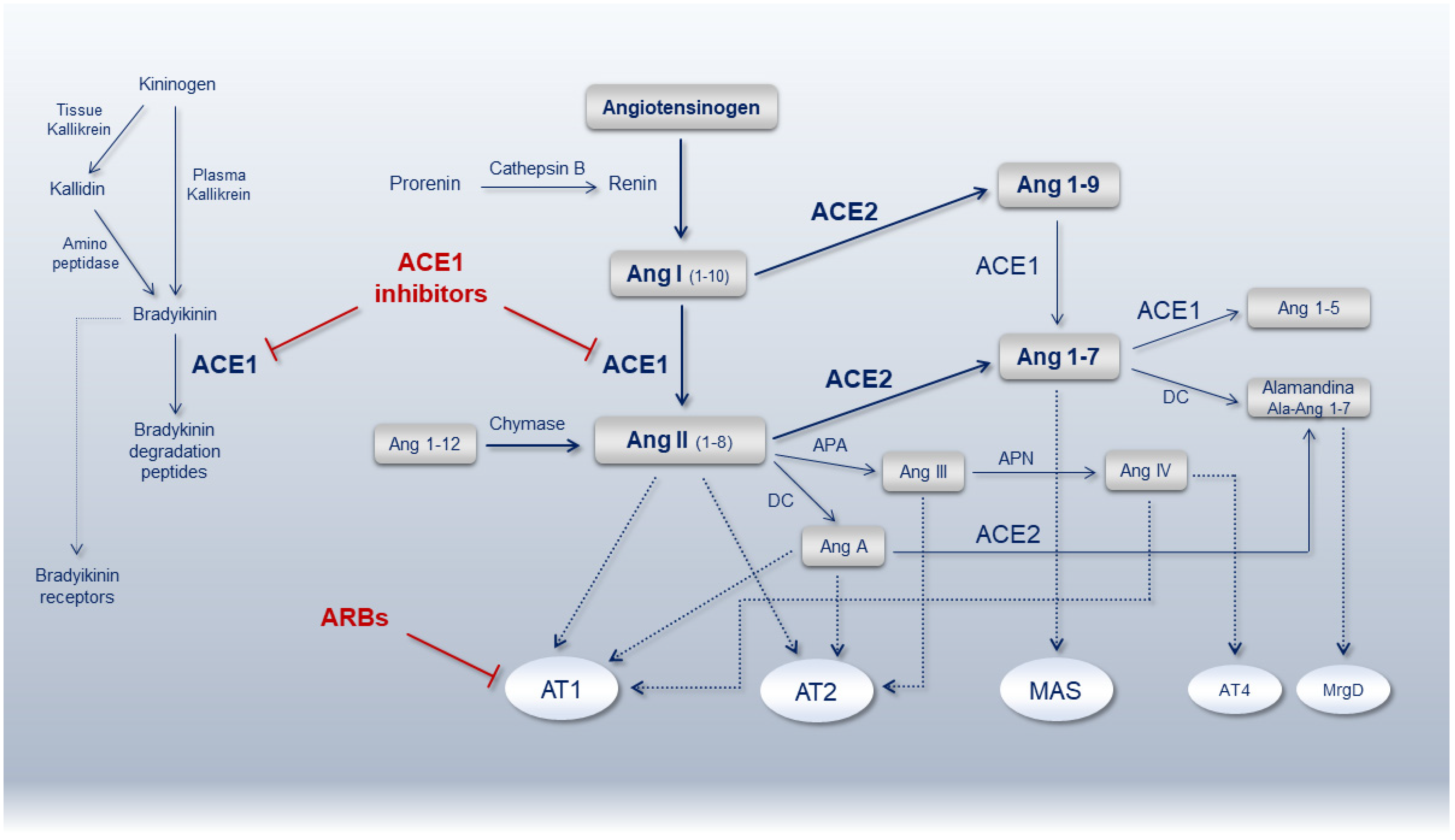
2. ACE1/ACE2 Balance
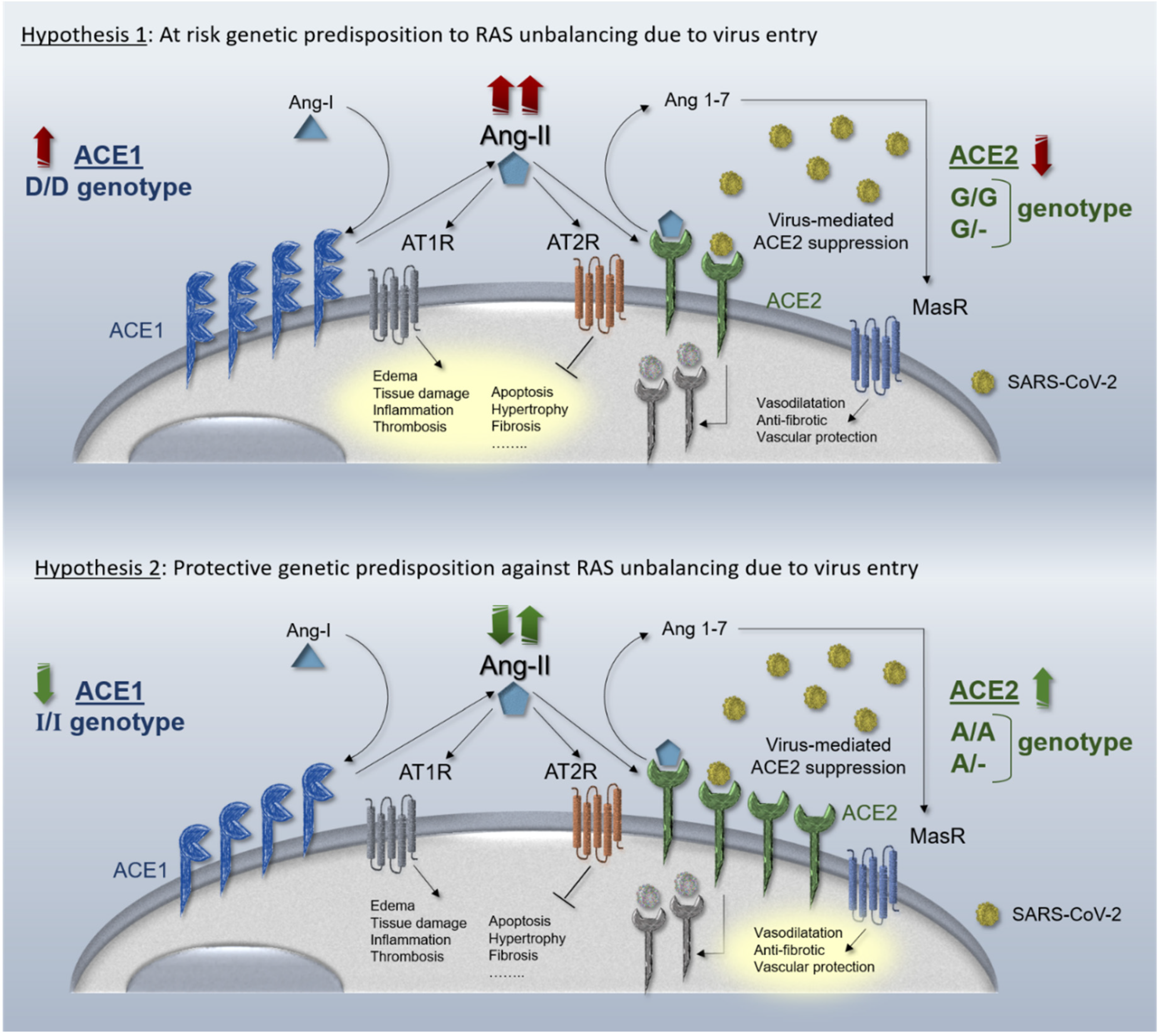
3. ACE1 and ACE2 Genes
4. ACEi and ARBs
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Gemmati, D.; Bramanti, B.; Serino, M.L.; Secchiero, P.; Zauli, G.; Tisato, V. COVID-19 and Individual Genetic Susceptibility/Receptivity: Role of ACE1/ACE2 Genes, Immunity, Inflammation and Coagulation. Might the Double X-chromosome in Females Be Protective against SARS-CoV-2 Compared to the Single X-Chromosome in Males? Int. J. Mol. Sci. 2020, 21, 3474. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, V.; Veeraveedu, P.T.; Gurusamy, N.; Lakshmanan, A.P.; Yamaguchi, K.; Ma, M.; Suzuki, K.; Nagata, M.; Takagi, R.; Kodama, M.; et al. Olmesartan attenuates the development of heart failure after experimental autoimmune myocarditis in rats through the modulation of ANG 1-7 mas receptor. Mol. Cell Endocrinol. 2012, 351, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M.; Jessup, J.; Chappell, M.C.; Averill, D.B.; Brosnihan, K.B.; Tallant, E.A.; Diz, D.I.; Gallagher, P.E. Effect of angiotensin-converting enzyme inhibition and angiotensin II receptor blockers on cardiac angiotensin-converting enzyme 2. Circulation 2005, 111, 2605–2610. [Google Scholar] [CrossRef]
- Sukumaran, V.; Veeraveedu, P.T.; Lakshmanan, A.P.; Gurusamy, N.; Yamaguchi, K.; Ma, M.; Suzuki, K.; Kodama, M.; Watanabe, K. Olmesartan medoxomil treatment potently improves cardiac myosin-induced dilated cardiomyopathy via the modulation of ACE-2 and ANG 1-7 mas receptor. Free Radic Res. 2012, 46, 850–860. [Google Scholar] [CrossRef]
- Soler, M.J.; Barrios, C.; Oliva, R.; Batlle, D. Pharmacologic modulation of ACE2 expression. Curr. Hypertens. Rep. 2008, 10, 410–414. [Google Scholar] [CrossRef]
- Campbell, D.J.; Zeitz, C.J.; Esler, M.D.; Horowitz, J.D. Evidence against a major role for angiotensin converting enzyme-related carboxypeptidase (ACE2) in angiotensin peptide metabolism in the human coronary circulation. J. Hypertens. 2004, 22, 1971–1976. [Google Scholar] [CrossRef]
- Furuhashi, M.; Moniwa, N.; Mita, T.; Fuseya, T.; Ishimura, S.; Ohno, K.; Shibata, S.; Tanaka, M.; Watanabe, Y.; Akasaka, H.; et al. Urinary angiotensin-converting enzyme 2 in hypertensive patients may be increased by olmesartan, an angiotensin II receptor blocker. Am. J. Hypertens. 2015, 28, 15–21. [Google Scholar] [CrossRef]
- Epelman, S.; Shrestha, K.; Troughton, R.W.; Francis, G.S.; Sen, S.; Klein, A.L.; Tang, W.H. Soluble angiotensin-converting enzyme 2 in human heart failure: Relation with myocardial function and clinical outcomes. J. Card. Fail. 2009, 15, 565–571. [Google Scholar] [CrossRef]
- Esler, M.; Esler, D. Can angiotensin receptor-blocking drugs perhaps be harmful in the COVID-19 pandemic? J. Hypertens. 2020, 38, 781–782. [Google Scholar] [CrossRef]
- Fang, L.; Karakiulakis, G.; Roth, M. Are patients with hypertension and diabetes mellitus at increased risk for COVID-19 infection? Lancet Respir. Med. 2020, 8, e21. [Google Scholar] [CrossRef]
- Su, L.; Ma, X.; Yu, H.; Zhang, Z.; Bian, P.; Han, Y.; Sun, J.; Liu, Y.; Yang, C.; Geng, J.; et al. The different clinical characteristics of corona virus disease cases between children and their families in China—The character of children with COVID-19. Emerg. Microbes Infect. 2020, 9, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.C.; Lam, C.W.; Li, A.M.; Wong, C.K.; Cheng, F.W.; Leung, T.F.; Hon, E.K.; Chan, I.H.; Li, C.K.; Fung, K.S.; et al. Inflammatory cytokine profile in children with severe acute respiratory syndrome. Pediatrics 2004, 113, e7–e14. [Google Scholar] [CrossRef] [PubMed]
- Karlberg, J.; Chong, D.S.; Lai, W.Y. Do men have a higher case fatality rate of severe acute respiratory syndrome than women do? Am. J. Epidemiol. 2004, 159, 229–231. [Google Scholar] [CrossRef]
- Gausman, J.; Langer, A. Sex and Gender Disparities in the COVID-19 Pandemic. J. Womens Health (Larchmt) 2020, 29, 465–466. [Google Scholar] [CrossRef]
- Rodriguez-Puertas, R. ACE2 activators for the treatment of COVID 19 patients. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Haschke, M.; Schuster, M.; Poglitsch, M.; Loibner, H.; Salzberg, M.; Bruggisser, M.; Penninger, J.; Krahenbuhl, S. Pharmacokinetics and pharmacodynamics of recombinant human angiotensin-converting enzyme 2 in healthy human subjects. Clin. Pharmacokinet. 2013, 52, 783–792. [Google Scholar] [CrossRef]
- Zhang, H.; Baker, A. Recombinant human ACE2: Acing out angiotensin II in ARDS therapy. Crit. Care 2017, 21, 305. [Google Scholar] [CrossRef]
- Khan, A.; Benthin, C.; Zeno, B.; Albertson, T.E.; Boyd, J.; Christie, J.D.; Hall, R.; Poirier, G.; Ronco, J.J.; Tidswell, M.; et al. A pilot clinical trial of recombinant human angiotensin-converting enzyme 2 in acute respiratory distress syndrome. Crit. Care 2017, 21, 234. [Google Scholar] [CrossRef]
- Mancia, G.; Rea, F.; Ludergnani, M.; Apolone, G.; Corrao, G. Renin-Angiotensin-Aldosterone System Blockers and the Risk of Covid-19. N. Engl. J. Med. 2020, 382, 2431–2440. [Google Scholar] [CrossRef]
- Reynolds, H.R.; Adhikari, S.; Pulgarin, C.; Troxel, A.B.; Iturrate, E.; Johnson, S.B.; Hausvater, A.; Newman, J.D.; Berger, J.S.; Bangalore, S.; et al. Renin-Angiotensin-Aldosterone System Inhibitors and Risk of Covid-19. N. Engl. J. Med. 2020, 382, 2441–2448. [Google Scholar] [CrossRef]
- Mehra, M.R.; Desai, S.S.; Kuy, S.; Henry, T.D.; Patel, A.N. Cardiovascular Disease, Drug Therapy, and Mortality in Covid-19. N. Engl. J. Med. 2020, 382, e102. [Google Scholar] [CrossRef] [PubMed]
- Brojakowska, A.; Narula, J.; Shimony, R.; Bander, J. Clinical Implications of SARS-CoV-2 Interaction With Renin Angiotensin System: JACC Review Topic of the Week. J. Am. Coll. Cardiol. 2020, 75, 3085–3095. [Google Scholar] [CrossRef] [PubMed]
- Vaduganathan, M.; Vardeny, O.; Michel, T.; McMurray, J.J.V.; Pfeffer, M.A.; Solomon, S.D. Renin-Angiotensin-Aldosterone System Inhibitors in Patients with Covid-19. N. Engl. J. Med. 2020, 382, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- South, A.M.; Tomlinson, L.; Edmonston, D.; Hiremath, S.; Sparks, M.A. Controversies of renin-angiotensin system inhibition during the COVID-19 pandemic. Nat. Rev. Nephrol. 2020, 16, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Hatami, N.; Ahi, S.; Sadeghinikoo, A.; Foroughian, M.; Javdani, F.; Kalani, N.; Fereydoni, M.; Keshavarz, P.; Hosseini, A. Worldwide ACE (I/D) polymorphism may affect COVID-19 recovery rate: An ecological meta-regression. Endocrine 2020, 68, 479–484. [Google Scholar] [CrossRef]
- Gupta, R.; Misra, A. COVID19 in South Asians/Asian Indians: Heterogeneity of data and implications for pathophysiology and research. Diabetes Res. Clin. Pract. 2020, 165, 108267. [Google Scholar] [CrossRef]
- Yamamoto, N.; Ariumi, Y.; Nishida, N.; Yamamoto, R.; Bauer, G.; Gojobori, T.; Shimotohno, K.; Mizokami, M. SARS-CoV-2 infections and COVID-19 mortalities strongly correlate with ACE1 I/D genotype. Gene 2020, 758, 144944. [Google Scholar] [CrossRef]
- Delanghe, J.R.; Speeckaert, M.M.; De Buyzere, M.L. ACE Ins/Del genetic polymorphism and epidemiological findings in COVID-19. Clin. Chem. Lab. Med. 2020, 58, 1129–1130. [Google Scholar] [CrossRef]
- Saadat, M. No significant correlation between ACE Ins/Del genetic polymorphism and COVID-19 infection. Clin. Chem. Lab. Med. 2020, 58, 1127–1128. [Google Scholar] [CrossRef]
- Delanghe, J.R.; Speeckaert, M.M.; De Buyzere, M.L. COVID-19 infections are also affected by human ACE1 D/I polymorphism. Clin. Chem. Lab. Med. 2020, 58, 1125–1126. [Google Scholar] [CrossRef] [PubMed]
- Delanghe, J.R.; Speeckaert, M.M.; De Buyzere, M.L. The host’s angiotensin-converting enzyme polymorphism may explain epidemiological findings in COVID-19 infections. Clin. Chim. Acta 2020, 505, 192–193. [Google Scholar] [CrossRef] [PubMed]
- Asselta, R.; Paraboschi, E.M.; Mantovani, A.; Duga, S. ACE2 and TMPRSS2 variants and expression as candidates to sex and country differences in COVID-19 severity in Italy. Aging (Albany N.Y.) 2020, 12, 10087–10098. [Google Scholar] [CrossRef]
- Qu, G.; Li, X.; Hu, L.; Jiang, G. An Imperative Need for Research on the Role of Environmental Factors in Transmission of Novel Coronavirus (COVID-19). Environ. Sci. Technol. 2020, 54, 3730–3732. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.H.; Harada Sassa, M.; Yamamoto, N. Letter to the Editor on “An Imperative Need for Research on the Role of Environmental Factors in Transmission of Novel Coronavirus (COVID-19)”, Back to Basics. Environ. Sci. Technol. 2020, 54, 7738–7739. [Google Scholar] [CrossRef]
- Ehlers, M.R.; Riordan, J.F. Angiotensin-converting enzyme: New concepts concerning its biological role. Biochemistry 1989, 28, 5311–5318. [Google Scholar] [CrossRef]
- Gonzalez-Villalobos, R.A.; Shen, X.Z.; Bernstein, E.A.; Janjulia, T.; Taylor, B.; Giani, J.F.; Blackwell, W.L.; Shah, K.H.; Shi, P.D.; Fuchs, S.; et al. Rediscovering ACE: Novel insights into the many roles of the angiotensin-converting enzyme. J. Mol. Med. (Berl.) 2013, 91, 1143–1154. [Google Scholar] [CrossRef][Green Version]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Li, X.; Rayford, H.; Uhal, B.D. Essential roles for angiotensin receptor AT1a in bleomycin-induced apoptosis and lung fibrosis in mice. Am. J. Pathol. 2003, 163, 2523–2530. [Google Scholar] [CrossRef]
- Uhal, B.D.; Dang, M.T.; Li, X.; Abdul-Hafez, A. Angiotensinogen gene transcription in pulmonary fibrosis. Int. J. Pept. 2012, 2012, 875910. [Google Scholar] [CrossRef][Green Version]
- He, X.; Han, B.; Mura, M.; Xia, S.; Wang, S.; Ma, T.; Liu, M.; Liu, Z. Angiotensin-converting enzyme inhibitor captopril prevents oleic acid-induced severe acute lung injury in rats. Shock 2007, 28, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Gooz, M. ADAM-17: The enzyme that does it all. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 146–169. [Google Scholar] [CrossRef] [PubMed]
- Lambert, D.W.; Yarski, M.; Warner, F.J.; Thornhill, P.; Parkin, E.T.; Smith, A.I.; Hooper, N.M.; Turner, A.J. Tumor necrosis factor-α convertase (ADAM17) mediates regulated ectodomain shedding of the severe-acute respiratory syndrome-coronavirus (SARS-CoV) receptor, angiotensin-converting enzyme-2 (ACE2). J. Biol. Chem. 2005, 280, 30113–30119. [Google Scholar] [CrossRef] [PubMed]
- Sriram, K.; Insel, P.A. Risks of ACE Inhibitor and ARB Usage in COVID-19: Evaluating the Evidence. Clin. Pharmacol. Ther. 2020, 108, 236–241. [Google Scholar] [CrossRef]
- Sriram, K.; Insel, P.A. A hypothesis for pathobiology and treatment of COVID-19: The centrality of ACE1/ACE2 imbalance. Br. J. Pharmacol. 2020. [Google Scholar] [CrossRef]
- Dragonetti, D.; Guarini, G.; Pizzuti, M. Detection of anti-heparin-PF4 complex antibodies in COVID-19 patients on heparin therapy. Blood Transfus 2020, 18, 328. [Google Scholar] [CrossRef]
- Miesbach, W.; Makris, M. COVID-19: Coagulopathy, Risk of Thrombosis, and the Rationale for Anticoagulation. Clin. Appl. Thromb. Hemost. 2020, 26. [Google Scholar] [CrossRef]
- Mazzitelli, M.; Serapide, F.; Tassone, B.; Lagana, D.; Trecarichi, E.M.; Torti, C. Spontaneous and Severe Haematomas in Patients with COVID-19 on Low-Molecular-Weight Heparin for Paroxysmal Atrial Fibrillation. Mediterr. J. Hematol. Infect. Dis. 2020, 12, e2020054. [Google Scholar] [CrossRef]
- Marietta, M.; Vandelli, P.; Mighali, P.; Vicini, R.; Coluccio, V.; D’Amico, R.; Group, C.-H.S. Randomised controlled trial comparing efficacy and safety of high versus low Low-Molecular Weight Heparin dosages in hospitalized patients with severe COVID-19 pneumonia and coagulopathy not requiring invasive mechanical ventilation (COVID-19 HD): A structured summary of a study protocol. Trials 2020, 21, 574. [Google Scholar] [CrossRef]
- Lingamaneni, P.; Gonakoti, S.; Moturi, K.; Vohra, I.; Zia, M. Heparin-Induced Thrombocytopenia in COVID-19. J. Investig. Med. High. Impact. Case Rep. 2020, 8. [Google Scholar] [CrossRef]
- Chi, G.; Lee, J.J.; Jamil, A.; Gunnam, V.; Najafi, H.; Memar Montazerin, S.; Shojaei, F.; Marszalek, J. Venous Thromboembolism among Hospitalized Patients with COVID-19 Undergoing Thromboprophylaxis: A Systematic Review and Meta-Analysis. J. Clin. Med. 2020, 9, 2489. [Google Scholar] [CrossRef]
- Patell, R.; Bogue, T.; Koshy, A.G.; Bindal, P.; Merrill, M.; Aird, W.C.; Bauer, K.A.; Zwicker, J.I. Post-discharge thrombosis and hemorrhage in patients with COVID-19. Blood 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.G.; Wang, Y.; Zhu, H.; Zhao, X. Meta analysis of angiotensin-converting enzyme I/D polymorphism as a risk factor for preeclampsia in Chinese women. Genet. Mol. Res. 2012, 11, 2268–2276. [Google Scholar] [CrossRef] [PubMed]
- Purwaningroom, D.L.P.; Saifurrohman, M.; Widodo, N.; Putri, J.F.; Lukitasari, M. Alteration of Splicing Pattern on Angiotensin Converting Enzyme Gene Due To The Insertion of Alu elements. Int. J. Comput. Biol. 2015, 4, 53–58. [Google Scholar] [CrossRef]
- Widodo; Wisnasari, S.; Saifur Rohman, M.; Yunita, L.; Lukitasari, M.; Nuril, M.; Holil, K.; Purwaningroom, D.L. Alu insertion/deletion of ACE gene polymorphism might not affect significantly the serum bradykinin level in hypertensive patients taking ACE inhibitors. Egypt. J. Med. Hum. Genet. 2017, 18, 187–191. [Google Scholar] [CrossRef]
- Itoyama, S.; Keicho, N.; Quy, T.; Phi, N.C.; Long, H.T.; Ha, L.D.; Ban, V.V.; Ohashi, J.; Hijikata, M.; Matsushita, I.; et al. ACE1 polymorphism and progression of SARS. Biochem. Biophys. Res. Commun. 2004, 323, 1124–1129. [Google Scholar] [CrossRef]
- Li, X.; Sun, X.; Jin, L.; Xue, F. Worldwide spatial genetic structure of angiotensin-converting enzyme gene: A new evolutionary ecological evidence for the thrifty genotype hypothesis. Eur. J. Hum. Genet. 2011, 19, 1002–1008. [Google Scholar] [CrossRef]
- Marshall, R.P.; Webb, S.; Bellingan, G.J.; Montgomery, H.E.; Chaudhari, B.; McAnulty, R.J.; Humphries, S.E.; Hill, M.R.; Laurent, G.J. Angiotensin converting enzyme insertion/deletion polymorphism is associated with susceptibility and outcome in acute respiratory distress syndrome. Am. J. Respir. Crit. Care Med. 2002, 166, 646–650. [Google Scholar] [CrossRef]
- Chiu, R.W.; Tang, N.L.; Hui, D.S.; Chung, G.T.; Chim, S.S.; Chan, K.C.; Sung, Y.M.; Chan, L.Y.; Tong, Y.K.; Lee, W.S.; et al. ACE2 gene polymorphisms do not affect outcome of severe acute respiratory syndrome. Clin. Chem. 2004, 50, 1683–1686. [Google Scholar] [CrossRef][Green Version]
- Chan, K.C.; Tang, N.L.; Hui, D.S.; Chung, G.T.; Wu, A.K.; Chim, S.S.; Chiu, R.W.; Lee, N.; Choi, K.W.; Sung, Y.M.; et al. Absence of association between angiotensin converting enzyme polymorphism and development of adult respiratory distress syndrome in patients with severe acute respiratory syndrome: A case control study. BMC Infect. Dis. 2005, 5, 26. [Google Scholar] [CrossRef]
- Jerng, J.S.; Yu, C.J.; Wang, H.C.; Chen, K.Y.; Cheng, S.L.; Yang, P.C. Polymorphism of the angiotensin-converting enzyme gene affects the outcome of acute respiratory distress syndrome. Crit. Care Med. 2006, 34, 1001–1006. [Google Scholar] [CrossRef] [PubMed]
- Saab, Y.B.; Gard, P.R.; Overall, A.D. The geographic distribution of the ACE II genotype: A novel finding. Genet. Res. 2007, 89, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Pati, A.; Mahto, H.; Padhi, S.; Panda, A.K. ACE deletion allele is associated with susceptibility to SARS-CoV-2 infection and mortality rate: An epidemiological study in the Asian population. Clin. Chim. Acta 2020, 510, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C. ACE-1 I/D Polymorphism Associated with COVID-19 Incidence and Mortality: An Ecological Study. Preprints 2020. [Google Scholar] [CrossRef]
- Yamamoto, N.; Bauer, G. Apparent difference in fatalities between Central Europe and East Asia due to SARS-COV-2 and COVID-19: Four hypotheses for possible explanation. Med. Hypotheses 2020, 144, 110160. [Google Scholar] [CrossRef]
- Smyth, L.J.; Canadas-Garre, M.; Cappa, R.C.; Maxwell, A.P.; McKnight, A.J. Genetic associations between genes in the renin-angiotensin-aldosterone system and renal disease: A systematic review and meta-analysis. BMJ Open 2019, 9, e026777. [Google Scholar] [CrossRef]
- Gemmati, D.; Varani, K.; Bramanti, B.; Piva, R.; Bonaccorsi, G.; Trentini, A.; Manfrinato, M.C.; Tisato, V.; Care, A.; Bellini, T. “Bridging the Gap” Everything that Could Have Been Avoided If We Had Applied Gender Medicine, Pharmacogenetics and Personalized Medicine in the Gender-Omics and Sex-Omics Era. Int. J. Mol. Sci. 2019, 21, 296. [Google Scholar] [CrossRef]
- Cao, Y.; Li, L.; Feng, Z.; Wan, S.; Huang, P.; Sun, X.; Wen, F.; Huang, X.; Ning, G.; Wang, W. Comparative genetic analysis of the novel coronavirus (2019-nCoV/SARS-CoV-2) receptor ACE2 in different populations. Cell Discov. 2020, 6, 11. [Google Scholar] [CrossRef]
- Strafella, C.; Caputo, V.; Termine, A.; Barati, S.; Gambardella, S.; Borgiani, P.; Caltagirone, C.; Novelli, G.; Giardina, E.; Cascella, R. Analysis of ACE2 Genetic Variability among Populations Highlights a Possible Link with COVID-19-Related Neurological Complications. Genes (Basel) 2020, 11, 741. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, Z.; Wang, Y.; Zhou, Y.; Ma, Y.; Zuo, W. Single-cell RNA Expression Profiling of ACE2, The Receptor of SARS-CoV-2. Am. J. Respir. Crit. Care Med. 2020. [Google Scholar] [CrossRef]
- Cai, G. Bulk and single-cell transcriptomics identify tobacco-use disparity in lung gene expression of ACE2, the receptor of 2019-nCov. medRxiv 2020. [Google Scholar] [CrossRef]
- Pinheiro, D.S.; Santos, R.S.; Jardim, P.; Silva, E.G.; Reis, A.A.S.; Pedrino, G.R.; Ulhoa, C.J. The combination of ACE I/D and ACE2 G8790A polymorphisms revels susceptibility to hypertension: A genetic association study in Brazilian patients. PLoS ONE 2019, 14, e0221248. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Li, J.Y.; Wang, C.; Zhang, L.M.; Qiao, H. The ACE2 G8790A Polymorphism: Involvement in Type 2 Diabetes Mellitus Combined with Cerebral Stroke. J. Clin. Lab. Anal. 2017, 31. [Google Scholar] [CrossRef] [PubMed]
- Tognazzo, S.; Gemmati, D.; Palazzo, A.; Catozzi, L.; Carandina, S.; Legnaro, A.; Tacconi, G.; Scapoli, G.L.; Zamboni, P. Prognostic role of factor XIII gene variants in nonhealing venous leg ulcers. J. Vasc. Surg. 2006, 44, 815–819. [Google Scholar] [CrossRef]
- Singh, A.V.; Subhashree, L.; Milani, P.; Gemmati, D.; Zamboni, P. Interplay of iron metallobiology, metalloproteinases, and FXIII, and role of their gene variants in venous leg ulcer. Int. J. Low. Extrem Wounds 2010, 9, 166–179. [Google Scholar] [CrossRef]
- Pascolo, L.; Gianoncelli, A.; Rizzardi, C.; Tisato, V.; Salome, M.; Calligaro, C.; Salvi, F.; Paterson, D.; Zamboni, P. Calcium micro-depositions in jugular truncular venous malformations revealed by Synchrotron-based XRF imaging. Sci. Rep. 2014, 4, 6540. [Google Scholar] [CrossRef]
- Imai, Y.; Kuba, K.; Rao, S.; Huan, Y.; Guo, F.; Guan, B.; Yang, P.; Sarao, R.; Wada, T.; Leong-Poi, H.; et al. Angiotensin-converting enzyme 2 protects from severe acute lung failure. Nature 2005, 436, 112–116. [Google Scholar] [CrossRef]
- Yang, M.; Zhao, J.; Xing, L.; Shi, L. The association between angiotensin-converting enzyme 2 polymorphisms and essential hypertension risk: A meta-analysis involving 14,122 patients. J. Renin Angiotensin Aldosterone Syst. 2015, 16, 1240–1244. [Google Scholar] [CrossRef]
- Li, Y.Y. Lack of Association of ACE2 G8790A Gene Mutation with Essential Hypertension in the Chinese Population: A Meta-Analysis Involving 5260 Subjects. Front. Physiol. 2012, 3, 364. [Google Scholar] [CrossRef]
- Benjafield, A.V.; Wang, W.Y.; Morris, B.J. No association of angiotensin-converting enzyme 2 gene (ACE2) polymorphisms with essential hypertension. Am. J. Hypertens. 2004, 17, 624–628. [Google Scholar] [CrossRef]
- Huang, W.; Yang, W.; Wang, Y.; Zhao, Q.; Gu, D.; Chen, R. Association study of angiotensin-converting enzyme 2 gene (ACE2) polymorphisms and essential hypertension in northern Han Chinese. J. Hum. Hypertens. 2006, 20, 968–971. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhou, J.B.; Yang, J.K. Meta-analysis of association of ACE2 G8790A polymorphism with Chinese Han essential hypertension. J. Renin Angiotensin Aldosterone Syst. 2009, 10, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.; Velkoska, E.; Freeman, M.; Wai, B.; Lancefield, T.F.; Burrell, L.M. From gene to protein-experimental and clinical studies of ACE2 in blood pressure control and arterial hypertension. Front. Physiol. 2014, 5, 227. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Gupta, G.; Mishra, A.; Chellappan, D.K.; Dua, K. Gender and Age Differences Reveal Risk Patterns in COVID-19 Outbreak. Altern. Ther. Health Med. 2020. [Google Scholar]
- Ji, L.D.; Li, J.Y.; Yao, B.B.; Cai, X.B.; Shen, Q.J.; Xu, J. Are genetic polymorphisms in the renin-angiotensin-aldosterone system associated with essential hypertension? Evidence from genome-wide association studies. J. Hum. Hypertens. 2017, 31, 695–698. [Google Scholar] [CrossRef]
- Fan, X.H.; Wang, Y.B.; Wang, H.; Sun, K.; Zhang, W.L.; Song, X.D.; Cheng, J.Z.; Wu, H.Y.; Zhou, X.L.; Hui, R.T. Polymorphisms of angiotensin-converting enzyme (ACE) and ACE2 are not associated with orthostatic blood pressure dysregulation in hypertensive patients. Acta Pharmacol. Sin. 2009, 30, 1237–1244. [Google Scholar] [CrossRef]
- Yi, L.; Gu, Y.H.; Wang, X.L.; An, L.Z.; Xie, X.D.; Shao, W.; Ma, L.Y.; Fang, J.R.; An, Y.D.; Wang, F.; et al. Association of ACE, ACE2 and UTS2 polymorphisms with essential hypertension in Han and Dongxiang populations from north-western China. J. Int. Med. Res. 2006, 34, 272–283. [Google Scholar] [CrossRef]
- Chung, C.M.; Wang, R.Y.; Chen, J.W.; Fann, C.S.; Leu, H.B.; Ho, H.Y.; Ting, C.T.; Lin, T.H.; Sheu, S.H.; Tsai, W.C.; et al. A genome-wide association study identifies new loci for ACE activity: Potential implications for response to ACE inhibitor. Pharm. J. 2010, 10, 537–544. [Google Scholar] [CrossRef]
- Ellinghaus, D.; Degenhardt, F.; Bujanda, L.; Buti, M.; Albillos, A.; Invernizzi, P.; Fernandez, J.; Prati, D.; Baselli, G.; Asselta, R.; et al. Genomewide Association Study of Severe Covid-19 with Respiratory Failure. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Breiman, A.; Ruven-Clouet, N.; Le Pendu, J. Harnessing the natural anti-glycan immune response to limit the transmission of enveloped viruses such as SARS-CoV-2. PLoS Pathog. 2020, 16, e1008556. [Google Scholar] [CrossRef]
- Murray, G.P.; Post, S.R.; Post, G.R. ABO blood group is a determinant of von Willebrand factor protein levels in human pulmonary endothelial cells. J. Clin. Pathol. 2020, 73, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M.; Crestani, S.; Frattini, F.; Sissa, C.; Bonfanti, C. ABO blood group and von Willebrand factor: Biological implications. Clin. Chem. Lab. Med. 2014, 52, 1273–1276. [Google Scholar] [CrossRef] [PubMed]
- Ohira, T.; Cushman, M.; Tsai, M.Y.; Zhang, Y.; Heckbert, S.R.; Zakai, N.A.; Rosamond, W.D.; Folsom, A.R. ABO blood group, other risk factors and incidence of venous thromboembolism: The Longitudinal Investigation of Thromboembolism Etiology (LITE). J. Thromb. Haemost. 2007, 5, 1455–1461. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.Q.; He, F.Z.; Luo, Z.Y.; Wen, J.G.; Wang, L.Y.; Sun, N.L.; Tang, G.F.; Li, Q.; Guo, D.; Liu, Z.Q.; et al. Rs495828 polymorphism of the ABO gene is a predictor of enalapril-induced cough in Chinese patients with essential hypertension. Pharm. Genom. 2014, 24, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Terao, C.; Bayoumi, N.; McKenzie, C.A.; Zelenika, D.; Muro, S.; Mishima, M.; Connell, J.M.; Vickers, M.A.; Lathrop, G.M.; Farrall, M.; et al. Quantitative variation in plasma angiotensin-I converting enzyme activity shows allelic heterogeneity in the ABO blood group locus. Ann. Hum. Genet. 2013, 77, 465–471. [Google Scholar] [CrossRef]
- Yamagata University Genomic Cohort, C. Pleiotropic effect of common variants at ABO Glycosyltranferase locus in 9q32 on plasma levels of pancreatic lipase and angiotensin converting enzyme. PLoS ONE 2014, 9, e55903. [Google Scholar] [CrossRef]
- Guillon, P.; Clement, M.; Sebille, V.; Rivain, J.G.; Chou, C.F.; Ruvoen-Clouet, N.; Le Pendu, J. Inhibition of the interaction between the SARS-CoV spike protein and its cellular receptor by anti-histo-blood group antibodies. Glycobiology 2008, 18, 1085–1093. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Cheng, Q.; Tang, N.L.; Chu, T.T.; Tomlinson, B.; Liu, F.; Kwok, T.C. Gender difference of serum angiotensin-converting enzyme (ACE) activity in DD genotype of ACE insertion/deletion polymorphism in elderly Chinese. J. Renin Angiotensin Aldosterone Syst. 2014, 15, 547–552. [Google Scholar] [CrossRef]
- Zhang, H.; Penninger, J.M.; Li, Y.; Zhong, N.; Slutsky, A.S. Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: Molecular mechanisms and potential therapeutic target. Intensive Care Med. 2020, 46, 586–590. [Google Scholar] [CrossRef]
- Araujo, F.C.; Milsted, A.; Watanabe, I.K.; Del Puerto, H.L.; Santos, R.A.; Lazar, J.; Reis, F.M.; Prokop, J.W. Similarities and differences of X and Y chromosome homologous genes, SRY and SOX3, in regulating the renin-angiotensin system promoters. Physiol. Genom. 2015, 47, 177–186. [Google Scholar] [CrossRef]
- Yuan, X.; Lu, M.L.; Li, T.; Balk, S.P. SRY interacts with and negatively regulates androgen receptor transcriptional activity. J. Biol. Chem. 2001, 276, 46647–46654. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.A.; Orias, M.; Weir, M.R. Novel RAAS agonists and antagonists: Clinical applications and controversies. Nat. Rev. Endocrinol. 2015, 11, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Arendse, L.B.; Danser, A.H.J.; Poglitsch, M.; Touyz, R.M.; Burnett, J.C., Jr.; Llorens-Cortes, C.; Ehlers, M.R.; Sturrock, E.D. Novel Therapeutic Approaches Targeting the Renin-Angiotensin System and Associated Peptides in Hypertension and Heart Failure. Pharmacol. Rev. 2019, 71, 539–570. [Google Scholar] [CrossRef] [PubMed]
- Bakhle, Y.S. How ACE inhibitors transformed the renin-angiotensin system. Br. J. Pharmacol. 2020, 177, 2657–2665. [Google Scholar] [CrossRef]
- Carey, R.M. AT2 Receptors: Potential Therapeutic Targets for Hypertension. Am. J. Hypertens. 2017, 30, 339–347. [Google Scholar] [CrossRef]
- Takezako, T.; Unal, H.; Karnik, S.S.; Node, K. Current topics in angiotensin II type 1 receptor research: Focus on inverse agonism, receptor dimerization and biased agonism. Pharmacol. Res. 2017, 123, 40–50. [Google Scholar] [CrossRef]
- Guo, X.; Zhu, Y.; Hong, Y. Decreased Mortality of COVID-19 With Renin-Angiotensin-Aldosterone System Inhibitors Therapy in Patients With Hypertension: A Meta-Analysis. Hypertension 2020, 76, e13–e14. [Google Scholar] [CrossRef]
- Bonaccorsi, G.; Gambacciani, M.; Gemmati, D. Can estrogens protect against COVID-19? The COVID-19 puzzling and gender medicine. Minerva Ginecol. 2020, 72. [Google Scholar] [CrossRef]
- Debnath, M.; Banerjee, M.; Berk, M. Genetic gateways to COVID-19 infection: Implications for risk, severity, and outcomes. FASEB J. 2020. [Google Scholar] [CrossRef]
- Dongarwar, D.; Salihu, H.M. COVID-19 Pandemic: Marked Global Disparities in Fatalities According to Geographic Location and Universal Health Care. Int. J. MCH AIDS 2020, 9, 213–216. [Google Scholar] [CrossRef]
- Dong, R.; Pei, S.; Yin, C.; He, R.L.; Yau, S.S. Analysis of the Hosts and Transmission Paths of SARS-CoV-2 in the COVID-19 Outbreak. Genes (Basel) 2020, 11, 637. [Google Scholar] [CrossRef]
- Goldstein, J.R.; Atherwood, S. Improved measurement of racial/ethnic disparities in COVID-19 mortality in the United States. medRxiv 2020. [Google Scholar] [CrossRef]
- Nepomuceno, M.R.; Acosta, E.; Alburez-Gutierrez, D.; Aburto, J.M.; Gagnon, A.; Turra, C.M. Besides population age structure, health and other demographic factors can contribute to understanding the COVID-19 burden. Proc. Natl. Acad. Sci. USA 2020, 117, 13881–13883. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Behrman, P.; Dulin, A.; Baskin, M.L.; Buscemi, J.; Alcaraz, K.I.; Goldstein, C.M.; Carson, T.L.; Shen, M.; Fitzgibbon, M. Addressing inequities in COVID-19 morbidity and mortality: Research and policy recommendations. Transl. Behav. Med. 2020, 10, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Helmer, A.; Slater, N.; Smithgall, S. A Review of ACE Inhibitors and ARBs in Black Patients With Hypertension. Ann. Pharmacother. 2018, 52, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Paula, G.H.; Pereira, S.C.; Tanus-Santos, J.E.; Lacchini, R. Pharmacogenomics And Hypertension: Current Insights. Pharmgenom. Pers. Med. 2019, 12, 341–359. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, P.; Gemmati, D. Clinical implications of gene polymorphisms in venous leg ulcer: A model in tissue injury and reparative process. Thromb. Haemost. 2007, 98, 131–137. [Google Scholar]
- Lewis, C.M.; Vassos, E. Polygenic risk scores: From research tools to clinical instruments. Genome Med. 2020, 12, 44. [Google Scholar] [CrossRef]
- Vuille-dit-Bille, R.N.; Camargo, S.M.; Emmenegger, L.; Sasse, T.; Kummer, E.; Jando, J.; Hamie, Q.M.; Meier, C.F.; Hunziker, S.; Forras-Kaufmann, Z.; et al. Human intestine luminal ACE2 and amino acid transporter expression increased by ACE-inhibitors. Amino Acids 2015, 47, 693–705. [Google Scholar] [CrossRef]
- Gasso, P.; Ritter, M.A.; Mas, S.; Lafuente, A. Influence of ABO genotype and phenotype on angiotensin-converting enzyme plasma activity. J. Renin Angiotensin Aldosterone Syst. 2014, 15, 580–584. [Google Scholar] [CrossRef]
- Ling, Y.; van Herpt, T.T.; van Hoek, M.; Dehghan, A.; Hofman, A.; Uitterlinden, A.G.; Jiang, S.; Lieverse, A.G.; Bravenboer, B.; Lu, D.; et al. A genetic variant in SLC6A20 is associated with Type 2 diabetes in white-European and Chinese populations. Diabet. Med. 2014, 31, 1350–1356. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Type of Manuscript | Geographical Regions (Ethnicity) | Association with SARS-CoV-2 Infection and/or COVID-19 Clinical Phenotype | Ref. |
|---|---|---|---|
| Epidemiological investigation | Asian continent | Positive correlation of D-allele with infection and SARS-CoV-2 mortality rates No significant correlation between D-allele with patient recovery rate | [63] |
| Ecological Study (meta-analysis) | Worldwide (50 high income countries) | Positive association of D-allele with cumulative incidence and death rate Possible biological plausibility for the association | [64] |
| Hypothesis paper | Central Europe versus East Asia | ACE1/ACE2 imbalance predicts DD-genotype and D-allele have higher disease prevalence and severity in COVID-19 | [65] |
| Meta-Analysis | Worldwide (30 different countries) | Positive correlation between higher I/D allele frequency ratio and increased disease recovery rate No significant difference in case death rate | [26] |
| Epidemiological investigation | Europe (25 different countries) | Inverse correlation between D-allele frequency with log-transformed prevalence of COVID-19 infections and mortality | [32] |
| Epidemiological investigation | European, North-African and Middle East countries (33 countries) | Negative correlation between log-transformed COVID-19 prevalence and associated mortality with D-allele frequency | [31] |
| Letter to the Editor (Counterpoint) | - | Necessity of considering well-defined geographical spaces to score a polymorphism in a multivariate analysis evaluating its influence on SARS-CoV-2 related infection risk and COVID-19 mortality risk | [29] |
| Epidemiological investigation | European, Middle Eastern, South Asian and East Asian countries | Negative correlation between II-genotype frequency and SARS-CoV-2 infection rate and number of COVID-19 deaths Increasing trend of II-genotype frequency from European to Asian countries | [28] |
| Hypothesis paper | - | Sex differences in I/D allele distribution suggest a higher chance of ACE1/ACE2 imbalance among males during ACE2 receptor suppression due to SARS-CoV-2 infection | [2] |
| Gene | Name | Locus |
|---|---|---|
| ABO | α 1-3-N-acetylgalactosaminyltransferase α 1-3-galactosyltransferase | 9q34.2 |
| ACE1 | Angiotensin I converting enzyme 1 | 17q23.3 |
| ACE2 | Angiotensin I converting enzyme 2 | Xp22.2 |
| ADAM17 | Metallopeptidase domain 17 TNFα-converting enzyme (TACE) | 2p25.1 |
| AGT | Angiotensinogen | 1q42.2 |
| AGTR1 (AT1) | Angiotensin II receptor type 1 | 3q24 |
| AGTR2 (AT2) | Angiotensin II receptor type 2 | Xq23 |
| REN | Renin | 1q32.1 |
| MAS1 | MAS1 proto-oncogene | 6q25.3 |
| SLC6A20 | Solute Carrier Family 6 Member 20 | 3p21.31 |
| SOX3 | SRY-box transcription factor 3 | Xq27.1 |
| SRY | Sex determining region Y | Yp11.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gemmati, D.; Tisato, V. Genetic Hypothesis and Pharmacogenetics Side of Renin-Angiotensin-System in COVID-19. Genes 2020, 11, 1044. https://doi.org/10.3390/genes11091044
Gemmati D, Tisato V. Genetic Hypothesis and Pharmacogenetics Side of Renin-Angiotensin-System in COVID-19. Genes. 2020; 11(9):1044. https://doi.org/10.3390/genes11091044
Chicago/Turabian StyleGemmati, Donato, and Veronica Tisato. 2020. "Genetic Hypothesis and Pharmacogenetics Side of Renin-Angiotensin-System in COVID-19" Genes 11, no. 9: 1044. https://doi.org/10.3390/genes11091044
APA StyleGemmati, D., & Tisato, V. (2020). Genetic Hypothesis and Pharmacogenetics Side of Renin-Angiotensin-System in COVID-19. Genes, 11(9), 1044. https://doi.org/10.3390/genes11091044