Genome-Wide Characterization and Expression Analysis of NHX Gene Family under Salinity Stress in Gossypium barbadense and Its Comparison with Gossypium hirsutum

 ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Characterization of Sodium Proton Antiporters
2.2. Phylogeny and Divergence Analysis
2.3. Promoter and Gene Structure Analysis
2.4. Protein-Protein Interaction and Physical Mapping
2.5. Expression Analysis under Salinity
2.6. RNA Extraction and Quantitative Real-Time PCR Analysis
3. Results
3.1. Characterization of NHX Genes in Cotton Species
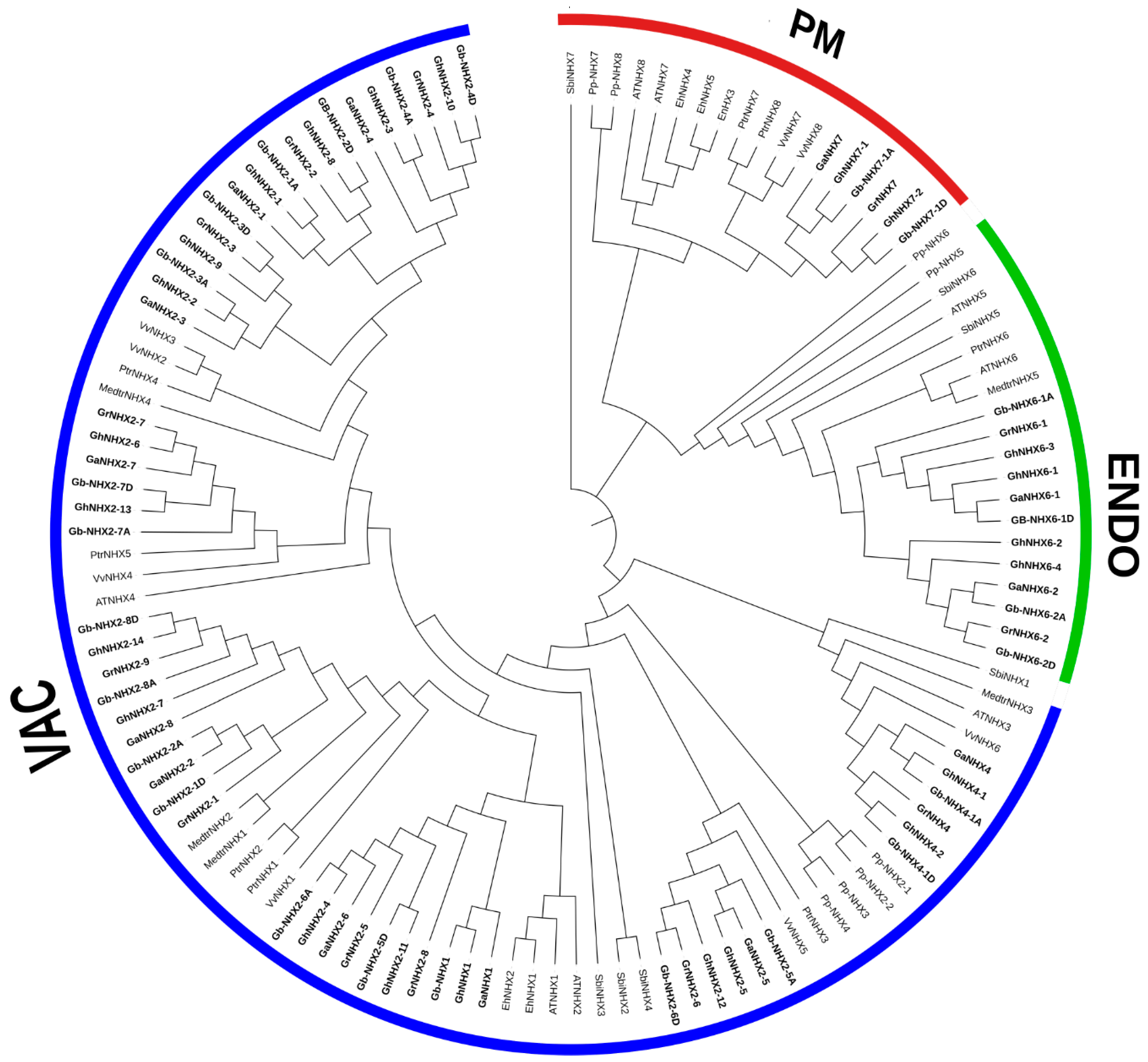
3.2. Phylogeny and Sequence Logos of GbNHX Genes with Different Species
3.3. Comparison of Motifs and Physical Genome Mapping of NHX Genes in G. barbadense and G. hirsutum
3.4. Synteny Analysis and Ka/Ks Ratio of NHX in Cotton Species
3.5. Promoter Analysis of G. barbadense and G. hirsutum NHX Genes
3.6. Expression Pattern of G. barbadense NHX Genes and Its Comparison with G. hirsutum under Salt Stress
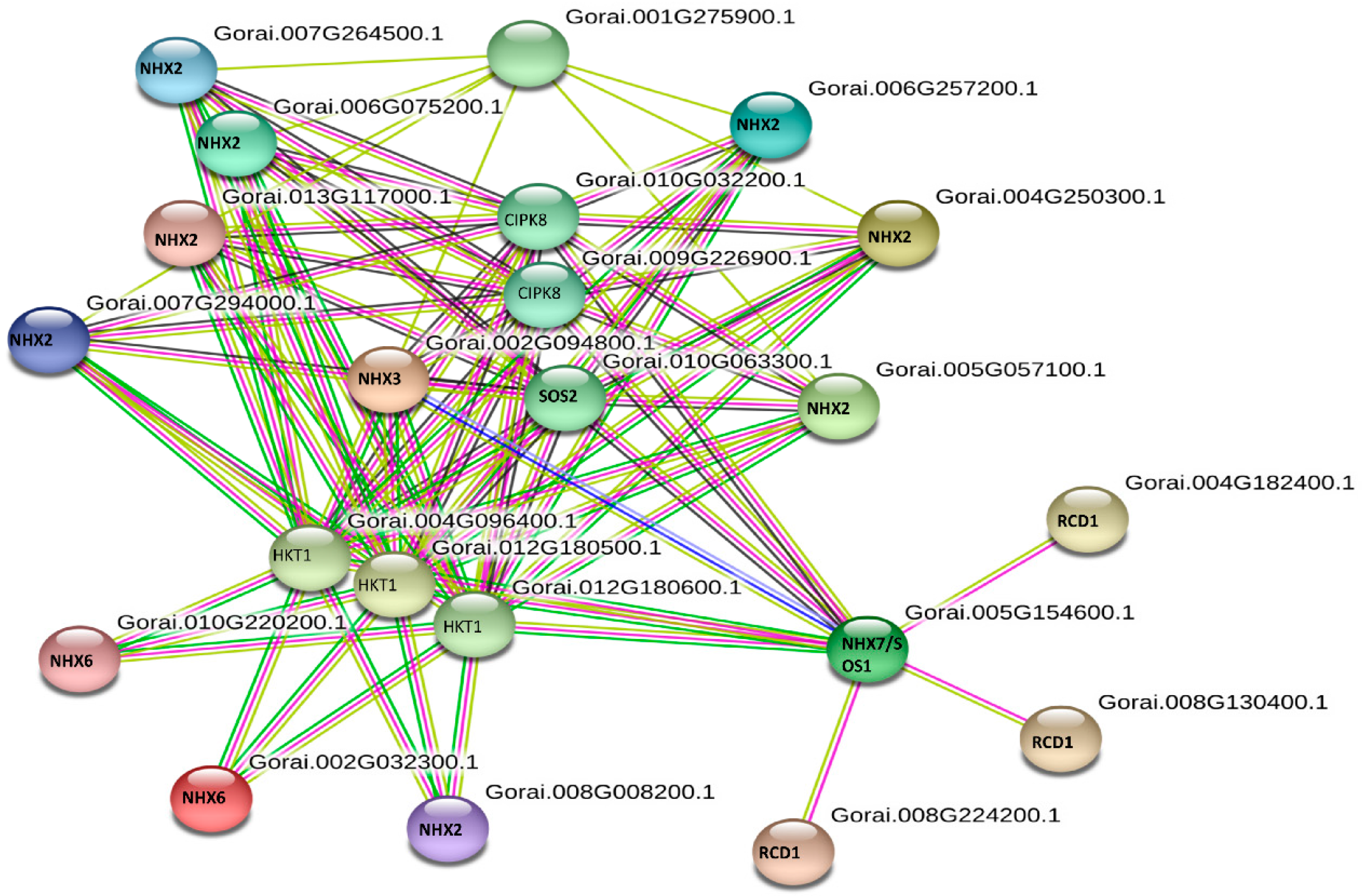
3.7. Protein-Protein Interaction Prediction and GO of GbNHX Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Quesada, V.; Ponce, M.R.; Micol, J.L. Genetic analysis of salt-tolerant mutants in Arabidopsis thaliana. Genetics 2000, 154, 421–436. [Google Scholar] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef]
- Hasegawa, P.M. Sodium (Na+) homeostasis and salt tolerance of plants. Environ. Exp. Bot. 2013, 92, 19–31. [Google Scholar] [CrossRef]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savoure, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Chen, Q.J.; Chai, M.F.; Lu, P.L.; Su, Z.; Qin, Z.X.; Chen, J.; Wang, X.C. AtNHX8, a member of the monovalent cation: Proton antiporter-1 family in Arabidopsis thaliana, encodes a putative Li+/H+ antiporter. Plant J. 2007, 49, 718–728. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Schachtman, D.; Liu, W. Molecular pieces to the puzzle of the interaction between potassium and sodium uptake in plants. Trends Plant Sci. 1999, 4, 281–287. [Google Scholar] [CrossRef]
- Amtmann, A.; Sanders, D. Mechanisms of Na+ uptake by plant cells. In Advances in Botanical Research; Elsevier: York, UK, 1998; Volume 29, pp. 75–112. [Google Scholar]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef] [PubMed]
- Sze, H.; Chanroj, S. Plant endomembrane dynamics: Studies of K+/H+ antiporters provide insights on the effects of pH and ion homeostasis. Plant Physiol. 2018, 177, 875–895. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, X.; Giraldo, J.P.; Shabala, S. It is not all about sodium: Revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1–17. [Google Scholar] [CrossRef]
- Ma, Y.C.; Augé, R.M.; Dong, C.; Cheng, Z.M. Increased salt tolerance with overexpression of cation/proton antiporter 1 genes: A meta-analysis. Plant Biotechnol. J. 2017, 15, 162–173. [Google Scholar] [CrossRef]
- Sharma, H.; Taneja, M.; Upadhyay, S.K. Identification, characterization and expression profiling of cation-proton antiporter superfamily in Triticum aestivum L. and functional analysis of TaNHX4-B. Genomics 2020, 112, 356–370. [Google Scholar] [CrossRef]
- Mäser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.; Sanders, D. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef]
- Brett, C.L.; Donowitz, M.; Rao, R. Evolutionary origins of eukaryotic sodium/proton exchangers. Am. J. Physiol. Cell Physiol. 2005, 288, C223–C239. [Google Scholar] [CrossRef]
- Rodríguez-Rosales, M.P.; Gálvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant Signal. Behav. 2009, 4, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Sardet, C.; Franchi, A.; Pouysségur, J. Molecular cloning, primary structure, and expression of the human growth factor-activatable Na+ H+ antiporter. Cell 1989, 56, 271–280. [Google Scholar] [CrossRef]
- Gaxiola, R.A.; Rao, R.; Sherman, A.; Grisafi, P.; Alper, S.L.; Fink, G.R. The Arabidopsis thaliana proton transporters, AtNhx1 and Avp1, can function in cation detoxification in yeast. Proc. Natl. Acad. Sci. USA 1999, 96, 1480–1485. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Fukada-Tanaka, S.; Inagaki, Y.; Saito, N.; Yonekura-Sakakibara, K.; Tanaka, Y.; Kusumi, T.; Iida, S. Genes encoding the vacuolar Na+/H+ exchanger and flower coloration. Plant Cell Physiol. 2001, 42, 451–461. [Google Scholar] [CrossRef]
- Apse, M.P.; Sottosanto, J.B.; Blumwald, E. Vacuolar cation/H+ exchange, ion homeostasis, and leaf development are altered in a T-DNA insertional mutant of AtNHX1, the Arabidopsis vacuolar Na+/H+ antiporter. Plant J. 2003, 36, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Leidi, E.O.; Barragán, V.; Rubio, L.; El-Hamdaoui, A.; Ruiz, M.T.; Cubero, B.; Fernández, J.A.; Bressan, R.A.; Hasegawa, P.M.; Quintero, F.J. The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of transgenic tomato. Plant J. 2010, 61, 495–506. [Google Scholar] [CrossRef]
- Bowers, K.; Levi, B.P.; Patel, F.I.; Stevens, T.H. The sodium/proton exchanger Nhx1p is required for endosomal protein trafficking in the yeast Saccharomyces cerevisiae. Mol. Biol. Cell 2000, 11, 4277–4294. [Google Scholar] [CrossRef]
- Sottosanto, J.B.; Gelli, A.; Blumwald, E. DNA array analyses of Arabidopsis thaliana lacking a vacuolar Na+/H+ antiporter: Impact of AtNHX1 on gene expression. Plant J. 2004, 40, 752–771. [Google Scholar] [CrossRef]
- Bassil, E.; Ohto, M.-A.; Esumi, T.; Tajima, H.; Zhu, Z.; Cagnac, O.; Belmonte, M.; Peleg, Z.; Yamaguchi, T.; Blumwald, E. The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development. Plant Cell 2011, 23, 224–239. [Google Scholar] [CrossRef]
- Bassil, E.; Tajima, H.; Liang, Y.-C.; Ohto, M.-A.; Ushijima, K.; Nakano, R.; Esumi, T.; Coku, A.; Belmonte, M.; Blumwald, E. Correction: The Arabidopsis Na+/H+ Antiporters NHX1 and NHX2 Control Vacuolar pH and K+ Homeostasis to Regulate Growth, Flower Development, and Reproduction. Plant Cell 2011, 23, 4526. [Google Scholar] [CrossRef]
- Ratner, A.; Jacoby, B. Effect of K+, its counter anion, and pH on sodium efflux from barley root tips. J. Exp. Bot. 1976, 27, 843–852. [Google Scholar] [CrossRef]
- Mennen, H.; Jacoby, B.; Marschner, H. Is sodium proton antiport ubiquitous in plant cells? J. Plant Physiol. 1990, 137, 180–183. [Google Scholar] [CrossRef]
- Allen, G.J.; Sanders, D. Calcineurin, a type 2B protein phosphatase, modulates the Ca2+-permeable slow vacuolar ion channel of stomatal guard cells. Plant Cell 1995, 7, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Blumwald, E.; Poole, R.J. Na+/H+ antiport in isolated tonoplast vesicles from storage tissue of Beta vulgaris. Plant Physiol. 1985, 78, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Garbarino, J.; DuPont, F.M. NaCl induces a Na+/H+ antiport in tonoplast vesicles from barley roots. Plant Physiol. 1988, 86, 231–236. [Google Scholar] [CrossRef]
- Ballesteros, E.; Blumwald, E.; Donaire, J.P.; Belver, A. Na+/H+ antiport activity in tonoplast vesicles isolated from sunflower roots induced by NaCl stress. Physiol. Plant. 1997, 99, 328–334. [Google Scholar] [CrossRef]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.-K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, Q.; Yu, M.; Zhang, Y.; Wu, Y.; Zhang, H. Transgenic salt-tolerant sugar beet (Beta vulgaris L.) constitutively expressing an Arabidopsis thaliana vacuolar Na/H antiporter gene, AtNHX3, accumulates more soluble sugar but less salt in storage roots. Plant Cell Environ. 2008, 31, 1325–1334. [Google Scholar] [CrossRef]
- Ohta, M.; Hayashi, Y.; Nakashima, A.; Hamada, A.; Tanaka, A.; Nakamura, T.; Hayakawa, T. Introduction of a Na+/H+ antiporter gene from Atriplex gmelini confers salt tolerance to rice. FEBS Lett. 2002, 532, 279–282. [Google Scholar] [CrossRef]
- Zhang, H.X.; Blumwald, E. Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol. 2001, 19, 765–768. [Google Scholar] [CrossRef]
- Zhang, Y.-M.; Zhang, H.-M.; Liu, Z.-H.; Li, H.-C.; Guo, X.-L.; Li, G.-L. The wheat NHX antiporter gene TaNHX2 confers salt tolerance in transgenic alfalfa by increasing the retention capacity of intracellular potassium. Plant Mol. Biol. 2015, 87, 317–327. [Google Scholar] [CrossRef]
- Zhang, H.-B.; Li, Y.; Wang, B.; Chee, P.W. Recent advances in cotton genomics. Int. J. Plant Genom. 2008, 2008. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, C.; Yang, X.; Liu, K.; Wu, Z.; Zhang, X.; Zheng, W.; Xun, Q.; Liu, C.; Lu, L. PAG1, a cotton brassinosteroid catabolism gene, modulates fiber elongation. New Phytol. 2014, 203, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Pang, C.; Fan, S.; Song, M.; Yu, J.; Wei, H.; Ma, Q.; Li, L.; Zhang, C.; Yu, S. Genome-wide analysis of the family 1 glycosyltransferases in cotton. Mol. Genet. Genom. 2015, 290, 1805–1818. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, N.; Iqbal, M.Z.; Hussain, A.; Hassan, M. Salt tolerance of cotton (Gossypium hirsutum L.). Asian J. Plant Sci. 2002, 1, 715–719. [Google Scholar]
- Zhang, G.-W.; Lu, H.-L.; Zhang, L.; Chen, B.-L.; Zhou, Z.-G. Salt tolerance evaluation of cotton (Gossypium hirsutum) at its germinating and seedling stages and selection of related indices. Yingyong Shengtai Xuebao 2011, 22, 2045–2053. [Google Scholar] [PubMed]
- Shaheen, H.L.; Iqbal, M.; Azeem, M.; Shahbaz, M.; Shehzadi, M. K-priming positively modulates growth and nutrient status of salt-stressed cotton (Gossypium hirsutum) seedlings. Arch. Agron. Soil Sci. 2016, 62, 759–768. [Google Scholar] [CrossRef]
- Silberbush, M.; Ben-Asher, J. The effect of salinity on parameters of potassium and nitrate uptake of cotton. Commun. Soil Sci. Plan. 1987, 18, 65–81. [Google Scholar] [CrossRef]
- Bernstein, L.; Hayward, H. Physiology of salt tolerance. Annu. Rev. Plant Physiol. 1958, 9, 25–46. [Google Scholar] [CrossRef]
- Peng, J.; Liu, J.; Zhang, L.; Luo, J.; Dong, H.; Ma, Y.; Zhao, X.; Chen, B.; Sui, N.; Zhou, Z. Effects of soil salinity on sucrose metabolism in cotton leaves. PLoS ONE 2016, 11, e0156241. [Google Scholar] [CrossRef]
- Longenecker, D. The influence of high sodium in soils upon fruiting and shedding, boll characteristics, fiber properties, and yields of two cotton species. Soil Sci. 1974, 118, 387–396. [Google Scholar] [CrossRef]
- Satir, O.; Berberoglu, S. Crop yield prediction under soil salinity using satellite derived vegetation indices. Field Crops Res. 2016, 192, 134–143. [Google Scholar] [CrossRef]
- Li, F.; Fan, G.; Lu, C.; Xiao, G.; Zou, C.; Kohel, R.J.; Ma, Z.; Shang, H.; Ma, X.; Wu, J. Genome sequence of cultivated Upland cotton (Gossypium hirsutum TM-1) provides insights into genome evolution. Nat. Biotechnol. 2015, 33, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fan, G.; Wang, K.; Sun, F.; Yuan, Y.; Song, G.; Li, Q.; Ma, Z.; Lu, C.; Zou, C. Genome sequence of the cultivated cotton Gossypium arboreum. Nat. Genet. 2014, 46, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Huang, G.; He, S.; Yang, Z.; Sun, G.; Ma, X.; Li, N.; Zhang, X.; Sun, J.; Liu, M. Resequencing of 243 diploid cotton accessions based on an updated A genome identifies the genetic basis of key agronomic traits. Nat. Genet. 2018, 50, 796–802. [Google Scholar] [CrossRef]
- Wang, K.; Wang, Z.; Li, F.; Ye, W.; Wang, J.; Song, G.; Yue, Z.; Cong, L.; Shang, H.; Zhu, S. The draft genome of a diploid cotton Gossypium raimondii. Nat. Genet. 2012, 44, 1098–1103. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, B.; Zheng, H.-J.; Hu, Y.; Lu, G.; Yang, C.-Q.; Chen, J.-D.; Chen, J.-J.; Chen, D.-Y.; Zhang, L. Gossypium barbadense genome sequence provides insight into the evolution of extra-long staple fiber and specialized metabolites. Sci. Rep. 2015, 5, 14139. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2015, 44, D279–D285. [Google Scholar] [CrossRef]
- Zhu, T.; Liang, C.; Meng, Z.; Sun, G.; Meng, Z.; Guo, S.; Zhang, R. CottonFGD: An integrated functional genomics database for cotton. BMC Plant Biol. 2017, 17, 1–9. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Moller, S.; Croning, M.D.; Apweiler, R. Evaluation of methods for the prediction of membrane spanning regions. Bioinformatics 2001, 17, 646–653. [Google Scholar] [CrossRef]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins: Struct. Funct. Bioinform. 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Hima Kumari, P.; Anil Kumar, S.; Ramesh, K.; Sudhakar Reddy, P.; Nagaraju, M.; Bhanu Prakash, A.; Shah, T.; Henderson, A.; Srivastava, R.K.; Rajasheker, G. Genome-wide identification and analysis of Arabidopsis sodium proton antiporter (NHX) and human sodium proton exchanger (NHE) homologs in sorghum bicolor. Genes 2018, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Taji, T.; Sakurai, T.; Mochida, K.; Ishiwata, A.; Kurotani, A.; Totoki, Y.; Toyoda, A.; Sakaki, Y.; Seki, M.; Ono, H. Large-scale collection and annotation of full-length enriched cDNAs from a model halophyte, Thellungiella halophila. BMC Plant Biol. 2008, 8, 115. [Google Scholar] [CrossRef]
- Tian, F.; Chang, E.; Li, Y.; Sun, P.; Hu, J.; Zhang, J. Expression and integrated network analyses revealed functional divergence of NHX-type Na+/H+ exchanger genes in poplar. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef]
- Sandhu, D.; Pudussery, M.V.; Kaundal, R.; Suarez, D.L.; Kaundal, A.; Sekhon, R.S. Molecular characterization and expression analysis of the Na+/H+ exchanger gene family in Medicago truncatula. Funct. Integr. Genom. 2018, 18, 141–153. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools—An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020. [Google Scholar] [CrossRef] [PubMed]
- Suyama, M.; Torrents, D.; Bork, P. PAL2NAL: Robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res. 2006, 34, W609–W612. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Zhu, Q.; Chen, X.; Luo, J. GSDS: A gene structure display server. Yi Chuan = Hereditas 2007, 29, 1023–1026. [Google Scholar] [CrossRef]
- Artico, S.; Nardeli, S.M.; Brilhante, O.; Grossi-de-Sa, M.F.; Alves-Ferreira, M. Identification and evaluation of new reference genes in Gossypium hirsutum for accurate normalization of real-time quantitative RT-PCR data. BMC Plant Biol. 2010, 10, 49. [Google Scholar] [CrossRef]
- Paterson, A.H.; Wendel, J.F.; Gundlach, H.; Guo, H.; Jenkins, J.; Jin, D.; Llewellyn, D.; Showmaker, K.C.; Shu, S.; Udall, J. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature 2012, 492, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.J.; Schmelzer, E.; Hahlbrock, K.; Somssich, I.E. Early nuclear events in plant defence signalling: Rapid gene activation by WRKY transcription factors. EMBO J. 1999, 18, 4689–4699. [Google Scholar] [CrossRef]
- Abul-Naas, A.A.; Omran, M.S. Salt tolerance of seventeen cotton cultivars during germination and early seedling development. Z Acker Pflanzenbau 1975, 140, 229–236. [Google Scholar]
- Ashraf, M. Salt tolerance of cotton: Some new advances. Crit. Rev. Plant Sci. 2002, 21, 1–30. [Google Scholar] [CrossRef]
- Witt, T.W.; Ulloa, M.; Schwartz, R.C.; Ritchie, G.L. Response to deficit irrigation of morphological, yield and fiber quality traits of upland (Gossypium hirsutum L.) and Pima (G. barbadense L.) cotton in the Texas High Plains. Field Crop. Res. 2020, 249, 107759. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, J.; Fang, L.; Zhang, Z.; Ma, W.; Niu, Y.; Ju, L.; Deng, J.; Zhao, T.; Lian, J. Gossypium barbadense and Gossypium hirsutum genomes provide insights into the origin and evolution of allotetraploid cotton. Nat. Genet. 2019, 51, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, J.; Grinstein, S. Diversity of the mammalian sodium/proton exchanger SLC9 gene family. Pflug. Arch. 2004, 447, 549–565. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Quintero, F.J.; Pardo, J.M.; Zhu, J.K. The putative plasma membrane Na(+)/H(+) antiporter SOS1 controls long-distance Na(+) transport in plants. Plant Cell 2002, 14, 465–477. [Google Scholar] [CrossRef]
- Wu, G.-Q.; Wang, J.-L.; Li, S.-J. Genome-wide identification of Na+/H+ antiporter (NHX) genes in sugar beet (Beta vulgaris L.) and their regulated expression under salt stress. Genes 2019, 10, 401. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, J.L.; Aronson, P.S. Amiloride inhibition of the Na+-H+ exchanger in renal microvillus membrane vesicles. Am. J. Physiol. 1981, 241, F374–F379. [Google Scholar] [CrossRef]
- Blumwald, E.; Poole, R.J. Salt tolerance in suspension cultures of sugar beet: Induction of na/h antiport activity at the tonoplast by growth in salt. Plant Physiol. 1987, 83, 884–887. [Google Scholar] [CrossRef]
- Counillon, L.; Franchi, A.; Pouyssegur, J. A point mutation of the Na+/H+ exchanger gene (NHE1) and amplification of the mutated allele confer amiloride resistance upon chronic acidosis. Proc. Natl. Acad. Sci. USA 1993, 90, 4508–4512. [Google Scholar] [CrossRef]
- Rong, J.; Feltus, F.A.; Liu, L.; Lin, L.; Paterson, A.H. Gene copy number evolution during tetraploid cotton radiation. Heredity 2010, 105, 463–472. [Google Scholar] [CrossRef]
- Conant, G.C.; Wolfe, K.H. Turning a hobby into a job: How duplicated genes find new functions. Nat Rev. Genet. 2008, 9, 938–950. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, J.; Liu, H.; Li, T.; Wang, K.; Hao, C.; Liu, H.; Zhang, X. TaBT1, affecting starch synthesis and thousand kernel weight, underwent strong selection during wheat improvement. J. Exp. Bot. 2019, 70, 1497–1511. [Google Scholar] [CrossRef] [PubMed]
- Bowers, J.E.; Chapman, B.A.; Rong, J.; Paterson, A.H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Paz-Ares, J. MYB transcription factors in plants. Trends Genet. 1997, 13, 67–73. [Google Scholar] [CrossRef]
- Liu, L.; Xu, W.; Hu, X.; Liu, H.; Lin, Y. W-box and G-box elements play important roles in early senescence of rice flag leaf. Sci. Rep. 2016, 6, 20881. [Google Scholar] [CrossRef]
- Venema, K.; Quintero, F.J.; Pardo, J.M.; Donaire, J.P. The Arabidopsis Na+/H+ exchanger AtNHX1 catalyzes low affinity Na+ and K+ transport in reconstituted liposomes. J. Biol. Chem. 2002, 277, 2413–2418. [Google Scholar] [CrossRef]
- Xia, T.; Apse, M.P.; Aharon, G.S.; Blumwald, E. Identification and characterization of a NaCl-inducible vacuolar Na+/H+ antiporter in Beta vulgaris. Physiol. Plant. 2002, 116, 206–212. [Google Scholar] [CrossRef]
- Rodríguez-Rosales, M.P.; Jiang, X.; Gálvez, F.J.; Aranda, M.N.; Cubero, B.; Venema, K. Overexpression of the tomato K+/H+ antiporter LeNHX2 confers salt tolerance by improving potassium compartmentalization. New Phytol. 2008, 179, 366–377. [Google Scholar] [CrossRef]
- Vasekina, A.; Yershov, P.; Reshetova, O.; Tikhonova, T.; Lunin, V.; Trofimova, M.; Babakov, A. Vacuolar Na+/H+ antiporter from barley: Identification and response to salt stress. Biochemistry (Moscow) 2005, 70, 100–107. [Google Scholar] [CrossRef]
- Zörb, C.; Noll, A.; Karl, S.; Leib, K.; Yan, F.; Schubert, S. Molecular characterization of Na+/H+ antiporters (ZmNHX) of maize (Zea mays L.) and their expression under salt stress. J. Plant Physiol. 2005, 162, 55–66. [Google Scholar] [CrossRef]
- Brini, F.; Gaxiola, R.A.; Berkowitz, G.A.; Masmoudi, K. Cloning and characterization of a wheat vacuolar cation/proton antiporter and pyrophosphatase proton pump. Plant Physiol. Biochem. 2005, 43, 347–354. [Google Scholar] [CrossRef]
- Li, W.Y.F.; Wong, F.L.; Tsai, S.N.; Phang, T.H.; Shao, G.; Lam, H.M. Tonoplast-located GmCLC1 and GmNHX1 from soybean enhance NaCl tolerance in transgenic bright yellow (BY)-2 cells. Plant Cell Environ. 2006, 29, 1122–1137. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Li, Q.; Wang, H.; Zhang, J.; Du, J.; Feng, H.; Blumwald, E.; Yu, L.; Xu, G. Two NHX-type transporters from Helianthus tuberosus improve the tolerance of rice to salinity and nutrient deficiency stress. Plant Biotechnol. J. 2018, 16, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, A.; Nakamura, A.; Hara, N.; Toki, S.; Tanaka, Y. Molecular and functional analyses of rice NHX-type Na+/H+ antiporter genes. Planta 2011, 233, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, J.; Zhong, Y.; Geng, F.; Cramer, G.R.; Cheng, Z.-M.M. Subfunctionalization of cation/proton antiporter 1 genes in grapevine in response to salt stress in different organs. Hortic. Res. 2015, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bao, A.K.; Du, B.Q.; Touil, L.; Kang, P.; Wang, Q.L.; Wang, S.M. Co-expression of tonoplast Cation/H+ antiporter and H+-pyrophosphatase from xerophyte Zygophyllum xanthoxylum improves alfalfa plant growth under salinity, drought and field conditions. Plant Biotechnol. J. 2016, 14, 964–975. [Google Scholar] [CrossRef]
- Wang, B.; Zhai, H.; He, S.; Zhang, H.; Ren, Z.; Zhang, D.; Liu, Q. A vacuolar Na+/H+ antiporter gene, IbNHX2, enhances salt and drought tolerance in transgenic sweetpotato. Sci. Hortic. 2016, 201, 153–166. [Google Scholar] [CrossRef]
- Lu, W.; Guo, C.; Li, X.; Duan, W.; Ma, C.; Zhao, M.; Gu, J.; Du, X.; Liu, Z.; Xiao, K. Overexpression of TaNHX3, a vacuolar Na+/H+ antiporter gene in wheat, enhances salt stress tolerance in tobacco by improving related physiological processes. Plant Physiol. Biochem. 2014, 76, 17–28. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Yadav, N.S.; Shukla, P.S.; Jha, A.; Agarwal, P.K.; Jha, B. The SbSOS1 gene from the extreme halophyte Salicornia brachiata enhances Na+ loading in xylem and confers salt tolerance in transgenic tobacco. BMC Plant Biol. 2012, 12, 188. [Google Scholar] [CrossRef]
- Zhang, W.-D.; Wang, P.; Bao, Z.; Ma, Q.; Duan, L.-J.; Bao, A.-K.; Zhang, J.-L.; Wang, S.-M. SOS1, HKT1; 5, and NHX1 synergistically modulate Na+ homeostasis in the halophytic grass Puccinellia tenuiflora. Front. Plant Sci. 2017, 8, 576. [Google Scholar]
- Ma, Q.; Li, Y.-X.; Yuan, H.-J.; Hu, J.; Wei, L.; Bao, A.-K.; Zhang, J.-L.; Wang, S.-M. ZxSOS1 is essential for long-distance transport and spatial distribution of Na+ and K+ in the xerophyte Zygophyllum xanthoxylum. Plant Soil 2014, 374, 661–676. [Google Scholar] [CrossRef]
- Quintero, F.J.; Martinez-Atienza, J.; Villalta, I.; Jiang, X.; Kim, W.-Y.; Ali, Z.; Fujii, H.; Mendoza, I.; Yun, D.-J.; Zhu, J.-K. Activation of the plasma membrane Na/H antiporter Salt-Overly-Sensitive 1 (SOS1) by phosphorylation of an auto-inhibitory C-terminal domain. Proc. Natl. Acad. Sci. USA 2011, 108, 2611–2616. [Google Scholar] [CrossRef] [PubMed]
- Katiyar-Agarwal, S.; Zhu, J.; Kim, K.; Agarwal, M.; Fu, X.; Huang, A.; Zhu, J.-K. The plasma membrane Na+/H+ antiporter SOS1 interacts with RCD1 and functions in oxidative stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 18816–18821. [Google Scholar] [CrossRef] [PubMed]
- Weinl, S.; Kudla, J. The CBL–CIPK Ca2+-decoding signaling network: Function and perspectives. New Phytol. 2009, 184, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-Y.; Ali, Z.; Park, H.J.; Park, S.J.; Cha, J.-Y.; Perez-Hormaeche, J.; Quintero, F.J.; Shin, G.; Kim, M.R.; Qiang, Z. Release of SOS2 kinase from sequestration with GIGANTEA determines salt tolerance in Arabidopsis. Nat. Commun. 2013, 4, 1–13. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Protein (aa) | CDS (bp) | MW (kDa) | pI | Localization | Na+/H+ Exchanger Domain (Start–End) |
|---|---|---|---|---|---|---|---|
| Gb-NHX1 | Gbar_D07G012760 | 164 | 1455 | 18.666 | 10.438 | Vac | Nil |
| Gb-NHX2-1A | Gbar_A02G004630 | 535 | 1527 | 59.063 | 8.663 | Vac | 22–437 |
| Gb-NHX2-2A | Gbar_A08G023430 | 320 | 1608 | 35.548 | 10.181 | Vac | 7–212 |
| Gb-NHX2-3A | Gbar_A09G007060 | 542 | 3459 | 59.844 | 7.858 | Vac | 29–444 |
| Gb-NHX2-4A | Gbar_A09G025000 | 541 | 1587 | 59.721 | 9.05 | Vac | 25–445 |
| Gb-NHX2-5A | Gbar_A11G028010 | 540 | 963 | 59.586 | 6.989 | Vac | 16–440 |
| Gb-NHX2-6A | Gbar_A11G025170 | 543 | 1629 | 60.106 | 7.657 | Vac | 24–445 |
| Gb-NHX2-7A | Gbar_A12G000720 | 445 | 1626 | 49.371 | 8.063 | Vac | 17–420 |
| Gb-NHX2-8A | Gbar_A13G011300 | 541 | 1632 | 59.706 | 8.552 | Vac | 25–444 |
| Gb-NHX4-1A | Gbar_A01G007690 | 508 | 1623 | 56.977 | 7.591 | Vac | 16–426 |
| Gb-NHX6-1A | Gbar_A01G002880 | 484 | 1338 | 53.246 | 6.795 | Endo | 25–433 |
| Gb-NHX6-2A | Gbar_A06G019530 | 528 | 1626 | 58.41 | 5.514 | Vac | 25–437 |
| Gb-NHX7-1A | Gbar_A03G012870 | 1152 | 1584 | 128.07 | 6.878 | PM | 29–445 |
| Gb-NHX2-1D | Gbar_D08G024100 | 551 | 1578 | 61.315 | 8.457 | Vac | 30–434 |
| GB-NHX2-2D | Gbar_D02G005160 | 535 | 1608 | 59.178 | 8.453 | Vac | 22–437 |
| Gb-NHX2-3D | Gbar_D09G006790 | 542 | 3459 | 60.015 | 8.731 | Vac | 29–444 |
| Gb-NHX2-4D | Gbar_D09G024630 | 541 | 1572 | 59.705 | 9.175 | Vac | 29–444 |
| Gb-NHX2-5D | Gbar_D11G026100 | 497 | 495 | 55.119 | 7.009 | Vac | 4–406 |
| Gb-NHX2-6D | Gbar_D11G028500 | 542 | 1656 | 59.758 | 6.42 | Vac | 19–448 |
| Gb-NHX2-7D | Gbar_D12G000860 | 525 | 1629 | 58.126 | 8.549 | Vac | 25–444 |
| Gb-NHX2-8D | Gbar_D13G011070 | 541 | 1626 | 59.715 | 8.554 | Vac | 31–442 |
| Gb-NHX4-1D | Gbar_D01G007950 | 525 | 1494 | 59.139 | 7.62 | Vac | 21–441 |
| GB-NHX6-1D | Gbar_D01G003050 | 527 | 1629 | 58.056 | 5.978 | Endo | 28–437 |
| Gb-NHX6-2D | Gbar_D06G020390 | 523 | 1578 | 57.72 | 5.494 | Endo | 28–432 |
| Gb-NHX7-1D | Gbar_D02G014810 | 1152 | 1626 | 128.14 | 6.764 | PM | 31–443 |
| Chromosome | G. arboreum | G. raimondii | G. barbadense | G. hirsutum |
|---|---|---|---|---|
| A01 | Ga_NHX6-1 | Gb-NHX6-1A | Gh_NHX6-1A | |
| A01 | Ga_NHX4 | Gb-NHX4-1A | Gh_NHX4-1A | |
| A02 | Gb-NHX2-1A | Gh_NHX2-1A | ||
| A03 | Ga_NHX2-1 | |||
| A03 | Ga_NHX7 | Gb-NHX7-1A | Gh_NHX7-1A | |
| A06 | Ga_NHX1 | |||
| A06 | Ga_NHX6-2 | Gb-NHX6-2A | Gh_NHX6-2A | |
| A08 | Ga_NHX2-2 | Gb-NHX2-2A | ||
| A09 | Ga_NHX2-3 | Gb-NHX2-3A | Gh_NHX2-2A | |
| A09 | Ga_NHX2-4 | Gb-NHX2-4A | Gh_NHX2-3A | |
| A11 | Ga_NHX2-5 | Gb-NHX2-6A | Gh_NHX2-4A | |
| A11 | Ga_NHX2-6 | Gb-NHX2-5A | Gh_NHX2-5A | |
| A12 | Ga_NHX2-7 | Gb-NHX2-7A | Gh_NHX2-6A | |
| A13 | Ga_NHX2-8 | Gb-NHX2-8A | Gh_NHX2-7A | |
| D01 | Gb-NHX6-1D | Gh_NHX6-1D | ||
| D01 | Gb-NHX4-1D | Gh_NHX4-1D | ||
| D02 | Gr_NHX6-1 | Gb-NHX2-2D | Gh_NHX2-1D | |
| D02 | Gr_NHX4 | Gb-NHX7-1D | Gh_NHX7-1D | |
| D04 | Gr_NHX2-1 | |||
| D05 | Gr_NHX2-2 | |||
| D05 | Gr_NHX7 | |||
| D06 | Gr_NHX2-3 | Gb-NHX6-2D | Gh_NHX6-2D | |
| D06 | Gr_NHX2-4 | |||
| D07 | Gr_NHX2-5 | Gb-NHX1 | Gh_NHX1 | |
| D07 | Gr_NHX2-6 | |||
| D08 | Gr_NHX2-7 | Gb-NHX2-1D | ||
| D09 | Gb-NHX2-3D | Gh_NHX2-2D | ||
| D09 | Gb-NHX2-4D | Gh_NHX2-3D | ||
| D10 | Gr_NHX2-8 | |||
| D10 | Gr_NHX6-2 | |||
| D11 | Gb-NHX2-5D | Gh_NHX2-4D | ||
| D11 | Gb-NHX2-6D | Gh_NHX2-5D | ||
| D12 | Gb-NHX2-7D | Gh_NHX2-6D | ||
| D13 | Gr_NHX2-9 | Gb-NHX2-8D | Gh_NHX2-7D |
| G. barbadense Orthologous/Paralogous | G. hirsutum Orthologous/Paralogous | ||
|---|---|---|---|
| Gene ID | Gene ID | Gene ID | Gene ID |
| Gb-NHX6-1A | GB-NHX6-1D | Gh_NHX6-1 | Gh_NHX2-2 |
| Gb-NHX4-1A | Gb-NHX4-1D | Gh_NHX6-1 | Gh_NHX2-3 |
| Gb-NHX2-1A | Gb-NHX2-4A | Gh_NHX6-1 | Gh_NHX6-3 |
| Gb-NHX2-1A | Gb-NHX2-3A | Gh_NHX6-1 | Gh_NHX2-9 |
| Gb-NHX2-1A | GB-NHX2-2D | Gh_NHX6-1 | Gh_NHX2-10 |
| Gb-NHX2-1A | Gb-NHX2-4D | Gh_NHX4-1 | Gh_NHX4-2 |
| Gb-NHX2-1A | Gb-NHX2-3D | Gh_NHX2-1 | Gh_NHX2-2 |
| Gb-NHX7-1A | Gb-NHX7-1D | Gh_NHX2-1 | Gh_NHX2-3 |
| Gb-NHX6-2A | Gb-NHX6-2D | Gh_NHX2-1 | Gh_NHX2-6 |
| Gb-NHX2-2A | Gb-NHX2-6A | Gh_NHX2-1 | Gh_NHX2-8 |
| Gb-NHX2-2A | Gb-NHX2-1D.1 | Gh_NHX2-1 | Gh_NHX2-10 |
| Gb-NHX2-2A | Gb-NHX2-8D.1 | Gh_NHX2-1 | Gh_NHX2-9 |
| Gb-NHX2-4A | GB-NHX2-2D | Gh_NHX7-1 | Gh_NHX7-2 |
| Gb-NHX2-3A | GB-NHX2-2D | Gh_NHX2-2 | Gh_NHX2-3 |
| Gb-NHX2-4A | Gb-NHX2-4D | Gh_NHX2-2 | Gh_NHX2-8 |
| Gb-NHX2-3A | Gb-NHX2-3D | Gh_NHX2-2 | Gh_NHX6-4 |
| Gb-NHX2-6A | Gb-NHX2-8A | Gh_NHX2-2 | Gh_NHX2-9 |
| Gb-NHX2-6A | Gb-NHX2-1D | Gh_NHX2-3 | Gh_NHX2-8 |
| Gb-NHX2-6A | Gb-NHX2-5D | Gh_NHX2-3 | Gh_NHX2-10 |
| Gb-NHX2-5A | Gb-NHX2-6D | Gh_NHX2-3 | Gh_NHX2-9 |
| Gb-NHX2-6A | Gb-NHX2-8D | Gh_NHX2-4 | Gh_NHX2-7 |
| Gb-NHX2-7A | Gb-NHX2-7D | Gh_NHX2-4 | Gh_NHX2-11 |
| Gb-NHX2-8A | Gb-NHX2-1D | Gh_NHX2-4 | Gh_NHX2-14 |
| Gb-NHX2-8A | Gb-NHX2-5D | Gh_NHX2-5 | Gh_NHX2-12 |
| Gb-NHX2-8A | Gb-NHX2-8D | Gh_NHX2-6 | Gh_NHX2-8 |
| GB-NHX2-2D | Gb-NHX2-4D | Gh_NHX2-7 | Gh_NHX2-11 |
| GB-NHX2-2D | Gb-NHX2-3D | Gh_NHX2-7 | Gh_NHX2-14 |
| Gb-NHX2-1D | Gb-NHX2-5D | Gh_NHX2-8 | Gh_NHX2-10 |
| Gb-NHX2-1D | Gb-NHX2-8D | Gh_NHX2-8 | Gh_NHX2-9 |
| Gb-NHX2-5D | Gb-NHX2-8D | Gh_NHX6-4 | Gh_NHX2-9 |
| Gh_NHX2-11 | Gh_NHX2-14 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akram, U.; Song, Y.; Liang, C.; Abid, M.A.; Askari, M.; Myat, A.A.; Abbas, M.; Malik, W.; Ali, Z.; Guo, S.; et al. Genome-Wide Characterization and Expression Analysis of NHX Gene Family under Salinity Stress in Gossypium barbadense and Its Comparison with Gossypium hirsutum. Genes 2020, 11, 803. https://doi.org/10.3390/genes11070803
Akram U, Song Y, Liang C, Abid MA, Askari M, Myat AA, Abbas M, Malik W, Ali Z, Guo S, et al. Genome-Wide Characterization and Expression Analysis of NHX Gene Family under Salinity Stress in Gossypium barbadense and Its Comparison with Gossypium hirsutum. Genes. 2020; 11(7):803. https://doi.org/10.3390/genes11070803
Chicago/Turabian StyleAkram, Umar, Yuhan Song, Chengzhen Liang, Muhammad Ali Abid, Muhammad Askari, Aye Aye Myat, Mubashir Abbas, Waqas Malik, Zulfiqar Ali, Sandui Guo, and et al. 2020. "Genome-Wide Characterization and Expression Analysis of NHX Gene Family under Salinity Stress in Gossypium barbadense and Its Comparison with Gossypium hirsutum" Genes 11, no. 7: 803. https://doi.org/10.3390/genes11070803
APA StyleAkram, U., Song, Y., Liang, C., Abid, M. A., Askari, M., Myat, A. A., Abbas, M., Malik, W., Ali, Z., Guo, S., Zhang, R., & Meng, Z. (2020). Genome-Wide Characterization and Expression Analysis of NHX Gene Family under Salinity Stress in Gossypium barbadense and Its Comparison with Gossypium hirsutum. Genes, 11(7), 803. https://doi.org/10.3390/genes11070803