Comparative Transcriptome Analysis Suggests Key Roles for 5-Hydroxytryptamlne Receptors in Control of Goose Egg Production

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
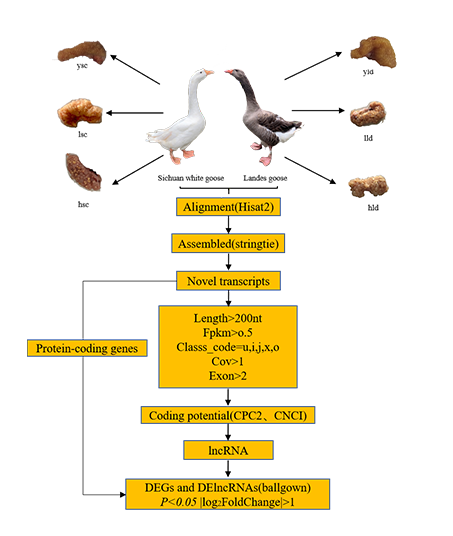
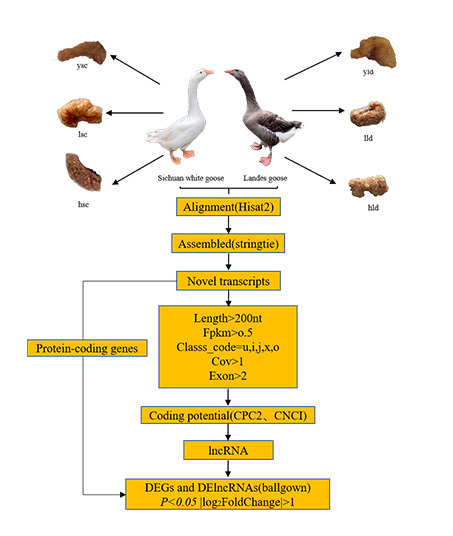
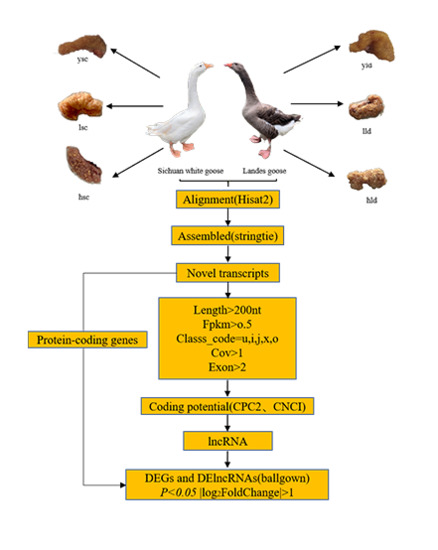
2.2. Experimental Design and Sample Collection
2.3. RNA Isolation and Sequencing
2.4. Transcriptome Alignment and Assembly
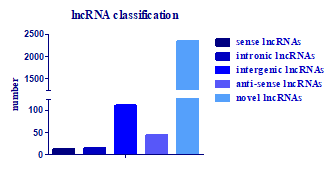
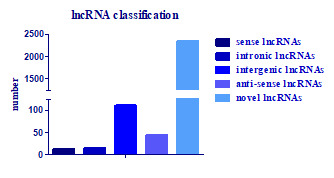
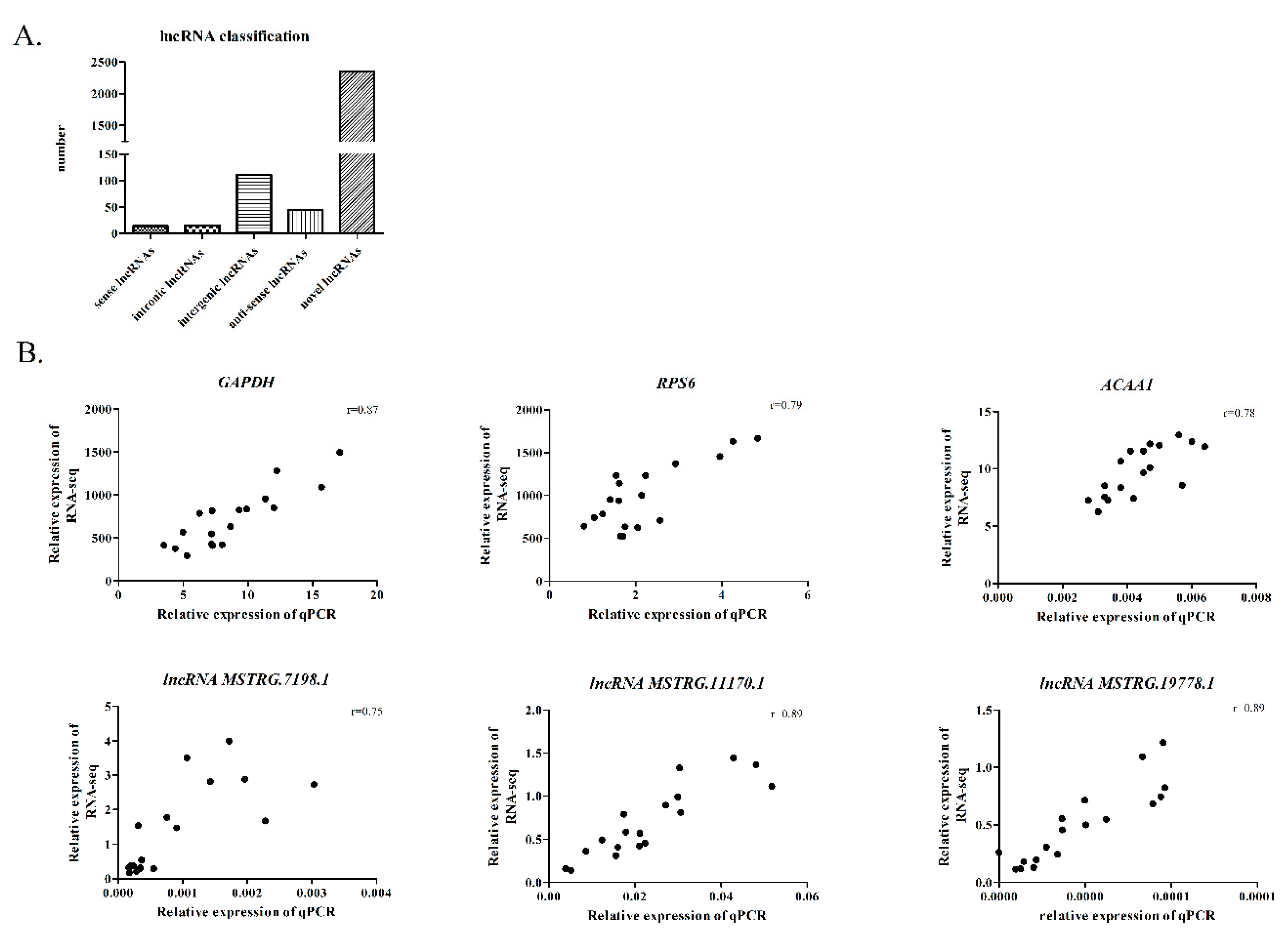
2.5. Identification of the lncRNAs
2.6. Identification of DEGs and DE lncRNAs
2.7. Analysis of the Identified DEGs and DE lncRNAs
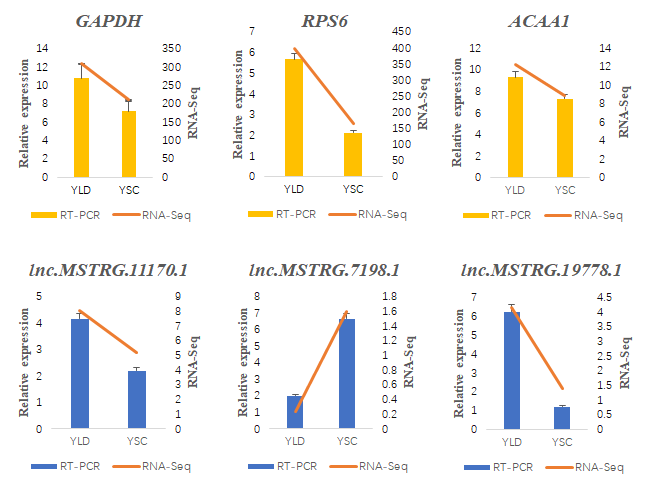
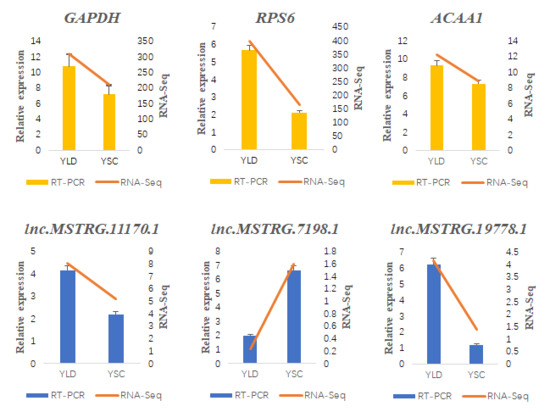
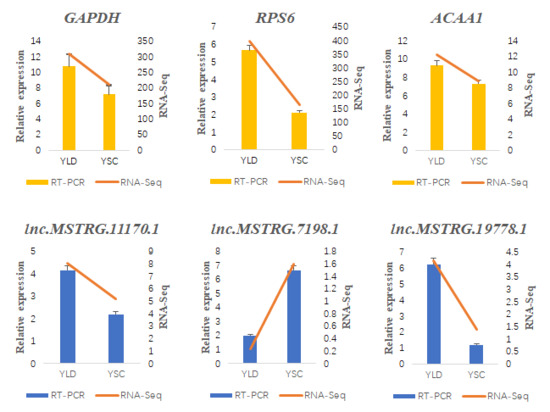
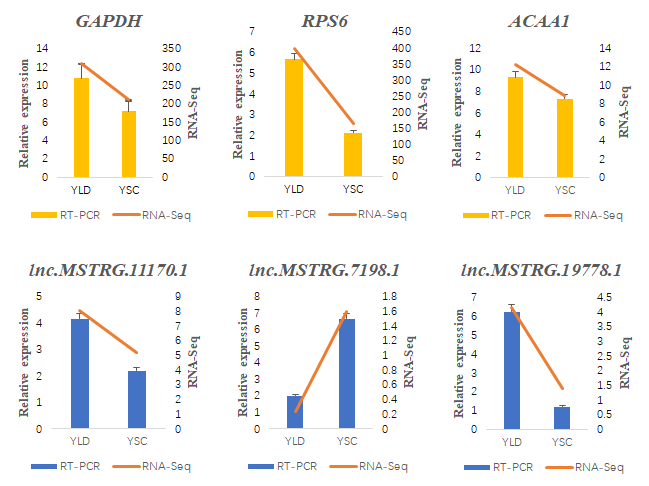
2.8. RT-qPCR Validation of the DE lncRNAs and Their Target Genes
2.9. Sequence Analysis of the HTR Family Genes
3. Results
3.1. Characteristics of All Obtained Ovarian Transcriptomes
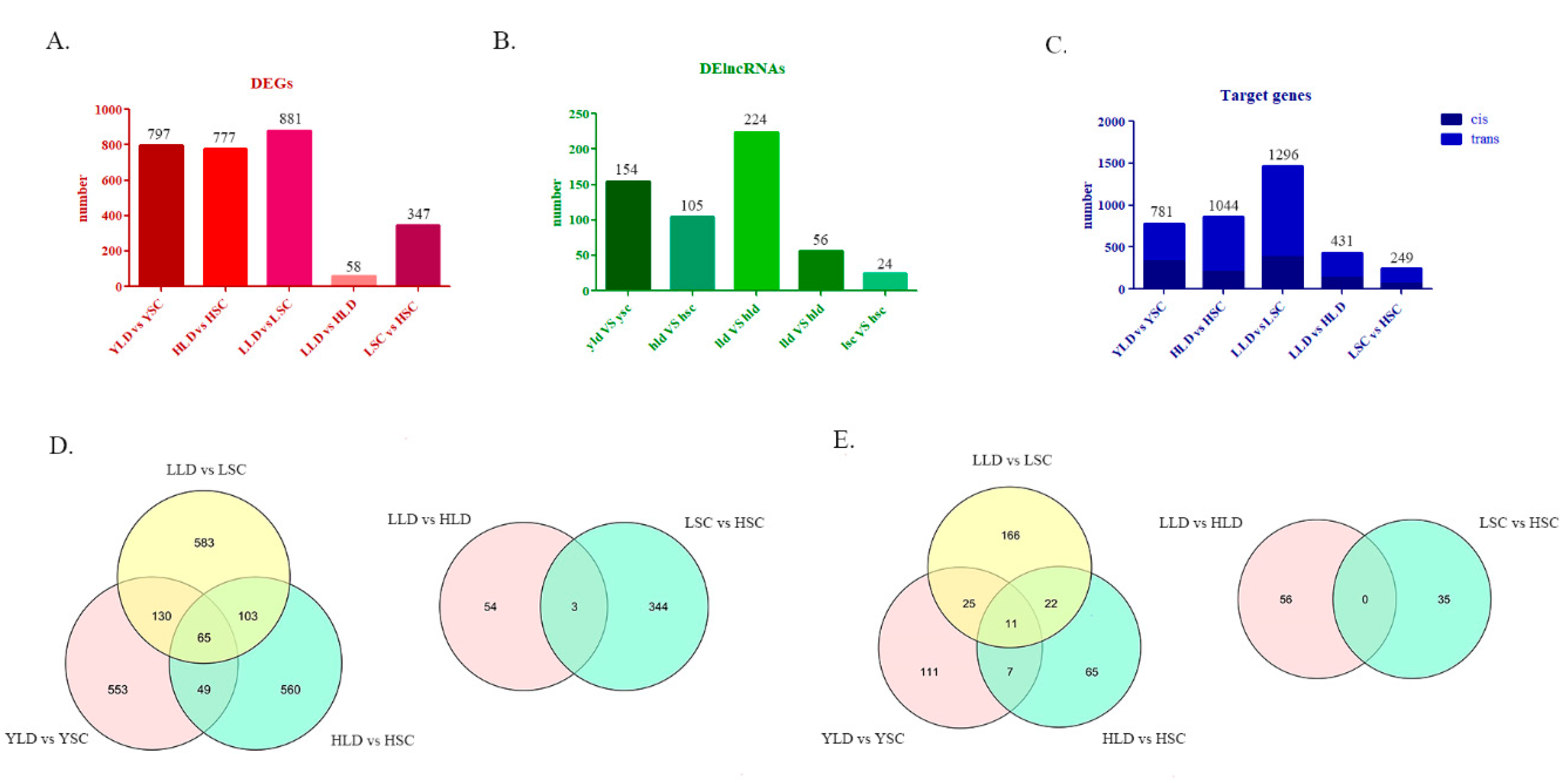
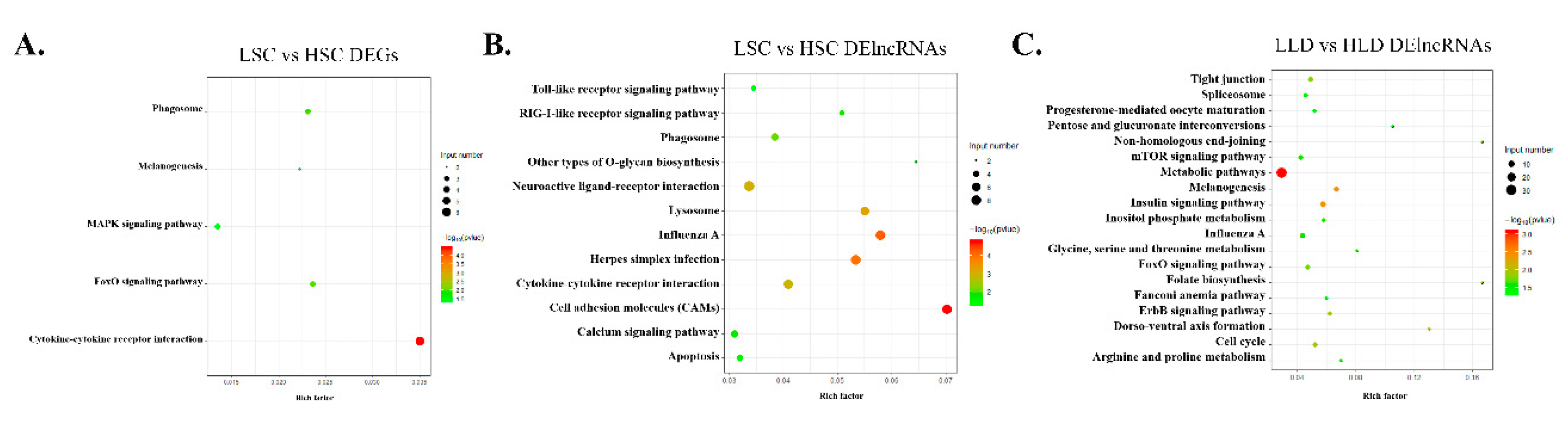
3.2. Identification of DEGs and DE lncRNAs Between Either Different Breeds or Different Egg Production Performance within the Same Breed
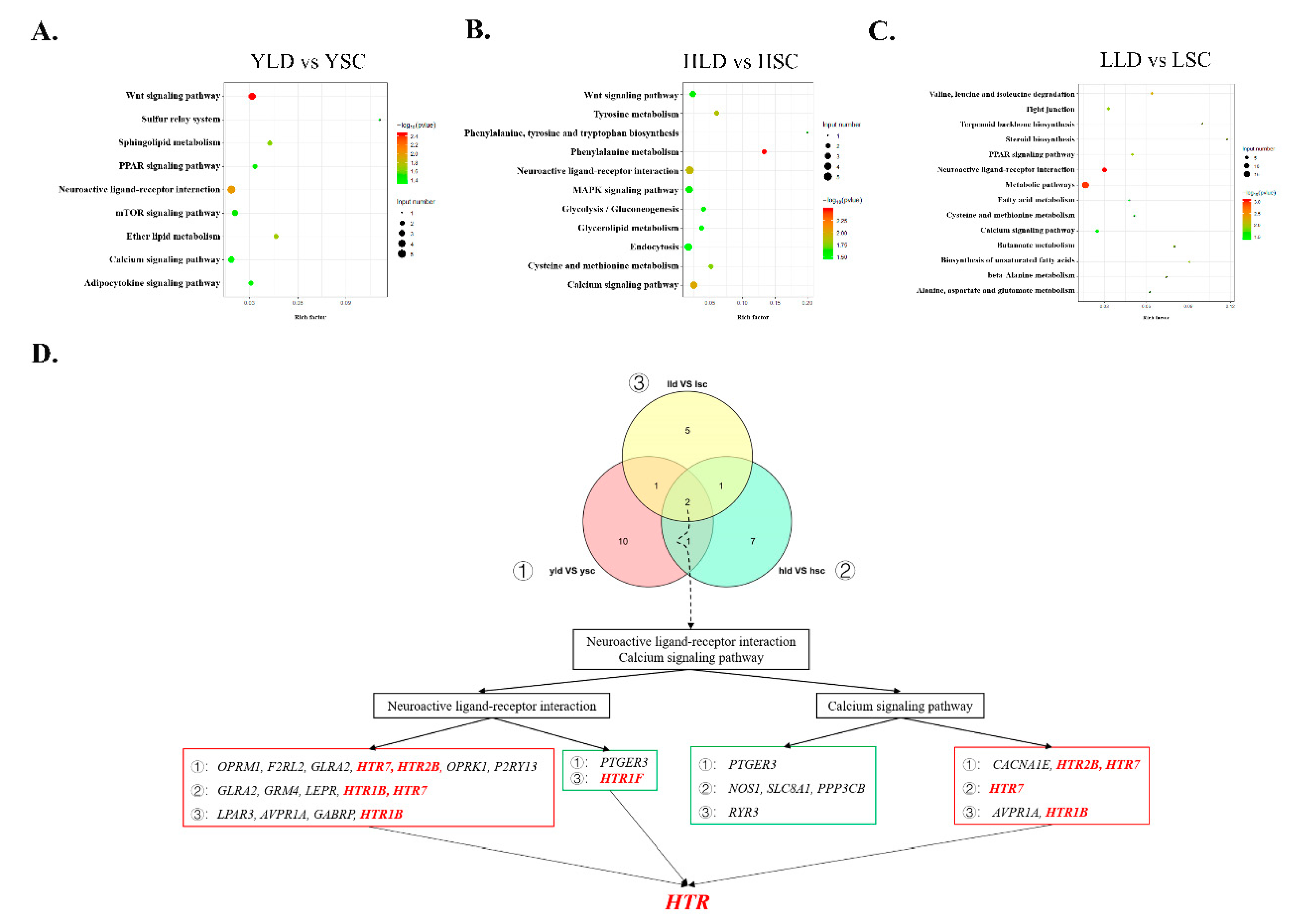
3.3. Functional Analysis of the Identified DEGs Reveals Key Roles for HTR in Control of Egg Production Performance
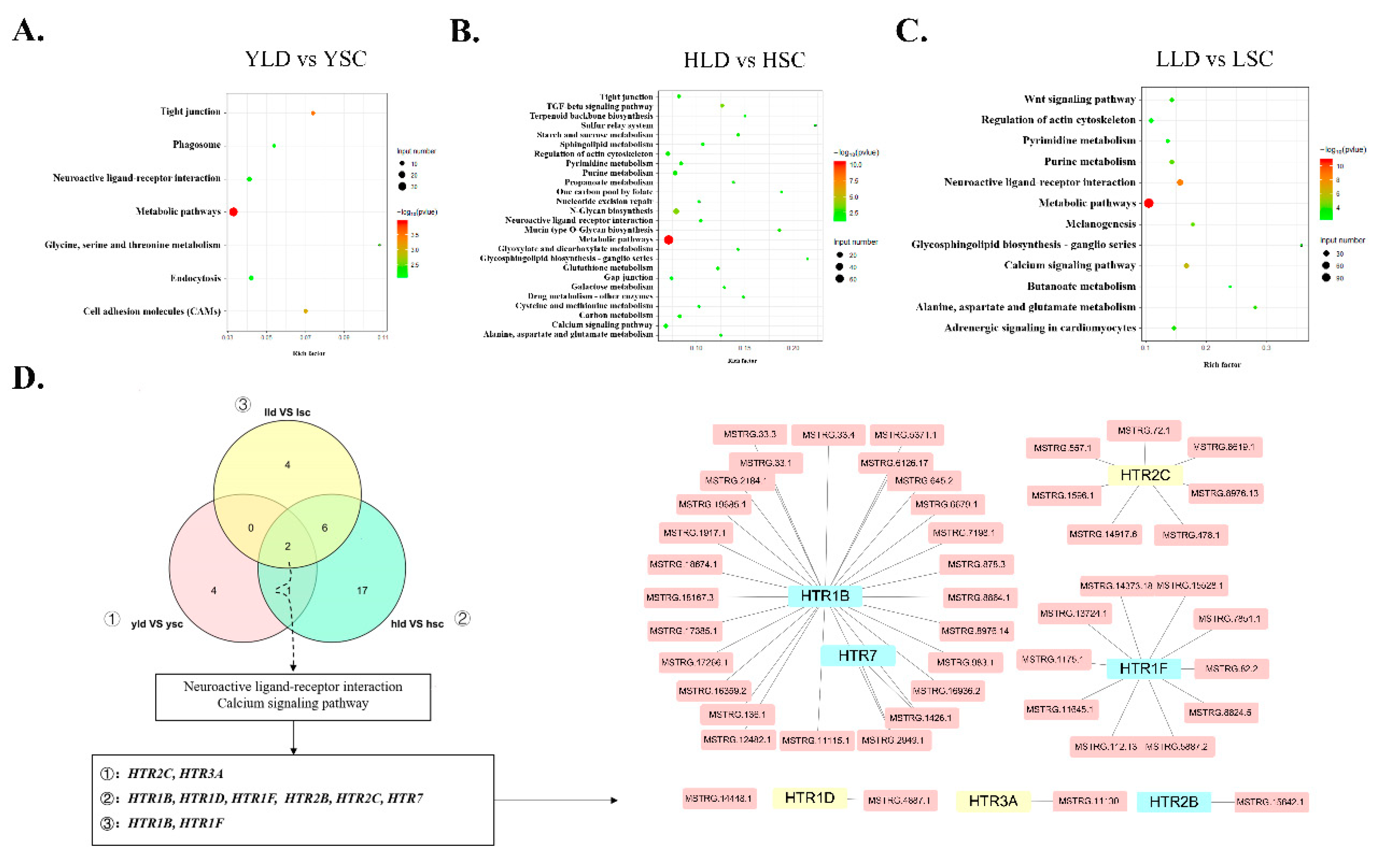
3.4. The DE lncRNAs Target Members of the HTR Family to Regulate Inter-Breed Differences in Egg Production Performance
3.5. HTR1B Regulates Intra-Breed Difference in Egg Production Performance within SWG as Opposed to LG
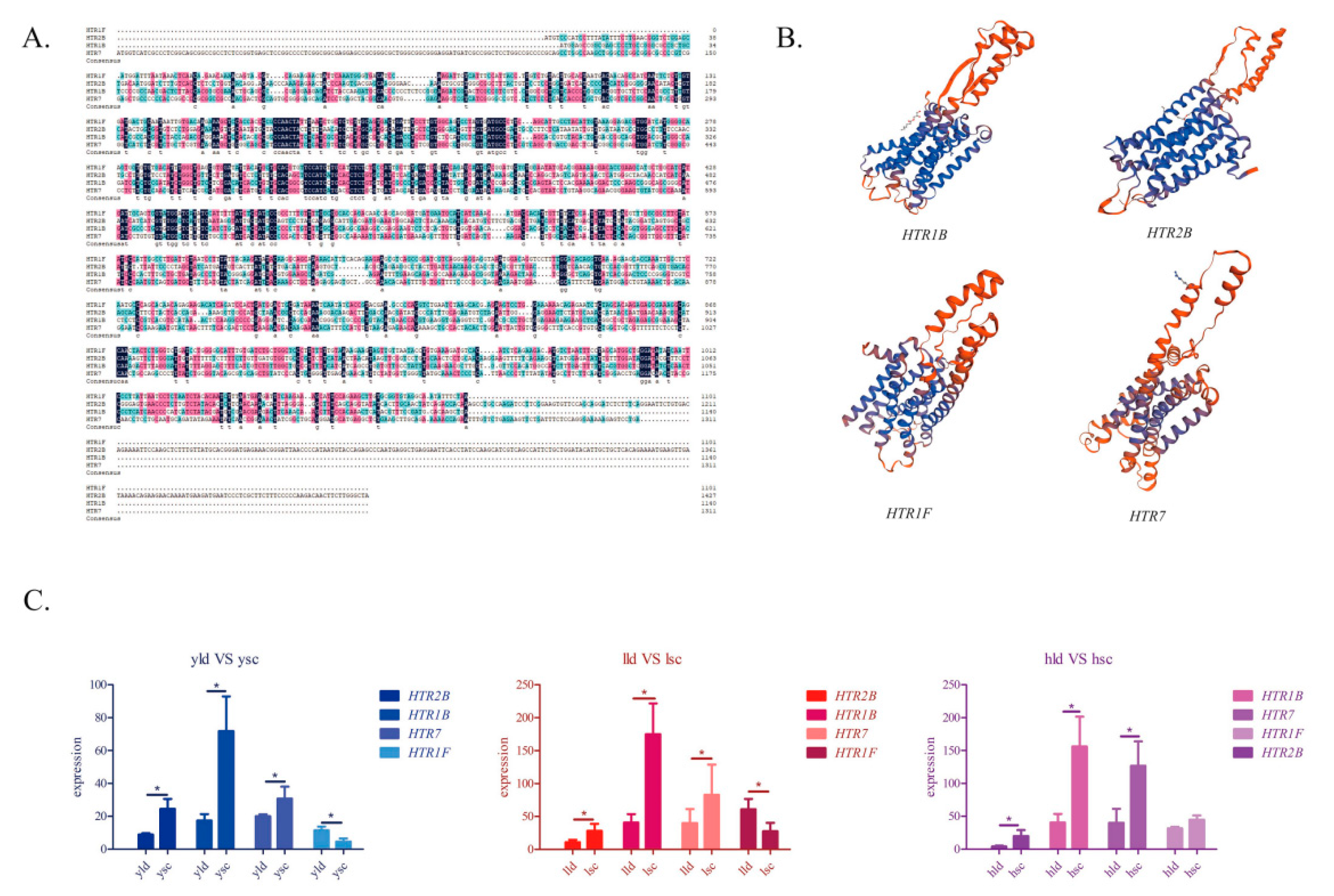
3.6. Structure Prediction and Expression Validation of Several Members of the HTR Gene Family
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Buckland, R.; Guy, G. Goose Production; Food and Agriculture Organization of the United Nations: Rome, Italty, 2002. [Google Scholar]
- Gao, G.L.; Zhao, X.Z.; Li, Q.; Su, J.; Wang, Q.G. Gene expression profiles in the pituitary glands of Sichuan White geese during prelaying and laying periods. Genet. Mol. Res. 2015, 14, 12636–12645. [Google Scholar] [CrossRef]
- Kozák, J.; Bódi, L.; Janan, J. Improvements in the reproductive characteristics of Hungarian Upgraded and Grey Landes geese in Hungary. Worlds Poult. Sci. J. 1997, 53, 197–201. [Google Scholar] [CrossRef]
- Peng, Y.; Chang, L.; Wang, Y.; Wang, R.; Hua, L.; Zhao, Z.; Geng, L.; Liu, Z.; Gong, Y.; Li, J.; et al. Genome-wide differential expression oflong noncoding RNAsandmRNAs in ovarian follicles of two different chicken breeds. Genomics 2019, 111, 1395–1403. [Google Scholar] [CrossRef]
- Tao, Z.; Song, W.; Zhu, C.; Xu, W.; Liu, H.; Zhang, S.; Huifang, L. Comparative transcriptomic analysis of high and low egg-producing duck ovaries. Poult. Sci. 2017, 96, 4378–4388. [Google Scholar] [CrossRef]
- Wu, N.; Gaur, U.; Zhu, Q.; Chen, B.; Xu, Z.; Zhao, X.; Yang, M.; Li, D. Expressed microRNA associated with high rate of egg production in chicken ovarian follicles. Anim. Genet. 2017, 48, 205–216. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, L.; Han, K.; Zhang, X.; Zhang, G.; Dai, G.; Wang, J.; Xie, K. Transcriptome analysis of ovary in relatively greater and lesser egg producing Jinghai Yellow Chicken. Anim. Reprod. 2019, 208, 106114. [Google Scholar] [CrossRef]
- Li, Q.; Hu, S.; Wang, Y.; Deng, Y.; Yang, S.; Hu, J.; Li, L.; Wang, J. mRNA and miRNA Transcriptome Profiling of Granulosa and Theca Layers from Geese Ovarian Follicles Reveals the Crucial Pathways and Interaction Networks for Regulation of Follicle Selection. Front. Genet. 2019, 10, 988. [Google Scholar] [CrossRef]
- Irving-Rodgers, H.F.; Hummitzsch, K.; Murdiyarso, L.S.; Bonner, W.M.; Sado, Y.; Ninomiya, Y.; Couchman, J.R.; Sorokin, L.M.; Rodgers, R.J. Dynamics of extracellular matrix in ovarian follicles and corpora lutea of mice. Cell Tissue Res. 2010, 339, 613–624. [Google Scholar] [CrossRef]
- Onagbesan, O.; Bruggeman, V.; Decuypere, E. Intra-ovarian growth factors regulating ovarian function in avian species: A review. Anim. Reprod. Sci. 2009, 111, 121–140. [Google Scholar] [CrossRef]
- Yerushalmi, G.M.; Salmon-Divon, M.; Yung, Y.; Maman, E.; Kedem, A.; Ophir, L.; Elemento, O.; Coticchio, G.; Canto, M.D.; Renzinu, M.M. Characterization of the human cumulus cell transcriptome during final follicular maturation and ovulation. Mol. Hum. Reprod. 2014, 20, 719–735. [Google Scholar] [CrossRef]
- Xu, X.F.; Li, J.; Cao, Y.X.; Chen, D.W.; Chen, B.L. Differential Expression of Long Noncoding RNAs in Human Cumulus Cells Related to Embryo Developmental Potential: A Microarray Analysis. Reprod. Sci. 2014, 22, 672. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Durruthy-Durruthy, J.; Sebastiano, V.; Wossidlo, M.; Cepeda, D.; Cui, J.; Grow, E.J.; Davila, J.; Mall, M.; Wong, W.H.; Wysocka, J. The primate-specific noncoding RNA HPAT5 regulates pluripotency during human preimplantation development and nuclear reprogramming. Nat. Genet. 2015, 48, 44–52. [Google Scholar] [CrossRef]
- Daehwan, K.; Ben, L.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar]
- Khalil, A.M.; Mitchell, G.; Maite, H.; Manuel, G.; Arjun, R.; Dianali, R.M.; Kelly, T.; Aviva, P.; Bernstein, B.E.; Alexander, V.O. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef]
- Mihaela, P.; Pertea, G.M.; Antonescu, C.M.; Tsung-Cheng, C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650. [Google Scholar] [CrossRef]
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef]
- Liang, S.; Haitao, L.; Dechao, B.; Guoguang, Z.; Kuntao, Y.; Changhai, Z.; Yuanning, L.; Runsheng, C.; Yi, Z. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar]
- Frazee, A.C.; Geo, P.; Jaffe, A.E.; Ben, L.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 2015, 33, 243. [Google Scholar] [CrossRef]
- Qi, Z.; Wang, X.J. GOEAST: A web-based software toolkit for Gene Ontology enrichment analysis. Nucleic Acids Res. 2008, 36, W358–W363. [Google Scholar]
- Ai, C.; Kong, L.; Bioinformatics, C.F. CGPS:A machine learning-based approach integrating multiple gene set analysis tools for better prioritization of biologically relevant pathways. Genetics 2018, 45, 489–504. [Google Scholar]
- Xiang, W.; Shang, Y.; Wang, Q.; Xu, Y.; Zhu, P.; Huang, K.; Xu, W. Identification of a chicken (Gallus gallus) endogenous reference gene (Actb) and its application in meat adulteration. Food Chem. 2017, 234, 472–478. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A.M. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Georjon, C.; Deleage, G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef]
- Schwede, T.; Kopp, J.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef]
- Xu, S.; Dingyue, W.; Dongsheng, Z.; Yan, L.; Lianqiang, C.; Zhengfeng, F.; De, W.; Wei, S. Reproductive Hormone and Transcriptomic Responses of Pituitary Tissue in Anestrus Gilts Induced by Nutrient Restriction. PLoS ONE 2015, 10, e0143219. [Google Scholar] [CrossRef]
- Feng, S.; Xiaoli, G.; Yanchao, W.; Yuding, W.; Guiling, C.; Yunliang, J. Genome-Wide Analysis on the Landscape of Transcriptomes and Their Relationship With DNA Methylomes in the Hypothalamus Reveals Genes Related to Sexual Precocity in Jining Gray Goats. Front. Endocrinol. 2018, 9, 501. [Google Scholar]
- Chen, H.; Feng, W.; Chen, K.; Qiu, X.; Xu, H.; Mao, G.; Zhao, T.; Ding, Y.; Wu, X. Transcriptomic analysis reveals potential mechanisms of toxicity in a combined exposure to dibutyl phthalate and diisobutyl phthalate in zebrafish (Danio rerio) ovary. Aquat. Toxicol. 2019, 216, 105290. [Google Scholar] [CrossRef]
- Wu, C.; Lv, C.; Wan, Y.; Li, X.; Zhang, J.; Li, J.; Wang, Y. Arginine vasotocin (AVT)/mesotocin (MT) receptors in chickens: Evidence for the possible involvement of AVT-AVPR1 signaling in the regulation of oviposition and pituitary prolactin expression. Gen. Comp. Endocrinol. 2019, 15, 91–104. [Google Scholar] [CrossRef]
- Agirregoitia, E.; Peralta, L.; Mendoza, R.; Expósito, A.; Ereo, E.D.; Matorras, R.; Agirregoitia, N. Expression and localization of opioid receptors during the maturation of human oocytes. Reprod. Biomed. Online 2012, 24, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Teodorov, E.; Ferrari, M.F.R.; Fior-Chadi, D.R.; Camarini, R.; Felício, L.F. Behavioral meaningful opioidergic stimulation activates kappa receptor gene expression. Braz. J. Med. Biol. Res. 2012, 45, 982–987. [Google Scholar] [CrossRef] [PubMed]
- Liszewska, E.; Reinaud, P.; Dubois, O.; Charpigny, G. Lysophosphatidic acid receptors in ovine uterus during estrous cycle and early pregnancy and their regulation by progesterone. Domest. Anim. Endocrinol. 2012, 42, 31–42. [Google Scholar] [CrossRef]
- Diao, H.; Li, R.; El Zowalaty, A.E.; Xiao, S.; Zhao, F.; Dudley, E.A.; Ye, X. Deletion of Lysophosphatidic Acid Receptor 3 (Lpar3) Disrupts Fine Local Balance of Progesterone and Estrogen Signaling in Mouse Uterus During Implantation. Biol. Reprod. 2015, 93, 123. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Diao, H.; Chun, J. 11-deoxy prostaglandin F2α, a thromboxane A2 receptor agonist, partially alleviates embryo crowding in Lpar3 females. Fertil. Steril. 2012, 97, 757–763. [Google Scholar] [CrossRef][Green Version]
- Gokul, C.; Parida, S.; Singh, T.U.; Panigrahi, M.; Suhas, K.S.; Soni, K.; Kesavan, M.; Srivastava, V.; Kumar, T.S.; Mahobiya, A. Lysophosphatidic acid enhances PGE2 to PGF2α ratio and nitric oxide level in nonpregnant buffalo uterus. Theriogenology 2018, 120, 47–55. [Google Scholar] [CrossRef]
- Zhao, X.; Shao, T.; Wang, Y.Q.; Lu, X.L.; Luo, J.B.; Zhou, W.D. The phytoestrogen daidzein may affect reproductive performance of Zhedong White geese by regulating gene mRNA levels in the HPG axis. Br. Poult. Sci. 2013, 54, 252–258. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Li, L.; Han, C.; Xu, H. Transcriptome analysis revealed the possible regulatory pathways initiating female geese broodiness within the hypothalamic-pituitary-gonadal axis. PLoS ONE 2018, 13, e0191213. [Google Scholar] [CrossRef]
- Woods, D.C.; Schorey, J.S.; Johnson, A.L. Toll-like receptor signaling in hen ovarian granulosa cells is dependent on stage of follicle maturation. Reproduction 2009, 137, 987–996. [Google Scholar] [CrossRef]
- Berger, M.; Gray, J.A.; Roth, B.L. The Expanded Biology of Serotonin—Annual Review of Medicine. Ann. Rev. Med. 2009, 60, 355. [Google Scholar] [CrossRef]
- Avital-Cohen, N.; Heiblum, R.; Argov-Argaman, N.; Rosenstrauch, A.; Chaiseha, Y.; Mobarkey, N.; Rozenboim, I. Age-related changes in gonadal and serotonergic axes of broiler breeder roosters. Domest. Anim. Endocrinol. 2015, 44, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, H.; Kawashima, M.; Kamiyoshi, M.; Tanaka, K. Effect of serotonin and β-endorphin on the release of luteinizing hormone in the hen (Gallus domesticus). Gen. Comp. Endocrinol. 1986, 63, 24–30. [Google Scholar] [CrossRef]
- El-Merahbi, R.; Löffler, M.; Mayer, A.; Sumara, G. The roles of peripheral serotonin in metabolic homeostasis. Febs Lett. 2015, 589, 1728–1734. [Google Scholar] [CrossRef] [PubMed]
- Hitoshi, W.; Daisuke, A.; Hideki, O.; Kan, S.; Masato, M.; Kazuki, S.; Yu, T.; Takashi, K.; Ikuro, T.; Tetsuya, H. Peripheral Serotonin Enhances Lipid Metabolism by Accelerating Bile Acid Turnover. Endocrinology 2010, 151, 4776–4786. [Google Scholar]
- Sumara, G.; Sumara, O.; Kim, J.K.; Karsenty, G. Gut-Derived Serotonin Is a Multifunctional Determinant to Fasting Adaptation. Cell Metab. 2012, 16, 588–600. [Google Scholar] [CrossRef]
- Söhle, J.; Machuy, N.; Smailbegovic, E.; Holtzmann, U.; Grönniger, E.; Wenck, H.; Stäb, F.; Winnefeld, M. Identification of New Genes Involved in Human Adipogenesis and Fat Storage. PLoS ONE 2012, 7, e31393. [Google Scholar] [CrossRef]
- Burhans, M.S.; Flowers, M.T.; Harrington, K.R.; Bond, L.M.; Guo, C.-A.; Anderson, R.M.; Ntambi, J.M. Hepatic oleate regulates adipose tissue lipogenesis and fatty acid oxidation. J. Lipid Res. 2015, 56, 304–318. [Google Scholar] [CrossRef]
- Sinner, D.I.; Kim, G.J.; Henderson, G.C.; Igal, R.A. StearoylCoA Desaturase-5: A Novel Regulator of Neuronal Cell Proliferation and Differentiation. PLoS ONE 2012, 7, e39787. [Google Scholar] [CrossRef]
- Han, C.; Wang, J.; Xu, H.; Li, L.; Ye, J.; Li, J.; Zhuo, W. Effect of Overfeeding on Plasma Parameters and mRNA Expression of Genes Associated with Hepatic Lipogenesis in Geese. Asian Aust. J. Anim. Sci. 2008, 21, 590–595. [Google Scholar] [CrossRef]
- Yunlei, L.I.; Sun, Y.; Chen, J. Research Progresses on Effects of Fat Deposition on Reproduction Performance of Poultry. China Poult. 2016, 38, 36–41. [Google Scholar]
- Máčajová, M.; Lamošová, D.; Zeman, M. Role of leptin in Japanese quail development. Acta Vet. Brno 2002, 71, 473–479. [Google Scholar] [CrossRef][Green Version]
- Sam, A.H.; Dhillo, W.S. Endocrine links between fat and reproduction. Obstet. Gynaecol. 2010, 12, 231–236. [Google Scholar] [CrossRef]
- Derkach, K.V.; Sukhov, I.B.; Bondareva, V.M.; Shpakov, A.O. Effect of Metformin on Metabolic Parameters and Hypothalamic Signaling Systems in Rats with Obesity Induced by a High-Carbohydrate and High-Fat Diet. Adv. Gerontol. 2018, 8, 228–234. [Google Scholar] [CrossRef]
- Nicole, B.; Nomi, A.; Diana, M.; Peter, V.; Gérard, W.; Vincent, M.; Matthias, E.; Murielle, B.; Philippe, R. Association between Variants of the Leptin Receptor Gene (LEPR) and Overweight: A Systematic Review and an Analysis of the CoLaus Study. PLoS ONE 2011, 6, e26157. [Google Scholar]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.L.; Bartel, D.P. Conserved Function of lincRNAs in Vertebrate Embryonic Development Despite Rapid Sequence Evolution. Cell 2011, 147, 1537–1550. [Google Scholar] [CrossRef]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2017, 19, 143–157. [Google Scholar] [CrossRef]
- Bai, L.; Chu, G.; Mai, Y.; Zheng, J.; Wang, W.; Zhang, Q.; Yang, G. Identification and expression analyses of BAMBI mediated by FSH in swine luteinizing granulosa cells. Theriogenology 2012, 82, 1094–1101. [Google Scholar] [CrossRef]
- Bai, L.; Chu, G.; Wang, W.; Xiang, A.; Yang, G. BAMBI promotes porcine granulosa cell steroidogenesis involving TGF-β signaling. Theriogenology 2017, 100, 24–31. [Google Scholar] [CrossRef]
- Ding, T.; Zhou, Y.; Long, R.; Chen, C.; Zhao, J.; Cui, P.; Guo, M.; Liang, G.; Xu, L. DUSP8 phosphatase: Structure, functions, expression regulation and the role in human diseases. Cell Biosci. 2019, 9, 70. [Google Scholar] [CrossRef]
- Wang, F.; Chang, J.T.-H.; Kao, C.J.; Huang, R.S. High expression of miR-532-5p, a tumor suppressor, leads to better prognosis in ovarian cancer both in vivo and in vitro. Mol. Cancer Ther. 2016, 15, 1535–7163. [Google Scholar] [CrossRef]
- Bai, H.; Li, H.; Li, W.; Gui, T.; Shen, K. The PI3K/AKT/mTOR pathway is a potential predictor of distinct invasive and migratory capacities in human ovarian cancer cell lines. Oncotarget 2015, 6, 25520–25532. [Google Scholar] [CrossRef] [PubMed]
- Melhem, A.; Yamada, S.D.; Fleming, G.F.; Delgado, B.; Brickley, D.R.; Wu, W.; Kocherginsky, M.; Conzen, S.D. Administration of Glucocorticoids to Ovarian Cancer Patients Is Associated with Expression of the Anti-apoptotic Genes SGK1 and MKP1/DUSP1 in Ovarian Tissues. Clin. Cancer Res. 2009, 15, 3196–3204. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.K.; Abdollah, N.A.; Shafie, N.H.; Yusof, N.M.; Razak, S.R.A. Dual-specificity phosphatase 6(DUSP6): A review of its molecular characteristics and clinical relevance in cancer. Cancer Biol. Med. 2018, 15, 22–36. [Google Scholar]
- Johnson, P.A.; Giles, J.R. The hen as a model of ovarian cancer. Nat. Rev. Cancer 2013, 13, 432–436. [Google Scholar] [CrossRef]
- Mccorvy, J.D.; Roth, B.L. Structure and function of serotonin G protein-coupled receptors. Pharmacol. Ther. 2015, 150, 129–142. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, Y.; Ma, J.; Wu, H.; Wacker, D.; Katritch, V.; Han, G.W.; Liu, W.; Huang, X.P.; Vardy, E. Structural Basis for Molecular Recognition at Serotonin Receptors. Science 2013, 340, 610–614. [Google Scholar] [CrossRef]
- Wacker, D.; Wang, C.; Katritch, V.; Han, G.W.; Huang, X.P.; Vardy, E.; Mccorvy, J.D.; Jiang, Y.; Chu, M.; Siu, F.Y. Structural Features for Functional Selectivity at Serotonin Receptors. Science 2013, 340, 615–619. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Name | Control Group | Experimental Group |
|---|---|---|
| YLD vs. YSC | LG at 145 days | SWG at 145 days |
| HLD vs. HSC | LG with high egg production performance at 730 days | SWG with high egg production performance at 730 days |
| LLD vs. LSC | LG with low egg production performance at 730 days | SWG with low egg production performance at 730 days |
| LLD vs. HLD | LG with low egg production performance at 730 days | LG with high egg production performance at 730 days |
| LSC vs. HSC | SWG with low egg production performance at 730 days | SWG with high egg production performance at 730 days |
| Primer Name | Sequence (5′-3′) | Product Length (bp) |
|---|---|---|
| ACAA1-F | CGCTTTGGTCGCAAGAGTT | 187 |
| ACAA1-R | ATTGGCACTTCTGAGGGACAT | |
| RPS6-F | TTGTCCGAATCAGTGGTGGC | 121 |
| RPS6-R | GTTCTCCTGGGGCGGTAGC | |
| GAPDH-F | CATGTTCGTGATGGGTGTG | 239 |
| GAPDH-R | CTGGGATAATGTTCTGGGC | |
| lncRNA.MSTRG.7198.1-F | TCCTTACTCCTGCTTCTACCA | 114 |
| lncRNA.MSTRG.7198.1-R | CCTGGCAACTTCTTGTCTGT | |
| lncRNA.MSTRG.19978.1-F | CCAGACCACAGAGCCAAACA | 100 |
| lncRNA.MSTRG.19978.1-R | CCCCCAGACATCAGCAAGAG | |
| lncRNA.MSTRG.11170.1-F | AGTGAGAGGAGTGAGGAACAG | 129 |
| lncRNA.MSTRG.11170.1-R | GGACAGCCTGCTTCACC | |
| HTR7-F | GCAGCCCTCCAACTATCTC | 225 |
| HTR7-R | AGAGGTCTTGTTATTCCCAGG | |
| HTR1F-F | CTGTAGCCCTGCCTTCTCCC | 99 |
| HTR1F-R | GTGGCTCGCTATGAACTGGTAAC | |
| HTR1B-F | TTCCCCACTTTGCTGCTGATA | 108 |
| HTR1B-R | AGCCCGAGTTAGTCTTTTACCC | |
| HTR2B-F | GAACCTCACTCTAAAGGGGAC | 187 |
| HTR2B-R | ATGGTAAACTGGTCATCTGCTA | |
| β-actin-F | CAACGAGCGGTTCAGGTGT | 99 |
| β-actin-R | TGGAGTTGAAGGTGGTCTCGT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, Q.; Hu, S.; Wang, G.; Hu, J.; Zhang, J.; Li, L.; Hu, B.; He, H.; Liu, H.; Xia, L.; et al. Comparative Transcriptome Analysis Suggests Key Roles for 5-Hydroxytryptamlne Receptors in Control of Goose Egg Production. Genes 2020, 11, 455. https://doi.org/10.3390/genes11040455
Ouyang Q, Hu S, Wang G, Hu J, Zhang J, Li L, Hu B, He H, Liu H, Xia L, et al. Comparative Transcriptome Analysis Suggests Key Roles for 5-Hydroxytryptamlne Receptors in Control of Goose Egg Production. Genes. 2020; 11(4):455. https://doi.org/10.3390/genes11040455
Chicago/Turabian StyleOuyang, Qingyuan, Shenqiang Hu, Guosong Wang, Jiwei Hu, Jiaman Zhang, Liang Li, Bo Hu, Hua He, Hehe Liu, Lu Xia, and et al. 2020. "Comparative Transcriptome Analysis Suggests Key Roles for 5-Hydroxytryptamlne Receptors in Control of Goose Egg Production" Genes 11, no. 4: 455. https://doi.org/10.3390/genes11040455
APA StyleOuyang, Q., Hu, S., Wang, G., Hu, J., Zhang, J., Li, L., Hu, B., He, H., Liu, H., Xia, L., & Wang, J. (2020). Comparative Transcriptome Analysis Suggests Key Roles for 5-Hydroxytryptamlne Receptors in Control of Goose Egg Production. Genes, 11(4), 455. https://doi.org/10.3390/genes11040455