Transforming Ocean Conservation: Applying the Genetic Rescue Toolkit


Abstract
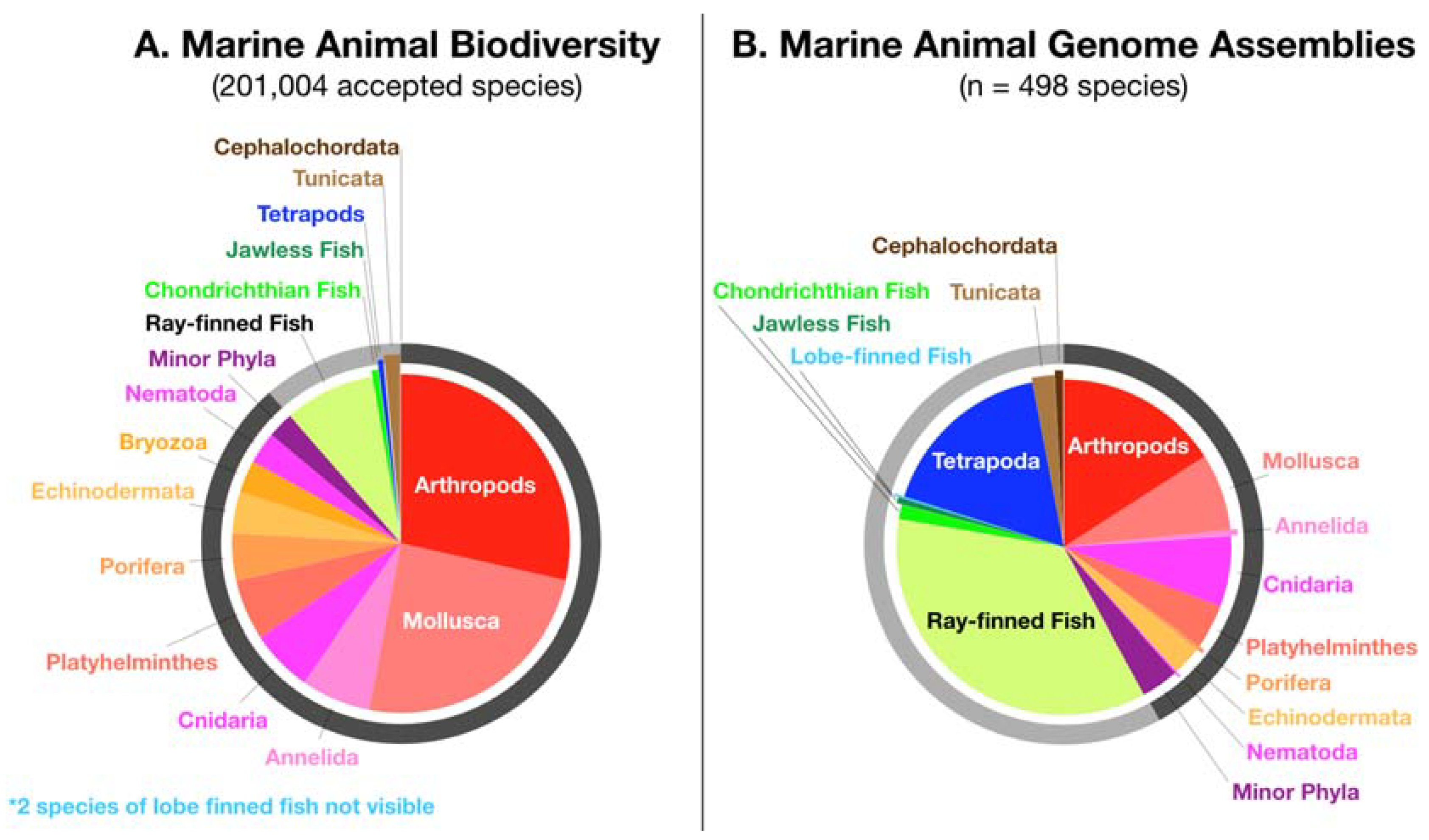
1. Introduction
2. The Ocean Genomics Rescue Toolkit Continuum
2.1. Overexploitation
2.1.1. Insight for Combating Illegal Trade
2.1.2. Insight for Sustainable Fishing
2.1.3. Insight and Intervention for Alternatives to Wild Harvest
2.2. Pollution
Insight and Intervention for Plastics Bioremediation
2.3. Invasives
2.3.1. Insight for Monitoring and Early Detection
2.3.2. Intervention for Genetic Biocontrol
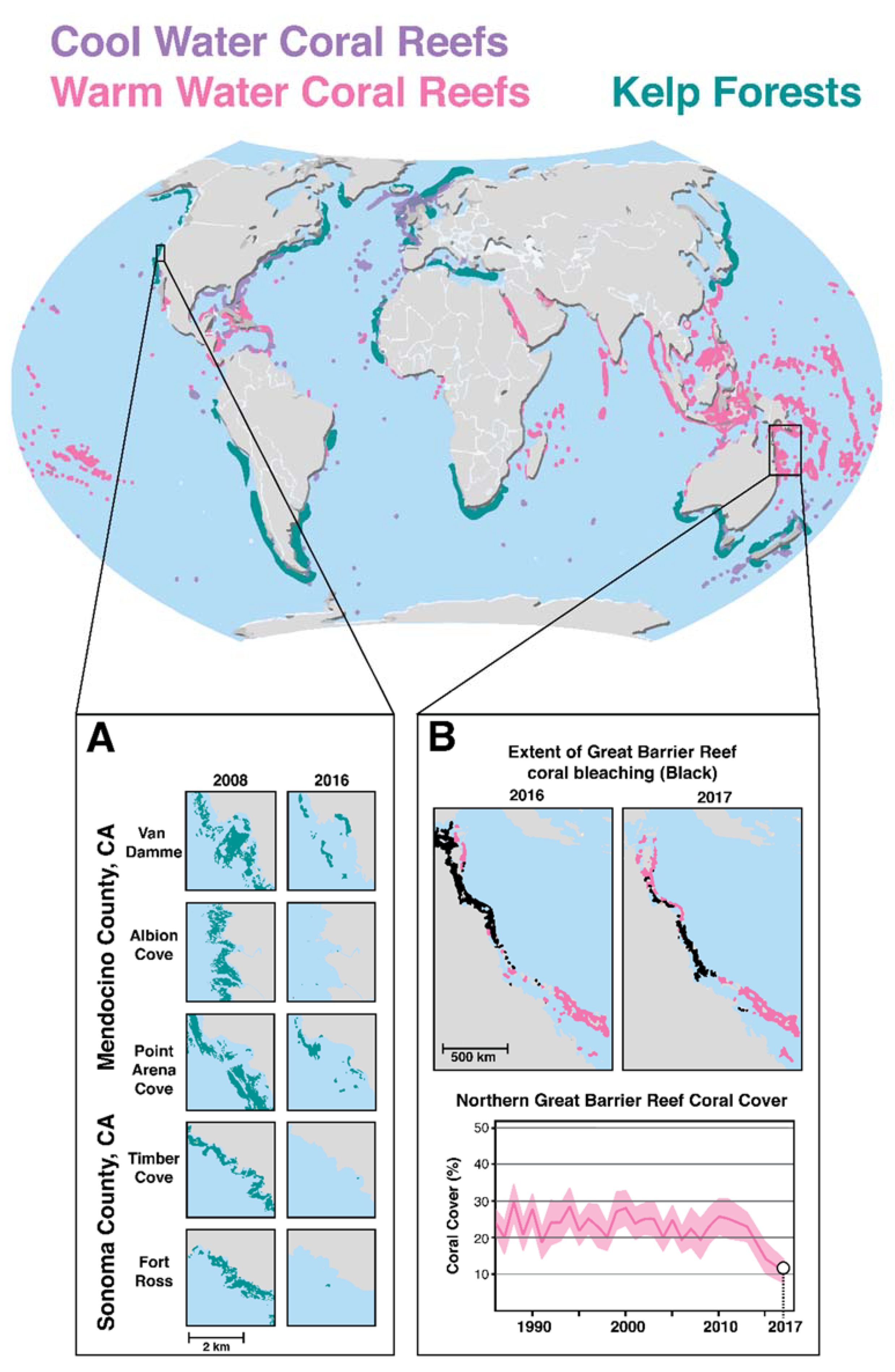
2.4. Climate Change
Insight and Intervention for Adaptation
3. The Coral Toolkit: A Case Study for Advancing Genomics in Conservation
3.1. Coral Cryopreservation
3.2. Inducible Spawning
3.3. Coral Stem Cells
4. The Applicability and Readiness of Genomic and Biotechnological Solutions for Conservation
5. Discussion
- Weighing Risks and Benefits: Genetic rescue program developers should conduct a thorough review of both the intended outcomes and the potential benefits of applying genetic insight or genomic intervention, as well as the risks and potential consequences. Questions to consider include: Have other more established interventions failed? Do other interventions, such as the application of antibiotics or pesticides, have potentially worse environmental consequences? Is this intervention the most efficacious, lasting, and least risky solution to an environmental problem, not merely an equivalent or novel approach?
- Transparency: Genetic rescue program developers in collaboration with conservation managers should proactively identify and inform key stakeholders in the early stages of technology development. This includes partners from the private sector (particularly biotech firms), social science experts, public sector partners, international and research organizations, religious and ethical organizations, NGOs, and local communities. This should involve objective discussions with the public about the risks and potential impacts of proposed environmental solutions and also subjective discussions about values.
- Procedures: Genetic rescue program developers should conduct systematic and data-driven reviews of recommended best practices. These should include surveys of recent intentional environmental release of organisms (including biocontrol, re-introductions, and translocations) into natural environments and studies of the resulting long-term effects.
- Tests: Genetic rescue programs should first field test genomic technologies in contained environments to minimize unintended environmental consequences. These should be simple enough to be cost-effective, yet complex enough to sufficiently mimic natural ecosystems to yield useful data on the efficacy of developing technologies. Suitable test environments will be especially important for marine environments.
- Predictions: Genetic rescue programs should employ computational models, particularly as they grow more sophisticated with time, to predict the long-term effects on ecosystems and highlight potential failures of specific applications of genetic technologies.
- Measures: Genetic rescue scientists should employ standard metrics for measuring safety and efficacy at appropriate environmental scales agreed upon in collaboration with regulators, so that direct comparisons of data sets can be made, and so universal analytic tools can be developed and useful to final-stage decision making.
- Protocols: Genetic rescue program developers should understand the regulatory approval processes and jurisdictions that currently guide the transition of new technologies from the lab to field trials, and should be prepared to help guide new policies when needed.
- Public-Private Partnerships: Genetic rescue programs should collaborate with philanthropists, NGOs, agencies, and community groups in the implementation of these technologies to evaluate risk, to address controversies, and to gain critical stakeholder support. These groups can convene conversations, commission studies, identify priorities, and connect with civil society to identify safeguards.
- Remediation: Genetic rescue program developers should prepare for the possibility of failure. In the event of unintended consequences, will it be possible to regain control of the organism? Possibilities for this include the creation of self-limiting gene drives.
- Learnings: Genetic rescue program scientists and conservation managers should continuously monitor introduced organisms and the relevant environment once the technology has been deployed. Since these technologies remain new, these findings should be shared so that researchers learn of the successes as well as failures. Ultimately, transparency will allow successful intervention techniques to be more rapidly adopted globally.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halpern, B.S.; Selkoe, K.A.; Micheli, F.; Kappel, C.V. Evaluating and ranking the vulnerability of global marine ecosystems to anthropogenic threats. Conserv. Biol. 2007, 21, 1301–1315. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.A.; Goold, H.D. Harnessing synthetic biology for kelp forest conservation. J. Phycol. 2019, 55, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Filbee-Dexter, K.; Smajdor, A. Ethics of Assisted Evolution in Marine Conservation. Front. Mar. Sci. 2019, 6, 20. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.; Crowder, L.B.; Halpern, B.S.; Micheli, F. Uncertainty analysis and robust areas of high and low modeled human impact on the global oceans. Conserv. Biol. 2018, 32, 1368–1379. [Google Scholar] [CrossRef]
- Chiba, S.; Saito, H.; Fletcher, R.; Yogi, T.; Kayo, M.; Miyagi, S.; Ogido, M.; Fujikura, K. Human footprint in the abyss: 30 year records of deep-sea plastic debris. Mar. Policy 2018, 96, 204–212. [Google Scholar] [CrossRef]
- Weatherdon L., V.; Magnan, A.K.; Rogers, A.D.; Sumaila, R.R.; Cheung, W.W.L. Observed and Projected Impacts of Climate Change on Marine Fisheries, Aquaculture, Coastal Tourism, and Human Health: An Update. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef]
- Béné, C.; Barange, M.; Subasinghe, R.; Pinstrep-Andersen, P.; Merino, G.; Hemre, G.; Williams, M. Feeding 9 billion by 2050–Putting fish back on the menu. Food Secur. 2015, 7, 261–274. [Google Scholar] [CrossRef]
- Cermak, N.; Becker, J.W.; Knudsen, S.M.; Chisholm, S.W.; Manalis, S.R.; Polz, M.F. Direct single-cell biomass estimates for marine bacteria via Archimedes’ principle. ISME J. 2017, 11, 825–828. [Google Scholar] [CrossRef]
- Broad, W.J. Mystery by the Trillions. New York Time. Available online: https://www.nytimes.com/2015/06/30/science/bristlemouth-ocean-deep-sea-cyclothone.html (accessed on 12 October 2019).
- Atkinson, A.; Siegel, V.; Pakhomov, E.A.; Jessop, M.J.; Loeb, V. A re-appraisal of the total biomass and annual production of Antarctic krill. Deep-Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 727–740. [Google Scholar] [CrossRef]
- Humes, A.G. How many copepods? Hydrobiologia 1994, 292, 1–7. [Google Scholar] [CrossRef]
- U.S. EIA. Annual Energy Outlook 2019 with Projections to 2050; U.S. Energy Information Administration. 2019. Available online: https://www.eia.gov/outlooks/aeo/ (accessed on 5 August 2019).
- Ban, N.C.; Davies, T.E.; Aguilera, S.E.; Brooks, C.M.; Cox, M.; Epstein, G.; Evans, L.S.; Maxwell, S.M. ’ Nenadovic, M. Social and ecological effectiveness of large marine protected areas. Glob. Environ. Chang. 2017, 43, 82–91. [Google Scholar] [CrossRef]
- Parsons, E.C.M.; Favaro, B.; Aguirre, A.A.; Bauer, A.L.; Blight, L.K.; Cigliano, J.A.; Coleman, M.A.; Côté, I.M.; Draheim, M.; Fletcher, S. Seventy-one important questions for the conservation of marine biodiversity. Conserv. Biol. 2014, 28, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, G.S.; McCarthy, J.J. Personalized medicine: Revolutionizing drug discovery and patient care. Trends Biotechnol. 2001, 19, 491–496. [Google Scholar] [CrossRef]
- Roukos, D.H. Next-generation sequencing and epigenome technologies: Potential medical applications. Expert Rev. Med. Devices 2010, 7, 723–726. [Google Scholar] [CrossRef]
- Hood, L.; Balling, R.; Auffray, C. Revolutionizing medicine in the 21st century through systems approaches. Biotechnol. J. 2012, 7, 992–1001. [Google Scholar] [CrossRef]
- Cardon, L.R.; Harris, T. Precision medicine, genomics and drug discovery. Hum. Mol. Genet. 2016, 25, R166–R172. [Google Scholar] [CrossRef]
- Henry, R.J. Genomics Strategies for Germplasm Characterization and the Development of Climate Resilient Crops, Crop Breeding; Apple Academic Press: Cambridge, MA, USA, 2016; pp. 25–34. [Google Scholar]
- Armenta, T.; Askew, J.; Baumgartner, B.; Bogomolni, A.; Demir-Hilton, E.; Fraser, D.; Getsy, J.; Holmes, N.; Kofler, N.; Maciej, M.; et al. Ocean Genomics Horizon Scan; Revive & Restore: Sausalito, CA, USA, 2019. [Google Scholar]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Baisch, D.A.; Holmes, H.R. Developing a DNA barcode scanner for conservation. Genome 2017, 60, 910. [Google Scholar]
- Baerwald, M.; Goodbla, A.; Gootengerg, J.; Abudayyeh, O.; Schreier, A. Fast and Easy: CRISPR-Based Assays for Rapid and Accurate Species Genetic Identification. In Proceedings of the American Fisheries Society (AFS) and the Wildlife Society Joint Conference, Reno, NV, USA, 29 September–23 October 2019. [Google Scholar]
- Bernatchez, L.; Wellenreuther, M.; Araneda, C.; Ashton, D.T.; Barth, J.M.I.; Beacham, T.D.; Maes, G.E.; Martinsohn, J.T.; Miller, K.M.; Naish, K.A.; et al. Harnessing the Power of Genomics to Secure the Future of Seafood. Trends Ecol. Evol. 2017, 32, 665–680. [Google Scholar] [CrossRef]
- Ogden, R.; Linacre, A. Wildlife forensic science: A review of genetic geographic origin assignment. Forensic Sci. Int. Genet. 2015, 18, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Hasselman, D.; Anderson, E.C.; Argo, E.E.; Bethoney, N.D.; Gephard, S.R.; Post, D.M.; Schondelmeier, B.P.; Schultz, T.F.; Willis, T.V.; Palkovacs, E.P. Genetic stock composition of marine bycatch reveals disproportional impacts on depleted river herring genetic stocks. Can. J. Fish. Aquat. Sci. 2015, 73, 951–963. [Google Scholar] [CrossRef]
- Palkovacs, E.P.; Hasselman, D.J.; Argo, E.E.; Gephard, S.T.; Limburg, K.E.; Post, D.M.; Schultz, T.F.; Willis, T.V. Combining genetic and demographic information to prioritize conservation efforts for anadromous alewife and blueback herring. Evol. Appl. 2015, 7, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Bravington, M.V.; Grewe, P.M.; Davies, C.R. Absolute abundance of southern bluefin tuna estimated by close-kin mark-recapture. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Baetscher, D.; Anderson, E.C.; Gilbert-Horvath, E.A.; Malone, D.P.; Saarman, E.T.; Carr, M.H.; Garza, J.C. Dispersal of a nearshore marine fish connects marine reserves and adjacent fished areas along an open coast. Mol. Ecol. 2018. [Google Scholar] [CrossRef]
- Maloney, T.; Phelan, R.; Simmons, N. Saving the horseshoe crab: A synthetic alternative to horseshoe crab blood for endotoxin detection. PLoS Biol. 2018, 16, e2006607. [Google Scholar] [CrossRef]
- Fletcher, G.L.; Shears, M.A.; Yaskowiak, E.S.; King, M.J.; Goddard, S.V. Gene transfer: Potential to enhance the genome of Atlantic salmon for aquaculture. Aust. J. Exp. Agric. 2004, 44, 1095. [Google Scholar] [CrossRef]
- Du, S.J.; Gong, Z.; Fletcher, G.L.; Shears, M.A.; King, M.J.; Idler, D.R.; Hew, C.L. Growth Enhancement in Transgenic Atlantic Salmon by the Use of an “All Fish” Chimeric Growth Hormone Gene Construct. Bio/Technology 1992, 10, 176–181. [Google Scholar] [CrossRef]
- Aqua Bounty Technologies, Inc. Environmental Assessment for AquAdvantage Salmon. Available online: http://www.fda.gov/downloads/AdvisoryCommittees/CommitteesMeetingMaterials/VeterinaryMedicineAdvisoryCommittee/UCM224760 (accessed on 25 August 2010).
- Fox, K. Vegan Seafood Is About To Become Big Business—And Not A Moment Too Soon. 2018. Available online: https://www.forbes.com/sites/katrinafox/2018/08/06/vegan-seafood-is-about-to-become-big-business-and-not-a-moment-too-soon/#32e5ca26645d (accessed on 10 August 2019).
- Rubio, N.; Datar, I.; Stachura, D.; Kaplan, D.; Krueger, K. Cell-Based Fish: A Novel Approach to Seafood Production and an Opportunity for Cellular Agriculture. Front. Sustain. Food Syst. 2019, 3, 43. [Google Scholar] [CrossRef]
- Scherr, S.J.; McNeely, J.A. Biodiversity conservation and agricultural sustainability: Towards a new paradigm of “ecoagriculture” landscapes. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 477–494. [Google Scholar] [CrossRef]
- Reiley, L. Will lab-grown meat replace plant-based meat? Wash Post 2019. Available online: https://www.washingtonpost.com/business/2019/05/03/lab-table-will-cell-based-meat-win-over-americans/ (accessed on 10 July 2019).
- Liu, L.; Bilal, M.; Duan, X.; Iqbal, H.M.N. Mitigation of environmental pollution by genetically engineered bacteria—Current challenges and future perspectives. Sci. Total Environ. 2019, 667, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Dvorák, P.; Nicel, P.I.; Damborský, J.; Lorenzo, V. Bioremediation 3.0: Engineering pollutant-removing bacteria in the times of systemic biology. Biotechnol. Adv. 2017, 35, 845–866. [Google Scholar] [CrossRef] [PubMed]
- Wilkes, R.A.; Ludmilla, A. Degradation and metabolism of synthetic plastics and associated products by Pseudomonas sp.: Capabilities and challenges. J. Appl. Microbiol. 2017, 123, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A bacterium that degrades and assimilates poly (ethylene terephthalate). Science 2016, 351, 1196–1199. [Google Scholar]
- Urbanek, A.K.; Rymowicz, W.; Strzelecki, M.C.; Kociuba, W.; Franczak, Ł.; Mirończuk, A.M. Isolation and characterization of Arctic microorganisms decomposing bioplastics. AMB Express 2017, 7, 148. [Google Scholar] [CrossRef]
- Garlapati, D.; Charankumar, B.; Ramu, K.; Madeswaran, P.; Murthy, M.V.R. A review on the applications and recent advances in environmental DNA (eDNA) metagenomics. Rev. Environ. Sci. Bio/Technol. 2019, 18, 389–411. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Miaud, C.; Pompanon, R.; Taberlet, P. Species detection using environmental DNA from water samples. Biol. Lett. 2008, 4, 423–425. [Google Scholar] [CrossRef]
- Robson, H.L.; Noble, T.H.; Saunders, R.J.; Robson, S.K.A.; Burrows, D.W.; Jerry, D.R. Fine tuning for the tropics: Application of eDNA technology for invasive fish detection in tropical freshwater ecosystems. Mol. Ecol. Resour. 2016, 16, 922–932. [Google Scholar] [CrossRef]
- Jerde, C.L.; Mahon, A.R.; Chadderton, W.L.; Lodge, D.M. “Sight-Unseen” detection of rare aquatic species using environmental DNA. Conserv. Lett. 2011, 4, 150–157. [Google Scholar] [CrossRef]
- Ricciardi, A.; Blackburn, T.M.; Carlton, J.T.; Dick, J.T.A.; Hulme, P.E.; Iacarella, J.C.; Jeschke, J.M.; Liebhold, A.M.; Lockwood, J.L.; Maclsaac, H.J.; et al. Invasion science: A horizon scan of emerging challenges and opportunities. Trends Ecol. Evol. 2017, 32, 464–474. [Google Scholar] [CrossRef]
- Zaiko, A.; Martinez, J.L.; Schmidt-Petersen, J.; Ribicic, D.; Samuiloviene, A.; Garcia-Vazquez, E. Metabarcoding approach for the ballast water surveillance–an advantageous solution or an awkward challenge? Mar. Pollut. Bull. 2015, 92, 25–34. [Google Scholar] [CrossRef]
- Duncombe, L.G.; Therriault, T.W. Evaluating trapping as a method to control the European green crab, Carcinus maenas, population at Pipestem Inlet, British Columbia. Manag. Biol. Invasions 2017, 8, 235–246. [Google Scholar] [CrossRef]
- Morris, J.A., Jr. (Ed.) Invasive Lionfish: A Guide to Control and Management; Gulf and Caribbean Fisheries Institute Special Publication Series: Marathon, FL, USA, 2012; p. 113. [Google Scholar]
- Thresher, R.E.; Hayes, K.; Bax, N.J.; Teem, J.; Benfey, T.J.; Gould, F. Genetic control of invasive fish: Technological options and its role in integrated pest management. Biol. Invasions 2014, 16, 1201–1216. [Google Scholar] [CrossRef]
- Campbell, K.J.; Beek, J.; Eason, C.T.; Glen, A.S.; Godwin, J.; Gould, F.; Holmes, N.D.; Howald, G.R.; Madden, F.M.; Ponder, J.B.; et al. The next generation of rodent eradications: Innovative technologies and tools to improve species specificity and increase their feasibility on islands. Biol. Conserv. 2015, 185, 47–58. [Google Scholar] [CrossRef]
- Adams, J.S.; Alphey, L.; Bennett, E.L.; Brooks, T.M.; Delborne, J.; Eggermont, H.; Esvelt, K.; Kingiri, A.; Kokotovich, A.; Kolodziejczyk, B.; et al. Genetic frontiers for conservation: An assessment of synthetic biology and biodiversity conservation. IUCN Tech. Assess. 2019. [Google Scholar] [CrossRef]
- Thomas, D.D.; Donelly, C.A.; Wood, R.J.; Alphey, L.S. Insect Population Control Using a Dominant, Repressible, Lethal Genetic System. Science 2000, 287, 2474–2476. [Google Scholar] [CrossRef] [PubMed]
- Maselko, M.; Heinsch, S.; Das, S.; Smanski, M.J. Genetic incompatibility combined with female-lethality is effective and robust in simulations of Aedes aegypti. BioRxiv 2018. [Google Scholar] [CrossRef]
- Burt, A. Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Hammond, A.; Galizi, R.; Kyrou, K.; Simoni, A.; Siniscalchi, C.; Katsanos, D.; Gribble, M.; Baker, D.; Marois, E. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat. Biotechnol. 2016, 34, 78. [Google Scholar] [CrossRef] [PubMed]
- Noble, C.; Adlam, B.; Church, G.M.; Esvelt, K.M.; Nowak, M.A. Current CRISPR gene drive systems are likely to be highly invasive in wild populations. Elife 2018, 7, e33423. [Google Scholar] [CrossRef] [PubMed]
- Unckless, R.L.; Clark, A.G.; Messer, P.W. Evolution of resistance against CRISPR/Cas9 gene drive. Genetics 2017, 205, 827–841. [Google Scholar] [CrossRef] [PubMed]
- Leitschuh, C.M.; Kanavy, D.; Backus, G.A.; Valdez, R.X.; Serr, M.; Pitts, E.A.; Threadgill, D.; Godwin, J. Developing gene drive technologies to eradicate invasive rodents from islands. J. Responsible Innov. 2018, 5 (Suppl. 1), S121–S138. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Wilson, S.K.; Carr, P.; Hoey, A.S.; Jennings, S.; MacNeil, M.A. Seabirds enhance coral reef productivity and functioning in the absence of invasive rats. Nature 2018, 559, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Barshis, D.J.; Ladner, J.T.; Oliver, T.A.; Seneca, F.O.; Traylor-Knowles, N.; Palumbi, S.R. Genomic basis for coral resilience to climate change. Proc. Natl. Acad. Sci. USA 2013, 110, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Grummer, J.A.; Beheregaray, L.B.; Bernatchez, L.; Hand, B.K.; Luikart, S.R.; Taylor, E.B. Aquatic landscape genomics and environmental effects on genetic variation. Trends Ecol. Evol. 2019, 34, 641–654. [Google Scholar] [CrossRef]
- van Oppen, M.J.H.; Oliver, J.K.; Putnam, H.M.; Gates, R.D. Building coral reef resilience through assisted evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 2307–2313. [Google Scholar] [CrossRef]
- Hogdins, K.A.; Moore, J.L. Adapting to a warming world: Ecological restoration, climate change, and genomics. Am. J. Bot. 2016, 2016 103, 590–592. [Google Scholar]
- Hoffmann, A.; Griffin, P.; Dillon, S.; Catullo, R.; Rane, R.; Byrne, M.; Jordan, R.; Oakeshott, J.; Weeks, A.; Joseph, L.; et al. A framework for incorporating evolutionarygenomics into biodiversity conservation andmanagement. Clim. Chang. Responses 2015, 2, 1. [Google Scholar] [CrossRef]
- Heron, S.F.; Maynard, J.A.; van Hooidonk, R.; Eakin, C.M. Warming Trends and Bleaching Stress of the World’s Coral Reefs 1985-2012. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Heron, S.F.; van Hooidonk, R.; Maynard, J.; Anderson, K.; Day, J.C.; Geiger, E.; Hoegh-Guldberg, O.; Hughes, T.; Marshall, P.; Obura, D.; et al. Impacts of Climate Change on World Heritage Coral Reefs: A First Global Scientific Assessment; UNESCO World Herit Centre: Paris, French, 2017. [Google Scholar]
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral Reef Biodiversity. In Life in the World’s Oceans: Diversity, Distribution, and Abundance; McIntyre, A.E., Ed.; Blackwell Publishing Ltd: Hoboken, NJ, USA, 2015; pp. 65–78. [Google Scholar] [CrossRef]
- dela Cruz, D.W.; Harrison, P.L. Enhanced larval supply and recruitment can replenish reef corals on degraded reefs. Sci. Rep. 2017, 7, 13985. [Google Scholar] [CrossRef]
- Hagedorn, M.; Carter, V.L.; Henley, E.M.; van Oppen, M.J.H.; Hobbs, R.; Spindler, R.E. Producing Coral Offspring with Cryopreserved Sperm: A Tool for Coral Reef Restoration. Sci. Rep. 2017, 7, 14432. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Schopmeyer, S.; Lirman, D. A Review of Reef Restoration and Coral Propagation Using the Threatened Genus Acropora in the Caribbean and Western Atlantic. Bull. Mar. Sci. 2012, 88, 1075–1098. [Google Scholar] [CrossRef]
- Page, C.A.; Muller, E.M.; Vaughan, D.E. Microfragmenting for the successful restoration of slow growing massive corals. Ecol. Eng. 2018, 123, 86–94. [Google Scholar] [CrossRef]
- Forsman, Z.H.; Page, C.A.; Toonen, R.J.; Vaughan, D. Growing coral larger and faster: Micro-colony-fusion as a strategy for accelerating coral cover. Peer J. 2015, 3, e1313. [Google Scholar] [CrossRef]
- Craggs, J.; Guest, J.R.; Davis, M.; Simmons, J.; Dashti, E.; Sweet, M. Inducing broadcast coral spawning ex situ: Closed system mesocosm design and husbandry protocol. Ecol. Evol. 2017, 7, 11066–11078. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, R.J. The functional genomics laboratory: Functional validation of genetic variants. J. Inherit. Metab. Dis. 2018, 41, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Cleves, P.A.; Strader, M.E.; Bay, L.K.; Pringle, J.R.; Matz, M.V. CRISPR/Cas9-mediated genome editing in a reef-building coral. Proc. Natl. Acad. Sci. USA 2018, 115, 5235–5240. [Google Scholar] [CrossRef]
- Anthony, K.; Bay, L.K.; Costanza, R.; Firn, J.; Gunn, J.; Harrison, P.; Heyward, A.; Lundgren, P.; Mead, D.; Moore, T.; et al. New interventions are needed to save coral reefs. Nat. Ecol. Evol. 2017, 1, 1420–1422. [Google Scholar] [CrossRef]
- Laird, S.A.; Wynberg, R.P. Locating responsible research and innovation within access and benefit sharing spaces of the Convention on Biological Diversity: The challenge of emerging technologies. NanoEthics 2016, 10, 189–200. [Google Scholar] [CrossRef]
- Blasiak, R. International regulatory changes poised to reshape access to marine genes. Nat. Biotechnol. 2019, 37, 357. [Google Scholar] [CrossRef]
- Collins, J.E.; Harden-Davies, H.; Jaspars, M.; Thiele, T.; Vanagt, T.; Huys, I. Inclusive innovation: Enhancing global participation in and benefit sharing linked to the utilization of marine genetic resources from areas beyond national jurisdiction. Mar. Policy 2019, 109, 103696. [Google Scholar] [CrossRef]
- Russello, M.A.; Kirk, S.L.; Frazer, K.K.; Askey, P.J. Detection of outlier loci and their utility for fisheries management. Evol. Appl. 2012, 5, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Larson, W.A.; Seeb, L.W.; Everett, M.V.; Waples, R.K.; Templin, W.D.; Seeb, J.E. Genotyping by sequencing resolves shallow population structure to inform conservation of Chinook salmon (Oncorhynchus tshawytscha). Evol. Appl. 2014, 7, 355–369. [Google Scholar] [CrossRef]
- Reid, K.; Palkovacs, E.P.; Hasselman, D.J.; Baestcher, D.; Kibele, J.; Gahagan, B.; Bentzen, P.; McBride, M.C.; Garza, J.C. Comprehensive evaluation of genetic population structure for anadromous river herring with single nucleotide polymorphism data. Fish. Res. 2018, 206, 247–258. [Google Scholar] [CrossRef]
- Dixon, G.B.; Davies, S.W.; Aglyamova, G.V.; Meyer, E.; Bay, L.K.; Matz, M.V. Genomic determinants of coral heat tolerance across latitudes. Science 2015, 348, 1460–1462. [Google Scholar] [CrossRef]
- Zozaya-Valdes, E.; Egan, S.; Thomas, T. A comprehensive analysis of the microbial communities of healthy and diseased marine macroalgae and the detection of known and potential bacterial pathogens. Front. Microbiol. 2015, 6, 146. [Google Scholar] [CrossRef]
- Manel, S.; Perrier, C.; Pratlong, M.; Abi-Rached, L.; Paganini, J.; Pontarotti, P.; Aurelle, D. Genomic resources and their influence on the detection of the signal of positive selection in genome scans. Mol. Ecol. 2016, 25, 170–184. [Google Scholar] [CrossRef]
- WoRMS Editorial Board. World Register of Marine Species. Available online: http://www.marinespecies.org at VLIZ. (accessed on 18 October 2019).
- Braden, L. Activist journalists, NGOs disinformation campaign against approved, fast-growing GMO Salmon. Genet. Lit. Proj. 2016. Available online: https://geneticliteracyproject.org/2016/12/08/activist-journalists-ngos-continue-disinformation-campaign-approved-fast-growing-gmo-salmon/ (accessed on 5 October 2019).
- Ding, J.L.; Navas, M.A.A., III. Molecular cloning and sequence analysis of Factor C cDNA from the Singapore horseshoe crab, Carcinoscoprius rotundicauda. Mol. Mar. Biol. Biotechnol. 1995, 4, 90–103. [Google Scholar]
- Pui, A.W.M.; Ho, B.; Ding, J.L. Yeast recombinant Factor C from horseshoe crab binds endotoxin and causes bacteriostasis. J. Endotoxin Res. 1997, 4, 391–400. [Google Scholar] [CrossRef]
- Skerritt, J.; Shanker, D. Veggie Burgers Go Mainstream with Bloody Impossible Burger. Available online: https://www.bloomberg.com/news/articles/2017-06-05/bloody-meat-free-burger-brings-former-hippy-staple-to-mainstream (accessed on 18 October 2009).
- Stephens, N.; Di Silvio, L.; Dunsford, I.; Ellis, M.; Glencross, A.; Sexton, A. Bringing cultured meat to market: Technical, socio-political, and regulatory challenges in cellular agriculture. Trends Food Sci. Technol. 2018, 78, 155–166. [Google Scholar] [CrossRef]
- Zhang, B.; Oakes, A.D.; Newhouse, A.E.; Baier, K.M.; Maynard, C.A.; Powell, W.A. A threshold level of oxalate oxidase transgene expression reduces Cryphonectria parasitica-induced necrosis in a transgenic American chestnut (Castanea dentata) leaf bioassay. Transgenic Res. 2013, 22, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Esvelt, K.M.; Gemmel, N.J. Conservation demands save gene drive. PLOS Biol. 2017, 15, e2003850. [Google Scholar] [CrossRef] [PubMed]
- van der Sluijs, J.P.; Vaage, N.S. Pollinators and Global Food Security: The Need for Holistic Global Stewardship. Food Ethics 2016, 1, 75–91. [Google Scholar] [CrossRef]
- National Marine Fisheries Service. (2018) Fisheries of the United States, 2017. U.S. Department of Commerce, NOAA Current Fishery Statistics No. Available online: https://www.fisheries.noaa.gov/feature-story/ fisheries-united-states-2017 (accessed on 7 August 2019).
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Paarlberg, R. Starving for Science. How Biotechnology is Being Kept Out of Africa, 1st ed.; Harvard University Press: Cambridge, UK, 2008. [Google Scholar]
- Bailey, R. GM Scaremongering in Africa is Disarming the Fight against Poverty, the Guardian. Available online: https://www.theguardian.com/global-development/poverty-matters/2014/jul/21/gm-genetic-modification-crops-africa-eradicate-poverty (accessed on 10 November 2019).
- Wesseler, J.; Zilberman, D. The economic power of the Golden Rice opposition. Environ. Dev. Econ. 2014, 19, 724–742. [Google Scholar] [CrossRef]
- Lynas, M. Tanzania is burning GM corn while people go hungry. Little Atoms 2017. Available online: http://littleatoms.com/science-world/tanzania-burning-GM-corn-while-people-go-hungry (accessed on 10 October 2019).
- Bryant, D.; Burke, L.; McManus, J.W.; Spalding, M.D. (Eds.) Reefs at Risk: A Map-Based Indicator of Threats to the World’s Coral Reefs; World Resources Institute: New York, NY, USA, 1998. [Google Scholar]
- Wernberg, T.; Straub, S.C. Impacts and Effects of Ocean Warming on Seaweeds. In Explaining Ocean Warming: Causes, Scale, Effects and Consequences; Laffoley, D., Baxter, J.M., Eds.; International Union for the Conservation of Nature: Gland, Switzerland, 2016; pp. 87–101. [Google Scholar]
- Harrison, A.L.; Costa, D.P.; Winship, A.J.; Benson, S.R.; Bograd, S.J.; Antolos, M.; Carlisle, A.B.; Dewar, H.; Dutton, P.H.; Jorgensen, S.J.; et al. The political biogeography of migratory marine predators. Nat. Ecol. Evol. 2018, 2, 1571–1578. [Google Scholar] [CrossRef]
- Batavia, C.; Nelson, M.P. For Goodness Sake! What Is Intrinsic Value and Why Should We Care? Biol. Conserv. 2017, 209, 366–376. [Google Scholar] [CrossRef]
- Novak, B.J.; Maloney, T.; Phelan, R. Advancing a New Toolkit for Conservation: From Science to Policy. CRISPR J. 2018, 1, 11–15. [Google Scholar] [CrossRef]
- Funk, W.C.; Forester, B.R.; Converse, S.J.; Darst, C.; Morey, S. Improving conservation policy with genomics: A guide to integrating adaptive potential into U.S. Endangered Species Act decisions for conservation practitioners and geneticists. Conserv. Genet. 2019, 20, 115–134. [Google Scholar] [CrossRef]
- UN Report: Nature’s Dangerous Decline ‘Unprecedented’; Species Extinction Rates ‘Accelerating’. Available online: https://www.un.org/sustainabledevelopment/blog/2019/05/nature-decline-unprecedented-report/ (accessed on 31 October 2019).




{kind=link}
{kind=link}
{kind=link}
| Genomic Initiative | Taxonomic Focus | Target | Status |
|---|---|---|---|
| Earth Biogenome Project (EBP) | All known eukaryote species | 1,500,000 | In Process |
| Darwin Tree of Life | All known UK eukaryote species | 66,000 | In Process |
| Vertebrate Genomes Project (VGP) | All known vertebrate species | 70,000 | In-process |
| G10k | One species of each vertebrate genus | 10,000 | Transitioned to VGP |
| B10K | All known bird species | 10,000 | In-process |
| Bat1k | All known bat species | 1300 | In-process |
| Global Invertebrate Genomics Alliance (GIGA) | Marine invertebrates | 7000 | In-process |
| i5k | Arthropods, primarily insects | 5000 | In-process |
| 1000 Fungal Genomes Project | Fungal species | 1000 | In-process |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novak, B.J.; Fraser, D.; Maloney, T.H. Transforming Ocean Conservation: Applying the Genetic Rescue Toolkit. Genes 2020, 11, 209. https://doi.org/10.3390/genes11020209
Novak BJ, Fraser D, Maloney TH. Transforming Ocean Conservation: Applying the Genetic Rescue Toolkit. Genes. 2020; 11(2):209. https://doi.org/10.3390/genes11020209
Chicago/Turabian StyleNovak, Ben J., Devaughn Fraser, and Thomas H. Maloney. 2020. "Transforming Ocean Conservation: Applying the Genetic Rescue Toolkit" Genes 11, no. 2: 209. https://doi.org/10.3390/genes11020209
APA StyleNovak, B. J., Fraser, D., & Maloney, T. H. (2020). Transforming Ocean Conservation: Applying the Genetic Rescue Toolkit. Genes, 11(2), 209. https://doi.org/10.3390/genes11020209