Uncovering Phenotypic Diversity and DArTseq Marker Loci Associated with Antioxidant Activity in Common Bean

 ,
,  ,
, 

 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Field Experiment and Phenotypic Evaluation
2.3. DNA Extraction and Genotyping for DArTseq Markers
2.4. Extraction and Determination of Total Antioxidant Activity
2.5. Statistical Analysis
2.5.1. Phenotypic Data Analysis
2.5.2. Population Structure and GWAS Analysis
2.5.3. Putative Candidate Gene Analysis
3. Results
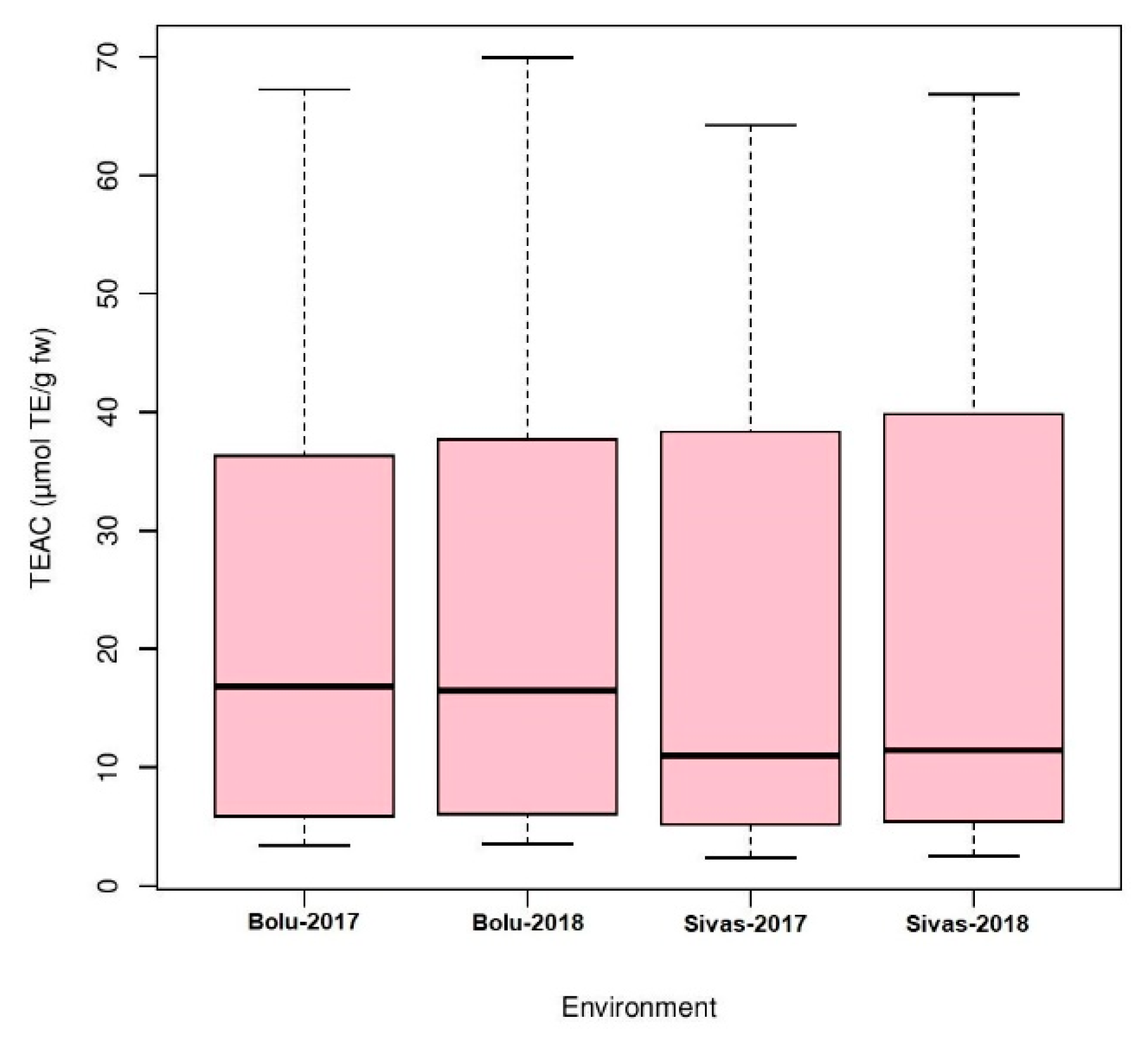
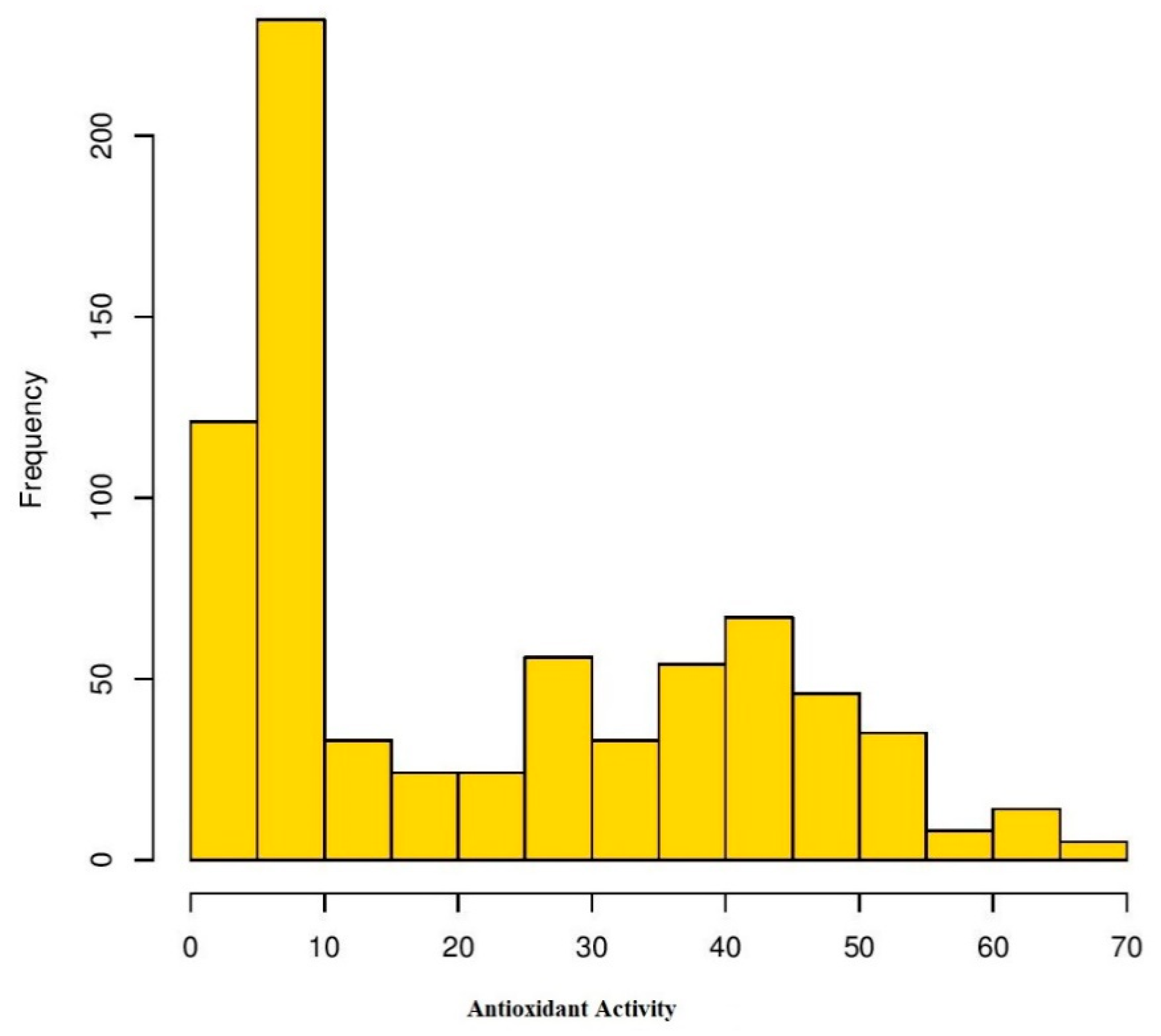
3.1. Phenotypic Diversity Evaluation
3.2. Population Structure and LD
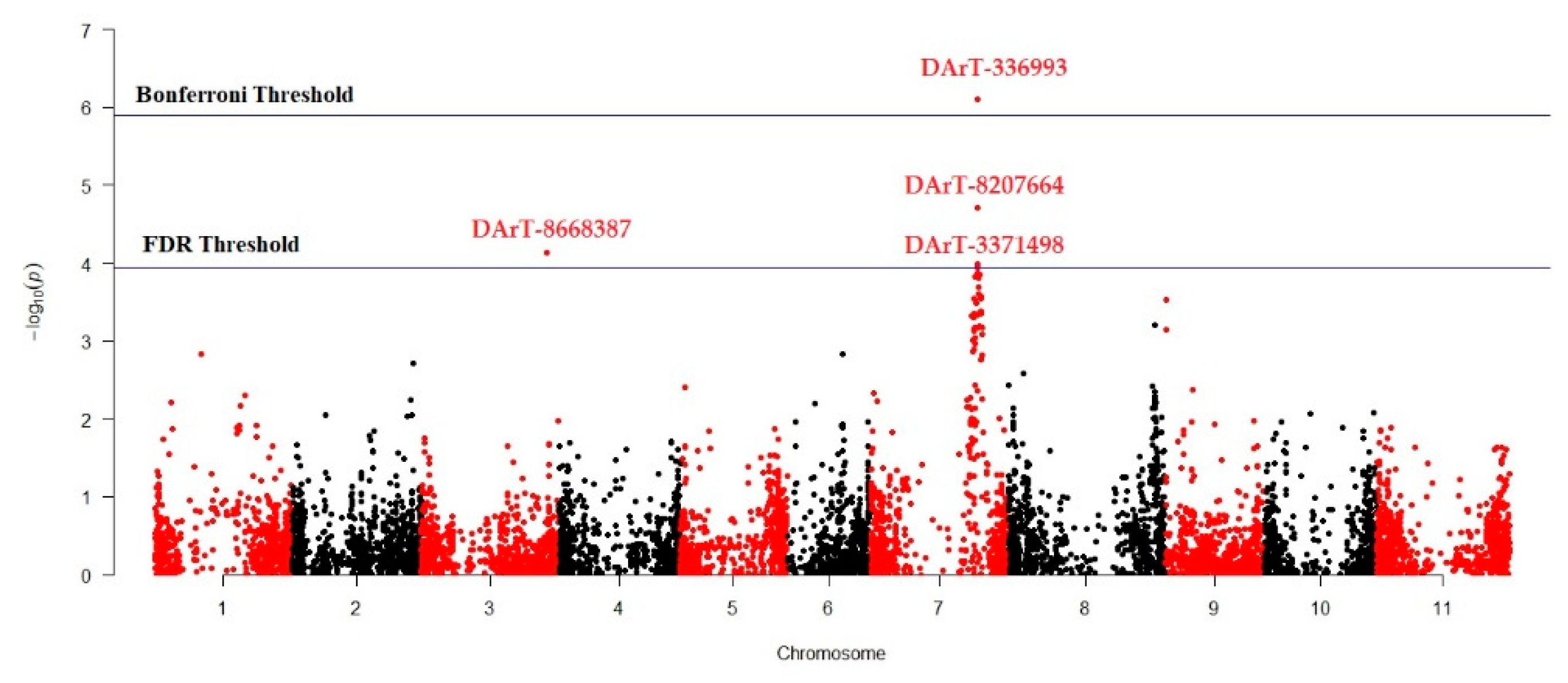
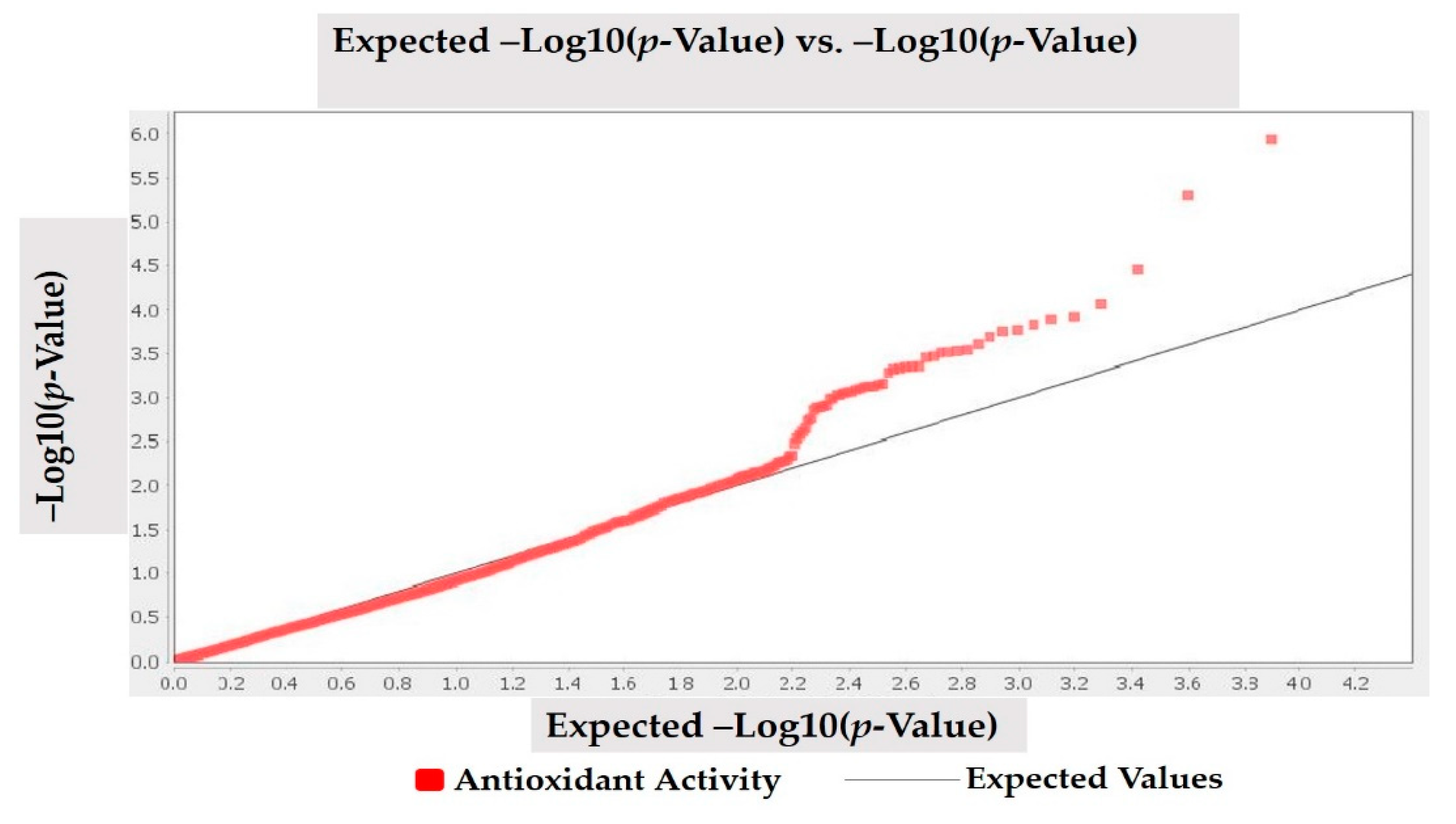
3.3. Marker Trait Association for Antioxidant Activity
3.4. Putative Candidate Genes for Antioxidant Activity
4. Discussion
4.1. DArTseq Markers Analysis and LD
4.2. Marker Trait Association and Putative Candidate Genes for Antioxidant Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Long, S.P.; Marshall-Colon, A.; Zhu, X.G. Meeting the global food demand of the future by engineering crop photosynthesis and yield potential. Cell 2015, 161, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization. 2012. Available online: http://www.faostat.fao.org/site/339/default.aspx> (accessed on 5 September 2018).
- Jensen, E.S.; Peoples, M.B.; Boddey, R.M.; Gresshoff, P.M.; Hauggaard-Nielsen, H.; Alves, B.J.; Morrison, M.J. Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agron. Sustain. Dev. 2012, 32, 329–364. [Google Scholar] [CrossRef]
- Petry, N.; Boy, E.; Wirth, J.; Hurrell, R. The potential of the common bean (Phaseolus vulgaris) as a vehicle for iron biofortification. Nutrients 2015, 7, 1144–1173. [Google Scholar] [CrossRef] [PubMed]
- Celmeli, T.; Sari, H.; Canci, H.; Sari, D.; Adak, A.; Eker, T.; Toker, C. The nutritional content of common bean (Phaseolus vulgaris L.) landraces in comparison to modern varieties. Agronomy 2018, 8, 166. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. FAO Statistical Yearbook. 2016. Available online: http://www.fao.org/docrep/017/i3138e/i3138e.pdf (accessed on 5 September 2018).
- Bitocchi, E.; Bellucci, E.; Giardini, A.; Rau, D.; Rodriguez, M.; Biagetti, E.; Santilocchi, R.; Spagnoletti Zeuli, P.; Gioia, T.; Logozzo, G.; et al. Molecular analysis of the parallel domestication of the common bean (P haseolus vulgaris) in Mesoamerica and the Andes. New Phytol. 2013, 197, 300–313. [Google Scholar] [CrossRef]
- Bozoglu, H.; Sozen, O. A sample for biodiversity in Turkey: Common bean (Phaseolus vulgaris L.) landraces from Artvin. Afr. J. Biotechnol. 2011, 10, 13789–13796. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Habyarimana, E.; Çiftçi, V.; Nawaz, M.A.; Karaköy, T.; Comertpay, G.; Shahid, M.Q.; Hatipoğlu, R.; Yeken, M.Z.; Ali, F.; et al. Characterization of genetic diversity in Turkish common bean gene pool using phenotypic and whole-genome DArTseq-generated silicoDArT marker information. PLoS ONE 2018, 13, 0205363. [Google Scholar] [CrossRef]
- Ozturk, I.; Kara, M.; Yildiz, C.; Ercisli, S. Physico-mechanical seed properties of the common Turkish bean (Phaseolus vulgaris) cultivars ‘Hinis’ and ‘Ispir’. N. Z. J. Crop Hort. Sci. 2009, 37, 41–50. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotec. Eq. 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef]
- Moghaddam, S.M.; Mamidi, S.; Osorno, J.M.; Lee, R.; Brick, M.; Kelly, J.; Miklas, P.; Urrea, C.; Song, Q.; Cregan, P.; et al. Genome-wide association study identifies candidate loci underlying agronomic traits in a Middle American diversity panel of common bean. Plant Genome 2016, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Hofmann, N.; Li, S.; Ferreira, M.E.; Song, B.; Jiang, G.; Ren, S.; Quigley, C.; Fickus, E.; Cregan, P.; et al. Identification of QTL with large effect on seed weight in a selective population of soybean with genome-wide association and fixation index analyses. BMC Genom. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
- Perseguini, J.M.; Oblessuc, P.R.; Rosa, J.R.; Gomes, K.A.; Chiorato, A.F.; Carbonell, S.A.; Garcia, A.A.; Vianello, R.P.; Benchimol-Reis, L.L. Genome-wide association studies of anthracnose and angular leaf spot resistance in common bean (Phaseolus vulgaris L.). PLoS ONE 2016, 11, e0150506. [Google Scholar] [CrossRef] [PubMed]
- Cichy, K.A.; Wiesinger, J.A.; Mendoza, F.A. Genetic diversity and genome-wide association analysis of cooking time in dry bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2015, 128, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Fritsche-Neto, R.; Souza, T.L.; Pereira, H.S.; Faria, L.C.; Melo, L.C.; Novaes, E.; Brum, I.J.; Jannink, J.L. Association mapping in common bean revealed regions associated with Anthracnose and Angular Leaf Spot resistance. Sci. Agric. 2019, 76, 321–327. [Google Scholar] [CrossRef]
- Hoyos-Villegas, V.; Song, Q.; Kelly, J.D. Genome-wide association analysis for drought tolerance and associated traits in common bean. Plant Genome 2016, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mamta, M.K.; Dhillon, G.S.; Brar, S.K.; Verma, M. Antioxidants. In Biotransformation of Waste Biomass into High Value Biochemicals; Brar, S., Dhillon, G., Soccol, C., Eds.; Springer: New York, NY, USA, 2014; pp. 117–138. [Google Scholar]
- Pooja, V.; Sunita, M. Antioxidants and disease prevention. Int. J. Adv. Sci. Tech. Res. 2014, 2, 903–911. [Google Scholar]
- Sudhir, S.; Budhiraja, R.D.; Miglani, G.P.; Arora, B.; Gupta, L.C.; Garg, K.N. Pharmacological studies on leaves of Withania somnifera. Planta Med. 1986, 52, 61–63. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Vaseghi, G.; Pourfarzam, M.; Abdollahi, A. Are antioxidants helpful for disease prevention? Res. Pharm. Sci. 2010, 5, 1–8. [Google Scholar]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative Stress and Neurodegenerative Diseases: A Review of Upstream and Downstream Antioxidant Therapeutic Options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Sikora, E.; Cieślik, E.; Topolska, K. The sources of natural antioxidants. Acta Sci. Pol. Technol. Aliment. 2008, 7, 5–17. [Google Scholar]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutr. J. 2010, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- La Vecchia, C.; Altieri, A.; Tavani, A. Vegetables, fruit, antioxidants and cancer: A review of Italian studies. Eur. J. Nutr. 2001, 40, 261–267. [Google Scholar] [CrossRef]
- Kusvuran, S.; Kiran, S.; Ellialtioglu, S.S. Antioxidant enzyme activities and abiotic stress tolerance relationship in vegetable crops. In Abiotic and Biotic Stress in Plants-Recent Advances and Future Perspectives, 1st ed.; InTech: London, UK, 2016; pp. 481–503. [Google Scholar] [CrossRef]
- Dolatabadian, A.; Jouneghani, R.S. Impact of exogenous ascorbic acid on antioxidant activity and some physiological traits of common bean subjected to salinity stress. Not. Bot. Horti Agrobot. Cluj Napoca. 2009, 37, 165–172. [Google Scholar] [CrossRef]
- Silvana, B.D.; Susanna, M.G.; María, P.B.; María, L.T. Behaviour of antioxidant defense system in the adaptive response to salt stress in Helianthus annuus L. cells. Plant Growth Regul. 2003, 40, 81–88. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, J.; Cai, S.; Chen, X.; Quan, X.; Zhang, G. Association mapping for total polyphenol content, total flavonoid content and antioxidant activity in barley. BMC Genom. 2018, 19, 81. [Google Scholar] [CrossRef]
- Xu, F.; Bao, J.; Kim, T.S.; Park, Y.J. Genome-wide association mapping of polyphenol contents and antioxidant capacity in whole-grain rice. J. Agric. Food Chem. 2016, 64, 4695–4703. [Google Scholar] [CrossRef]
- Cardador-Martinez, A.; Castano-Tostado, E.; Loarca-Pina, G. Antimutagenic activity of natural phenolic compounds present in the common bean (Phaseolus vulgaris) against aflatoxin B 1. Food Addit. Contam. 2002, 19, 62–69. [Google Scholar] [CrossRef]
- Weidner, S.; Król, A.; Karamać, M.; Amarowicz, R. Phenolic compounds and the antioxidant properties in seeds of green-and yellow-podded bean (Phaseolus vulgaris L.) varieties. CyTA J. Food 2018, 16, 373–380. [Google Scholar] [CrossRef]
- Khaidizar, M.I.; Haliloglu, K.; Elkoca, E.; Aydin, M.; Kantar, F. Genetic diversity of common bean (Phaseolus vulgaris L.) landraces grown in northeast Anatolia of Turkey assessed with simple sequence repeat markers. Turk. J. Field Crops. 2012, 17, 145–150. [Google Scholar]
- Ceylan, A.; Öcal, N.; Akbulut, M. Genetic diversity among the Turkish common bean cultivars (Phaseolus vulgaris L.) as assessed by SRAP, POGP and cpSSR markers. Biochem. Syst. Ecol. 2014, 54, 219–229. [Google Scholar] [CrossRef]
- Federer, W.T.; Reynolds, M.; Crossa, J. Combining results from augmented designs over sites. Agron. J. 2001, 93, 389–395. [Google Scholar] [CrossRef]
- Boyle, C.R.; Montgomery, R.D. An application of the augmented randomized complete block design to poultry research. Poult. Sci. 1996, 75, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Baloch, F.S.; Alsaleh, A.; Andeden, E.E.; Hatipoğlu, R.; Nachit, M.; Özkan, H. High levels of segregation distortion in the molecular linkage map of bread wheat representing the West Asia and North Africa region. Turk. J. Agric. For. 2016, 40, 352–364. [Google Scholar] [CrossRef]
- Saracoglu, O.; Ozturk, B.; Yildiz, K.; Kucuker, E. Pre-harvest methyl jasmonate treatments delayed ripening and improved quality of sweet cherry fruits. Sci. Hort. 2017, 226, 19–23. [Google Scholar] [CrossRef]
- Rathore, A.; Parsad, R.; Gupta, V.K. Computer aided construction and analysis of augmented designs. J. Indian Soc. Agric. Stat. 2004, 57, 320–344. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons Inc.: New York, NY, USA, 1984. [Google Scholar]
- Habyarimana, E. Genomic prediction for yield improvement and safeguarding of genetic diversity in CIMMYT spring wheat (Triticum aestivum L.). Aust. J. Crop. Sci. 2016, 10, 127–136. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 15 April 2019).
- Payne, R.W.; Harding, S.A.; Murray, D.A.; Soutar, D.M.; Baird, D.B.; Glaser, A.I.; Welham, S.J.; Gilmour, A.R.; Thompson, R.; Webster, R. The Guide to Gen Stat Release 14, Part 2: Statistics; VSN International: Hemel Hempstead, UK, 2011. [Google Scholar]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Achenbach, U.; Paulo, J.; Ilarionova, E.; Lübeck, J.; Strahwald, J.; Tacke, E.; Hofferbert, H.R.; Gebhardt, C. Using SNP markers to dissect linkage disequilibrium at a major quantitative trait locus for resistance to the potato cyst nematode Globodera pallida on potato chromosome V. Theor. Appl. Genet. 2009, 118, 619–629. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hill, W.G.; Robertson, A. Linkage disequilibrium in finite populations. Theor. Appl. Genet. 1968, 38, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Birlea, S.A.; Fain, P.R.; Gowan, K.; Riccardi, S.L.; Holland, P.J.; Bennett, D.C.; Herbstman, D.M.; Wallace, M.R.; McCormack, W.T.; et al. Genome-wide analysis identifies a quantitative trait locus in the MHC class II region associated with generalized vitiligo age of onset. J. Investig. Dermatol. 2011, 131, 1308–1312. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.D. qqman: An R package for visualizing GWAS results using QQ and manhattan plots. Biorxiv 2014, 005165. [Google Scholar] [CrossRef]
- Huber, K.; Brigide, P.; Bretas, E.B.; Canniatti-Brazaca, S.G. Phenolic Acid, Flavonoids and Antioxidant Activity of Common Beans (Phaseolus vulgaris L.) Before and After Cooking. J. Nutr. Food Sci. 2016, 6, 551. [Google Scholar] [CrossRef]
- Bojilov, D.G.; Manolov, S.P.; Bozadzhiev, B.V.; Stremski, J.I.; Ivanov, I.I. Antioxidant activity in two species common beans (Phaseolus vulgaris L.) from village of smilyan, Bulgaria. Agric. Food 2018, 6, 314–324. [Google Scholar]
- García-Díaz, Y.D.; Aquino-Bolaños, E.N.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C. Bioactive compounds and antioxidant activity in the common bean are influenced by cropping season and genotype. Chil. J. Agric. Res. 2018, 78, 255–265. [Google Scholar] [CrossRef]
- Ullah, F.; Alam, A.; Yamamoto, S.; Kahn, N.; Honna, T. Screening for agronomic performance of six indigenous cultivars of alfalfa (Medicago sativa) at Karina northern areas of Pakistan. Electron. J. Environ. Agric. Food Chem. 2009, 8, 950–968. [Google Scholar]
- Di Silvestro, R.; Di Loreto, A.; Bosi, S.; Bregola, V.; Marotti, I.; Benedettelli, S.; Segura-Carretero, A.; Dinelli, G. Environment and genotype effects on antioxidant properties of organically grown wheat varieties: A 3-year study. J. Sci. Food Agric. 2017, 97, 641–649. [Google Scholar] [CrossRef]
- Khamphasan, P.; Lomthaisong, K.; Harakotr, B.; Ketthaisong, D.; Scott, M.; Lertrat, K.; Suriharn, B. Genotypic variation in anthocyanins, phenolic compounds, and antioxidant activity in cob and husk of purple field corn. Agronomy 2018, 8, 271. [Google Scholar] [CrossRef]
- Boateng, J.; Verghese, M.; Walker, L.T.; Ogutu, S. Effect of processing on antioxidant contents in selected dry beans (Phaseolus spp. L.). LWT-Food Sci. Technol. 2008, 41, 1541–1547. [Google Scholar] [CrossRef]
- Ranilla, L.G.; Genovese, M.I.; Lajolo, F.M. Polyphenols and antioxidant capacity of seed coat and cotyledon from Brazilian and Peruvian bean cultivars (Phaseolus vulgaris L.). J. Agric. Food Chem. 2007, 55, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Capistrán-Carabarin, A.; Aquino-Bolaños, E.N.; García-Díaz, Y.D.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C. Complementarity in Phenolic Compounds and the Antioxidant Activities of Phaseolus coccineus L. and P. vulgaris L. Landraces. Foods 2019, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Briñez, B.; Blair, M.W.; Kilian, A.; Carbonell, S.A.; Chiorato, A.F.; Rubiano, L.B. A whole genome DArT assay to assess germplasm collection diversity in common beans. Mol. Breed. 2012, 30, 181–193. [Google Scholar] [CrossRef]
- Mogga, M.; Sibiya, J.; Shimelis, H.; Lamo, J.; Yao, N. Diversity analysis and genome-wide association studies of grain shape and eating quality traits in rice (Oryza sativa L.) using DArT markers. PloS ONE 2018, 13, e0198012. [Google Scholar] [CrossRef]
- Aydin, M.F.; Baloch, F.S. Exploring the genetic diversity and population structure of Turkish common bean germplasm by the iPBS-retrotransposons markers. Legum. Res. 2019, 42, 18–24. [Google Scholar] [CrossRef]
- Flint-Garcia, S.A.; Thornsberry, J.M.; Buckler, E.S., 4th. Structure of linkage disequilibrium in plants. Annu. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef]
- Hamblin, M.T.; Mitchell, S.E.; White, G.M.; Gallego, J.; Kukatla, R.; Wing, R.A.; Paterson, A.H.; Kresovich, S. Comparative population genetics of the panicoid grasses: Sequence polymorphism, linkage disequilibrium and selection in a diverse sample of Sorghum bicolor. Genetics 2004, 167, 471–483. [Google Scholar] [CrossRef]
- Remington, D.L.; Thornsberry, J.M.; Matsuoka, Y.; Wilson, L.M.; Whitt, S.R.; Doebley, J.; Kresovich, S.; Goodman, M.M.; Buckler, E.S. Structure of linkage disequilibrium and phenotypic associations in the maize genome. Proc. Natl. Acad. Sci. USA 2001, 98, 11479–11484. [Google Scholar] [CrossRef]
- Campa, A.; Murube, E.; Ferreira, J. Genetic diversity, population structure, and linkage disequilibrium in a spanish common bean diversity panel revealed through genotyping-by-sequencing. Genes 2018, 9, 518. [Google Scholar] [CrossRef]
- Ates, D.; Asciogul, T.K.; Nemli, S.; Erdogmus, S.; Esiyok, D.; Tanyolac, M.B. Association mapping of days to flowering in common bean (Phaseolus vulgaris L.) revealed by DArT markers. Mol. Breed. 2018, 38, 113–127. [Google Scholar] [CrossRef]
- Wang, H.; Xu, X.; Vieira, F.G.; Xiao, Y.; Li, Z.; Wang, J.; Nielsen, R.; Chu, C. The power of inbreeding: NGS-based GWAS of rice reveals convergent evolution during rice domestication. Mol. Plant 2016, 9, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Sun, J.; Gong, D.; Kong, Y. The Roles of Arabidopsis C1-2i Subclass of C2H2-type Zinc-Finger Transcription Factors. Genes 2019, 10, 653. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Muhovski, Y.; Žižková, E.; Dobrev, P.I.; Franco-Zorrilla, J.M.; Solano, R.; Lopez-Vidriero, I.; Motyka, V.; Lutts, S. The Solanum lycopersicum Zinc Finger2 cysteine-2/histidine-2 repressor-like transcription factor regulates development and tolerance to salinity in tomato and Arabidopsis. Plant Physiol. 2014, 164, 1967–1990. [Google Scholar] [CrossRef]
- Cao, H.; Huang, P.; Zhang, L.; Shi, Y.; Sun, D.; Yan, Y.; Liu, X.; Dong, B.; Chen, G.; Snyder, J.H.; et al. Characterization of 47 Cys2-His2 zinc finger proteins required for the development and pathogenicity of the rice blast fungus Magnaporthe oryzae. New Phytol. 2016, 211, 1035–1051. [Google Scholar] [CrossRef]
- Cheuk, A.; Houde, M. Genome wide identification of C1-2i zinc finger proteins and their response to abiotic stress in hexaploid wheat. Mol. Genet. Genom. 2016, 291, 873–890. [Google Scholar] [CrossRef]
- Zang, D.; Li, H.; Xu, H.; Zhang, W.; Zhang, Y.; Shi, X.; Wang, Y. An Arabidopsis zinc finger protein increases abiotic stress tolerance by regulating sodium and potassium homeostasis, reactive oxygen species scavenging and osmotic potential. Front. Plant Sci. 2016, 7, 1272. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.; Wen, F.; Yao, D.; Wang, L.; Guo, J.; Ni, L.; Zhang, A.; Tan, M.; Jiang, M. A novel rice C2H2-type zinc finger protein, ZFP36, is a key player involved in abscisic acid-induced antioxidant defence and oxidative stress tolerance in rice. J. Exp. Bot. 2014, 65, 5795–5809. [Google Scholar] [CrossRef]
- Bowles, D.; Lim, E.K.; Poppenberger, B.; Vaistij, F.E. Glycosyltransferases of lipophilic small molecules. Annu. Rev. Plant Biol. 2006, 57, 567–597. [Google Scholar] [CrossRef]
- Li, P.; Li, Y.J.; Zhang, F.J.; Zhang, G.Z.; Jiang, X.Y.; Yu, H.M.; Hou, B.K. The Arabidopsis UDP-glycosyltransferases UGT79B2 and UGT79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. Plant J. 2017, 89, 85–103. [Google Scholar] [CrossRef]
- Rehman, H.M.; Nawaz, M.A.; Shah, Z.H.; Ludwig-Müller, J.; Chung, G.; Ahmad, M.Q.; Yang, S.H.; Lee, S.I. Comparative genomic and transcriptomic analyses of Family-1 UDP glycosyltransferase in three Brassica species and Arabidopsis indicates stress-responsive regulation. Sci. Rep. 2018, 8, 1875. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.G.; Wang, B.; Jin, S.H.; Qu, X.X.; Li, Y.J.; Hou, B.K. Ectopic expression of Arabidopsis glycosyltransferase UGT85A5 enhances salt stress tolerance in tobacco. PLoS ONE 2013, 8, 59924. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.R.; Lima, R.P.; Lisboa, J.V.; Chaves, T.R.; Luna, R.C.; do Nascimento, R.A.; de Oliveira, Y.; Persuhn, D.C.; Sérgio da Silva, A.; Gonçalves, M.D.; et al. Influence of the C677T polymorphism of the MTHFR gene on oxidative stress in women with overweight or obesity: Response to a dietary folate intervention. J. Am. Coll. Nutr. 2018, 37, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Raeisi, J.; Pakkish, Z.; Saffari, V.R. Efficiency of Folic Acid in Improving Yield and Fruit Quality of Strawberry. J. Plant Physiol. Breed. 2017, 7, 15–25. [Google Scholar]
- Small, I.D.; Peeters, N. The PPR motif—a TPR-related motif prevalent in plant organellar proteins. Trends Biochem. Sci. 2000, 25, 45–47. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, H.; Jia, Z.; Fang, Q.; Luo, K. Combined expression of antimicrobial genes (Bbchit1 and LJAMP2) in transgenic poplar enhances resistance to fungal pathogens. Tree Physiol. 2012, 32, 1313–1320. [Google Scholar] [CrossRef]
- Zsigmond, L.; Rigó, G.; Szarka, A.; Székely, G.; Ötvös, K.; Darula, Z.; Medzihradszky, K.F.; Koncz, C.; Koncz, Z.; Szabados, L. Arabidopsis PPR40 connects abiotic stress responses to mitochondrial electron transport. Plant Physiol. 2008, 146, 1721–1737. [Google Scholar] [CrossRef]
- Chen, G.; Zou, Y.; Hu, J.; Ding, Y. Genome-wide analysis of the rice PPR gene family and their expression profiles under different stress treatments. BMC Genom. 2018, 19, 720. [Google Scholar] [CrossRef]
- Yaldiz, G.; Camlica, M.; Nadeem, M.A.; Nawaz, M.A.; Baloch, F.S. Genetic diversity assessment in Nicotiana tabacum L. with iPBS-retrotransposons. Turk. J. Agric. For. 2018, 42, 154–164. [Google Scholar] [CrossRef]
- Guliyev, N.; Sharifova, S.; Ojaghi, J.; Abbasov, M.; Akparov, Z. Genetic diversity among melon (Cucumis melo L.) accessions revealed by morphological traits and ISSR markers. Turk. J. Agric. For. 2018, 42, 393–401. [Google Scholar] [CrossRef]
- Gramazio, P.; Plesa, I.M.; Truta, A.M.; Sestras, A.F.; Vilanova, S.; Plazas, M.; Vicente, O.; Boscaiu, M.; Prohens, J.; Sestras, R.E. Highly informative SSR genotyping reveals large genetic diversity and limited differentiation in European larch (Larix decidua) populations from Romania. Turk. J. Agric. For. 2018, 42, 165–175. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Df | Sum Sq | Mean Sq | F Value | Pr (>F) |
|---|---|---|---|---|---|
| LINE | 187 | 199487 | 1066.8 | 11.937 | <2 × 10−16 |
| ENV | 3 | 115 | 38.2 | 0.427 | 0.734 |
| Residuals | 561 | 50133 | 89.4 | ||
| Heritability | 0.92 |
| Variables | White | Purple | Brown | Beige | Yellow | Dark red | Black | Antioxidant |
|---|---|---|---|---|---|---|---|---|
| White | 1 | −0.448 ** | −0.328 ** | −0.427 ** | −0.236 ** | −0.253 ** | −0.177 | −0.422 ** |
| Purple | 1 | −0.105 | −0.137 | −0.076 | −0.081 | −0.057 | 0.108 | |
| Brown | 1 | −0.100 | −0.055 | −0.059 | −0.042 | 0.214 ** | ||
| Beige | 1 | −0.072 | −0.077 | −0.054 | 0.309 ** | |||
| Yellow | 1 | −0.043 | −0.030 | −0.014 | ||||
| Dark red | 1 | −0.032 | 0.060 | |||||
| Black | 1 | 0.080 | ||||||
| Antioxidant | 1 |
| Chr. | Number of Markers | Mean Distance (Mbs) | r2 | Significant LD (%) | LD Decay (Mb) |
|---|---|---|---|---|---|
| 1 | 658 | 12.61 | 0.52 | 53.76 | 1.2 |
| 2 | 931 | 18.99 | 0.48 | 56.47 | 1.2 |
| 3 | 863 | 16.53 | 0.51 | 53.61 | 1.15 |
| 4 | 632 | 13.80 | 0.41 | 54.58 | 1.18 |
| 5 | 659 | 16.38 | 0.39 | 59.16 | 1.3 |
| 6 | 591 | 18.48 | 0.50 | 55.16 | 1.15 |
| 7 | 680 | 13.15 | 0.45 | 49.55 | 1.08 |
| 8 | 863 | 14.47 | 0.46 | 55.4 | 1.15 |
| 9 | 637 | 17.03 | 0.59 | 45.89 | 1.05 |
| 10 | 599 | 13.86 | 0.45 | 50.62 | 1.1 |
| 11 | 787 | 15.68 | 0.46 | 59.8 | 1.2 |
| 7900 | 15.45 | 0.47 | 54.0 | 1.15 |
| Marker | Chromosome | Position | p-Value | R2 | Putative Gene |
|---|---|---|---|---|---|
| DArT-3369938 | 7 | 40398124 | 7.93 × 10−7 | 0.15657 | Phvul.009G180200.2 |
| DArT-8668387 | 3 | 47385021 | 7.40 × 10−5 | 0.09497 | Phvul.003G239700 |
| DArT-8207664 | 7 | 40398185 | 1.93 × 10−5 | 0.12493 | Phvul.007G281200.1 |
| DArT-3371498 | 7 | 40409459 | 1.03 × 10−4 | 0.09404 | Phvul.007G171700.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadeem, M.A.; Gündoğdu, M.; Ercişli, S.; Karaköy, T.; Saracoğlu, O.; Habyarimana, E.; Lin, X.; Hatipoğlu, R.; Nawaz, M.A.; Sameeullah, M.; et al. Uncovering Phenotypic Diversity and DArTseq Marker Loci Associated with Antioxidant Activity in Common Bean. Genes 2020, 11, 36. https://doi.org/10.3390/genes11010036
Nadeem MA, Gündoğdu M, Ercişli S, Karaköy T, Saracoğlu O, Habyarimana E, Lin X, Hatipoğlu R, Nawaz MA, Sameeullah M, et al. Uncovering Phenotypic Diversity and DArTseq Marker Loci Associated with Antioxidant Activity in Common Bean. Genes. 2020; 11(1):36. https://doi.org/10.3390/genes11010036
Chicago/Turabian StyleNadeem, Muhammad Azhar, Müttalip Gündoğdu, Sezai Ercişli, Tolga Karaköy, Onur Saracoğlu, Ephrem Habyarimana, Xiao Lin, Ruştu Hatipoğlu, Muhammad Amjad Nawaz, Muhammad Sameeullah, and et al. 2020. "Uncovering Phenotypic Diversity and DArTseq Marker Loci Associated with Antioxidant Activity in Common Bean" Genes 11, no. 1: 36. https://doi.org/10.3390/genes11010036
APA StyleNadeem, M. A., Gündoğdu, M., Ercişli, S., Karaköy, T., Saracoğlu, O., Habyarimana, E., Lin, X., Hatipoğlu, R., Nawaz, M. A., Sameeullah, M., Ahmad, F., Jung, B.-M., Chung, G., & Baloch, F. S. (2020). Uncovering Phenotypic Diversity and DArTseq Marker Loci Associated with Antioxidant Activity in Common Bean. Genes, 11(1), 36. https://doi.org/10.3390/genes11010036