Genome-Wide Identification and Transcriptional Expression of the METTL21C Gene Family in Chicken

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of METTL21C in Sequenced Genomes of G. gallus
2.2. METTL21C Protein Alignment, Phylogenetic Analysis, Pfam Domain Detection, and Chromosome Location Analysis of G. gallus METTL21C Genes
2.3. Gene Structure and Motif Analysis of METTL21C Genes
2.4. The Materials, Treatments, and Collections of Different Breeds of Chickens
2.5. Materials, Treatments, and Collection of the Transcriptome Profile of the METTL21C Gene Gamily of Chickens
2.6. Quantitative Real-Time PCR
3. Results
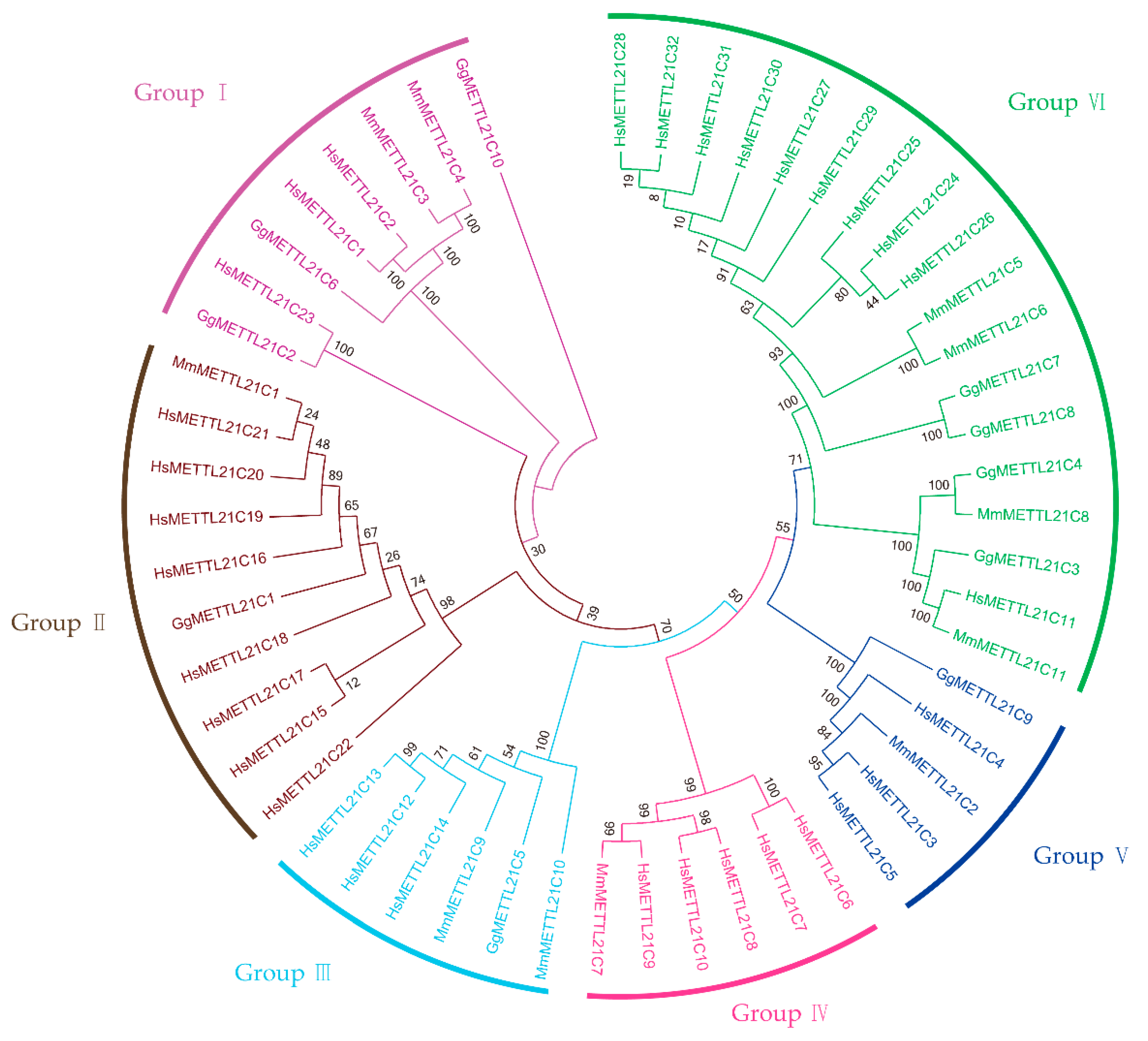
3.1. Identification and Phylogenetic Analysis of METTL21C Genes
3.2. Chromosomal Locations of METTL21C Genes and Their Relationship with G. gallus
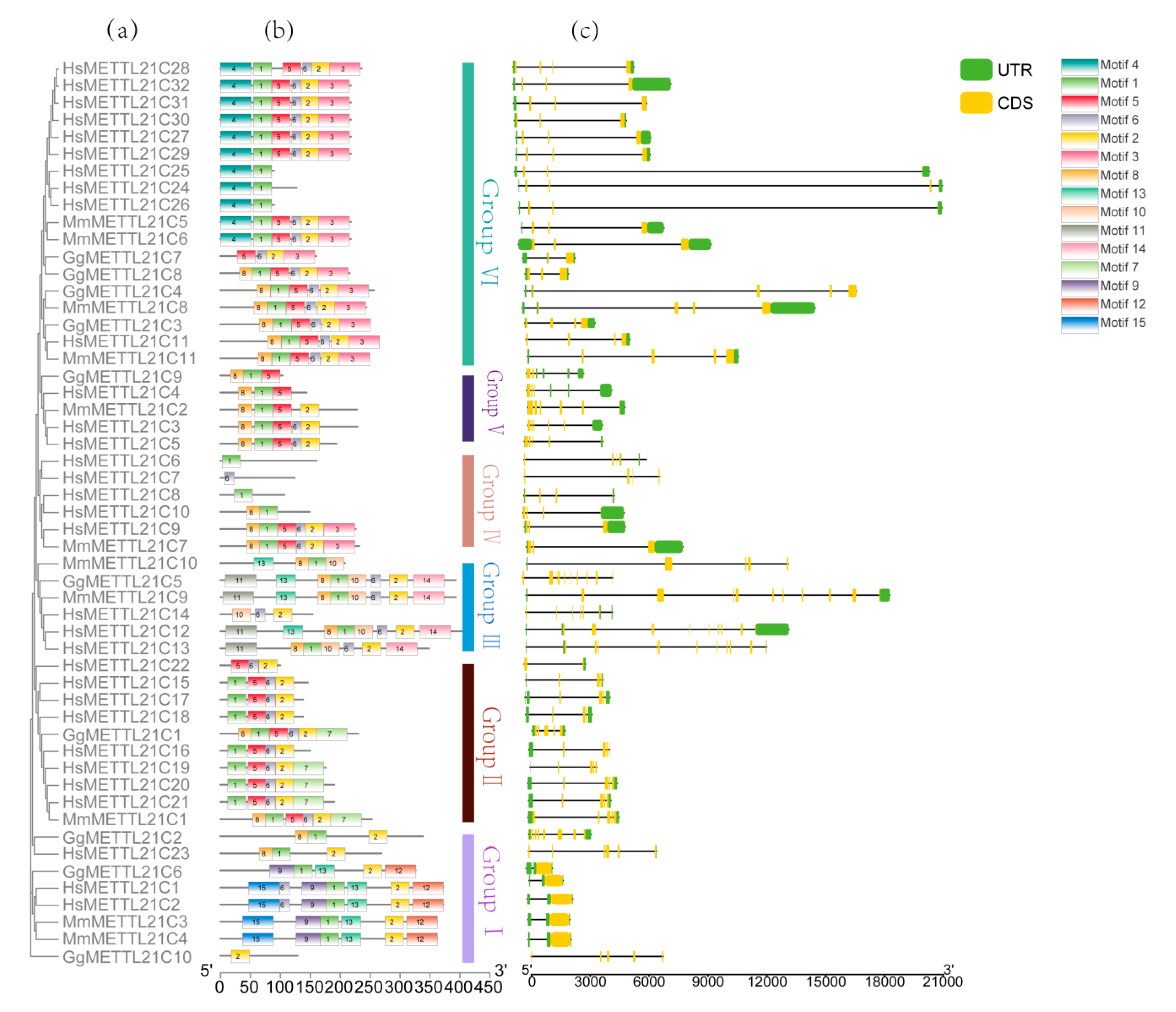
3.3. METTL21C Gene Structures and Conserved Motifs
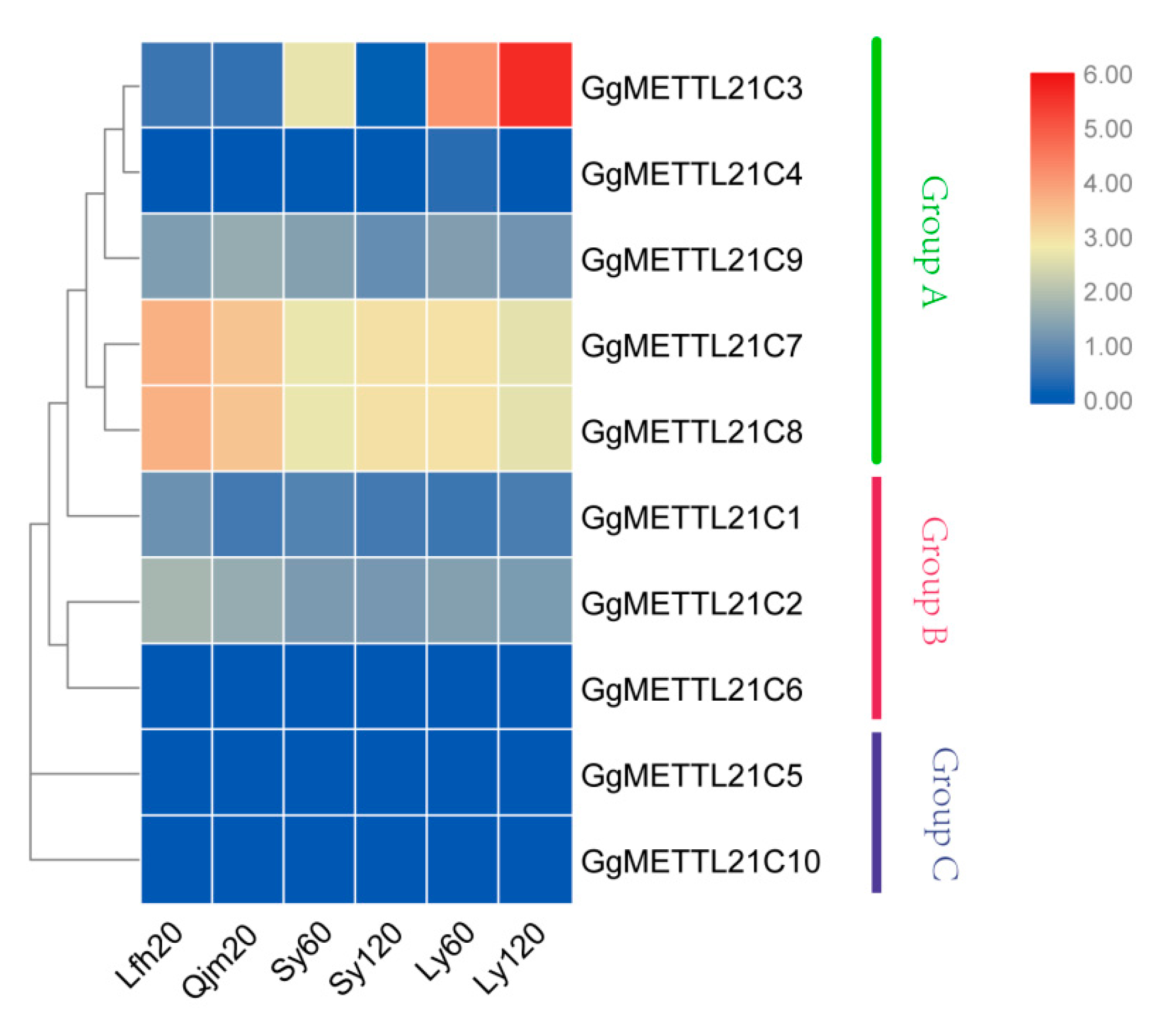
3.4. Expression Profile Analysis of G. gallus METTL21C Genes
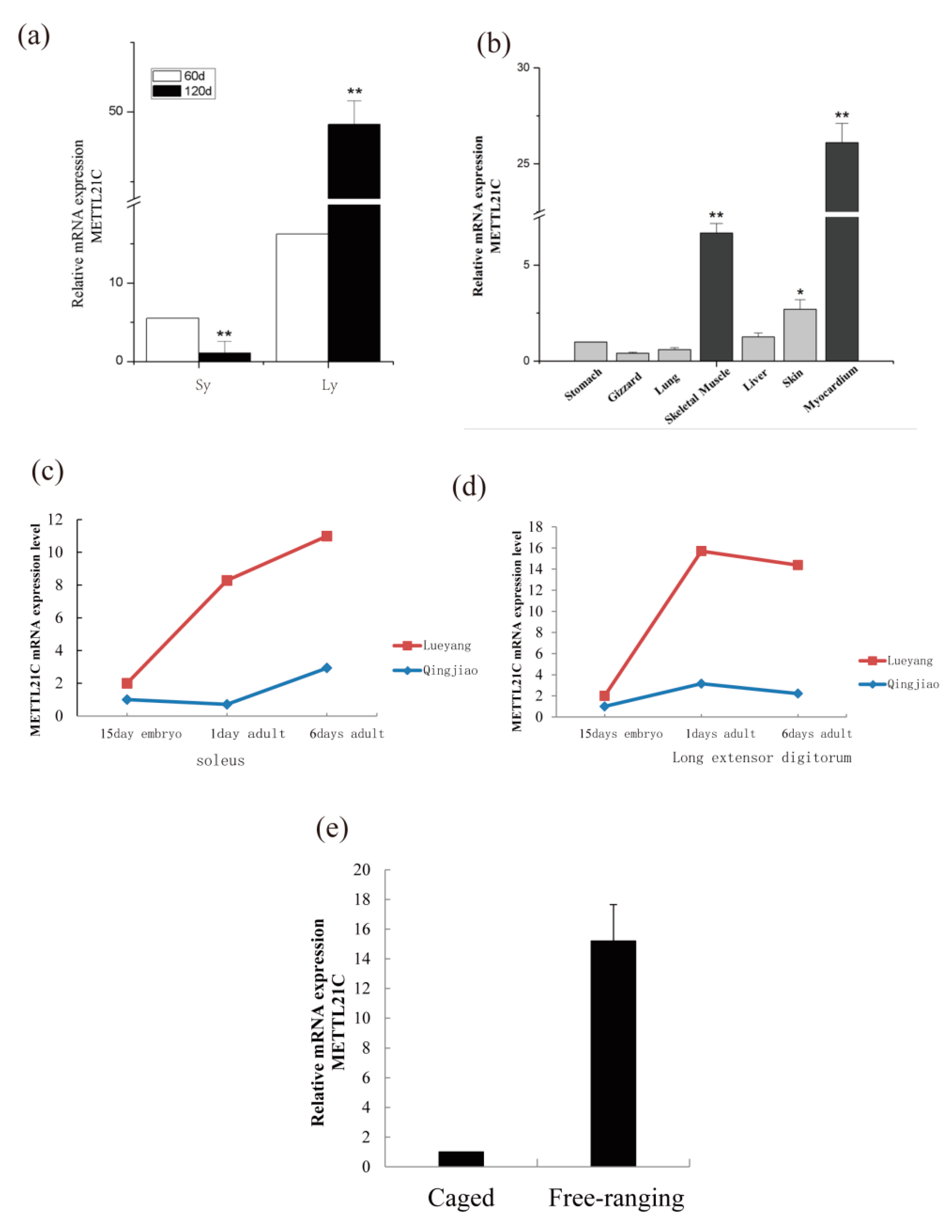
3.5. Real-Time PCR of METTL21C Genes of the METTL21C Gene Family
4. Discussion
4.1. The Characteristics of the METTL21C Gene Family
4.2. The Evolution History of the METTL21C Gene Family of Chickens, Humans, and Mice
4.3. The Function of the METTL21C Gene Family of Chickens
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fumihito, A.; Miyake, T.; Sumi, S.; Takada, M.; Ohno, S.; Kondo, N. One subspecies of the red junglefowl (Gallus gallus gallus) suffices as the matriarchic ancestor of all domestic breeds. Proc. Natl. Acad. Sci. USA 1994, 91, 12505–12509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, H.; Yang, L.K.; Yin, Y.J.; Lu, H.Z.; Wang, L. The complete mitochondrial genome and molecular phylogeny of Lueyang black-bone chicken. Br. Poult. Sci. 2018, 59, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Fleming, R.H.; Whitehead, C.C.; Alvey, D.; Gregory, N.G.; Wilkins, L.J. Bone structure and breaking strength in laying hens housed in different husbandry systems. Br. Poult. Sci. 1994, 35, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Wiederstein, J.L.; Nolte, H.; Günther, S.; Piller, T.; Baraldo, M.; Kostin, S.; Bloch, W.; Schindler, N.; Sandri, M.; Blaauw, B.; et al. Skeletal Muscle-Specific Methyltransferase METTL21C Trimethylates p97 and Regulates Autophagy-Associated Protein Breakdown. Cell Rep. 2018, 23, 1342–1356. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, P.; Lavallée-Adam, M.; Faubert, D.; Blanchette, M.; Coulombe, B. A Newly Uncovered Group of Distantly Related Lysine Methyltransferases Preferentially Interact with Molecular Chaperones to Regulate Their Activity. PLoS Genet. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Kernstock, S.; Davydova, E.; Jakobsson, M.; Moen, A.; Pettersen, S.; Maelandsmo, G.M.; Egge-Jacobsen, W.; Falnes, P.Ø. Lysine methylation of VCP by a member of a novel human protein methyltransferase family. Nat. Commun. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.L.; McMillan, F.M. SAM (dependent) I AM: The S-adenosylmethionine-dependent methyltransferase fold. Curr. Opin. Struct. Biol. 2002, 12, 783–793. [Google Scholar] [CrossRef]
- Schubert, H.L.; Blumenthal, R.M.; Cheng, X. Many paths to methyltransfer: A chronicle of convergence. Trends Biochem. Sci. 2003, 28, 329–335. [Google Scholar] [CrossRef]
- Mosquera-Rendón, J.; Cárdenas-Brito, S.; Pineda, J.D.; Corredor, M.; Benítez-Páez, A. Evolutionary and sequence-based relationships in bacterial AdoMet-dependent non-coding RNA methyltransferases. BMC Res. Notes 2014, 7, 440. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef]
- Huang, J.; Hsu, Y.H.; Mo, C.; Abreu, E.; Kiel, D.P.; Bonewald, L.F.; Brotto, M.; Karasik, D. METTL21C is a potential pleiotropic gene for osteoporosis and sarcopenia acting through the modulation of the NF-kappaB signaling pathway. J. Bone Miner. Res. 2014, 29, 1531–1540. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Gao, L.H.; Li, S.S.; Wei, Z.Y.; Fu, W.Z.; He, J.W.; Liu, Y.J.; Hu, Y.Q.; Dong, J.; Zhang, Z.L. Association between SNPs and haplotypes in the METTL21C gene and peak bone mineral density and body composition in Chinese male nuclear families. J. Bone Miner. Metab. 2017, 35, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.; Fry, A.N.; Kang, M. The complexities of p97 function in health and disease. Mol. Biosyst. 2011, 7, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Raasi, S.; Wolf, D.H. Ubiquitin receptors and ERAD: A network of pathways to the proteasome. Semin. Cell Dev. Biol. 2007, 18, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Pye, V.E.; Dreveny, I.; Briggs, L.C.; Sands, C.; Beuron, F.; Zhang, X.; Freemont, P.S. Going through the motions: The ATPase cycle of p97. J. Struct. Biol. 2006, 156, 12–28. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y. Diverse functions with a common regulator: Ubiquitin takes command of an AAA ATPase. J. Struct. Biol. 2006, 156, 29–40. [Google Scholar] [CrossRef]
- Latterich, M.; Fröhlich, K.U.; Schekman, R. Membrane fusion and the cell cycle: Cdc48p participates in the fusion of ER membranes. Cell 1995, 82, 885–893. [Google Scholar] [CrossRef]
- Ye, Y.; Meyer, H.H.; Rapoport, T.A. The AAA ATPase Cdc48/p97 and its partners transport proteins from the ER into the cytosol. Nature 2001, 414, 652–656. [Google Scholar] [CrossRef]
- Indig, F.E.; Partridge, J.J.; von Kobbe, C.; Aladjem, M.I.; Latterich, M.; Bohr, V.A. Werner syndrome protein directly binds to the AAA ATPase p97/VCP in an ATP-dependent fashion. J. Struct. Biol. 2004, 146, 251–259. [Google Scholar] [CrossRef]
- Partridge, J.J.; Lopreiato, J.O.; Latterich, M.; Indig, F.E. DNA damage modulates nucleolar interaction of the Werner protein with the AAA ATPase p97/VCP. Mol. Biol. Cell 2003, 14, 4221–4229. [Google Scholar] [CrossRef]
- Watts, G.D.; Wymer, J.; Kovach, M.J.; Mehta, S.G.; Mumm, S.; Darvish, D.; Pestronk, A.; Whyte, M.P.; Kimonis, V.E. Inclusion body myopathy associated with Paget disease of bone and frontotemporal dementia is caused by mutant valosin-containing protein. Nat. Genet. 2004, 36, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Weihl, C.C.; Dalal, S.; Pestronk, A.; Hanson, P.I. Inclusion body myopathy-associated mutations in p97/VCP impair endoplasmic reticulum-associated degradation. Hum. Mol. Genet. 2006, 15, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Thiele, W.; Novac, N.; Mink, S.; Schreiber, C.; Plaumann, D.; Fritzmann, J.; Cremers, N.; Rothley, M.; Schwager, C.; Regiert, T.; et al. Discovery of a novel tumour metastasis-promoting gene, NVM-1. J. Pathol. 2011, 225, 96–105. [Google Scholar] [CrossRef] [PubMed]
- López Hernández, J.F.; Zanders, S.E. Veni, vidi, vici: The success of wtf meiotic drivers in fission yeast. Yeast 2018, 35, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Hillier, L.D.; Miller, W.; Birney, E.; Warren, W.; Hardison, R.C.; Ponting, C.P.; Bork, P.; Burt, D.W.; Groenen, M.A.M.; Delany, M.E.; et al. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar]
- Church, D.M.; Goodstadt, L.; Hillier, L.W.; Zody, M.C.; Goldstein, S.; She, X.; Bult, C.J.; Agarwala, R.; Cherry, J.L.; DiCuccio, M.; et al. Lineage-specific biology revealed by a finished genome assembly of the mouse. PLoS Biol. 2009, 7, e1000112. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.Y.; Levy-Sakin, M.; Kwok, P.Y. De novo human genome assemblies reveal spectrum of alternative haplotypes in diverse populations. Nat. Commun. 2018, 9, 3040. [Google Scholar] [CrossRef]
- Wang, G.M.; Yin, H.; Qiao, X.; Tan, X.; Gu, C.; Wang, B.H.; Cheng, R.; Wang, Y.Z.; Zhang, S.L. F-box genes: Genome-wide expansion, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri). Plant Sci. 2016, 253, 164–175. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Woeste, K.E.; Zhao, P. Completion of the Chloroplast Genomes of Five Chinese Juglans and Their Contribution to Chloroplast Phylogeny. Front. Plant Sci. 2016, 7, 1955. [Google Scholar] [CrossRef] [PubMed]
- Sassa, H.; Kakui, H.; Miyamoto, M.; Suzuki, Y.; Hanada, T.; Ushijima, K.; Kusaba, M.; Hirano, H.; Koba, T. S locus F-box brothers: Multiple and pollen-specific F-box genes with S haplotype-specific polymorphisms in apple and Japanese pear. Genetics 2007, 175, 1869–1881. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. BioTechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. HemI: A toolkit for illustrating heatmaps. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Kakui, H.; Kato, M.; Ushijima, K.; Kitaguchi, M.; Kato, S.; Sassa, H. Sequence divergence and loss-of-function phenotypes of S locus F-box brothers genes are consistent with non-self recognition by multiple pollen determinants in self-incompatibility of Japanese pear (Pyrus pyrifolia). Plant J. 2011, 68, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Piao, Y.; Shon, H.S.; Ryu, K.H. Comparing the normalization methods for the differential analysis of Illumina high-throughput RNA-Seq data. BMC Bioinform. 2015, 16, 347. [Google Scholar] [CrossRef]
- Maag, J.L.V. Gganatogram: An R package for modular visualisation of anatograms and tissues based on ggplot2. F1000Research 2018, 7, 1576. [Google Scholar] [CrossRef]
- Feng, S.; Tan, H.; Ling, H.; Yuan, X. Detecting overexpression level of HER2 gene in NSCLC by real-time quantitative PCR and the 2[-Delta Delta C(T)] method. Chin. J. Lung Cancer 2011, 14, 938–942. [Google Scholar]
- Expósito-Rodríguez, M.; Borges, A.A.; Borges-Pérez, A.; Pérez, J.A. Selection of internal control genes for quantitative real-time RT-PCR studies during tomato development process. BMC Plant Biol. 2008, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Petrossian, T.C.; Clarke, S.G. Uncovering the human methyltransferasome. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.W.; Peng, M.S.; Wu, G.S.; Ouyang, Y.N.; Yang, Z.Y.; Yu, N.; Liang, J.P.; Pianchou, G.; Beja-Pereira, A.; Mitra, B.; et al. Chicken domestication: An updated perspective based on mitochondrial genomes. Heredity 2013, 110, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Burt, D.W.; Carrë, W.; Fell, M.; Law, A.S.; Antin, P.B.; Maglott, D.R.; Weber, J.A.; Schmidt, C.J.; Burgess, S.C.; McCarthy, F.M. The Chicken Gene Nomenclature Committee report. BMC Genom. 2009, 10, S5. [Google Scholar] [CrossRef] [PubMed]
- Reiff, R.E.; Ali, B.R.; Baron, B.; Yu, T.W.; Ben-Salem, S.; Coulter, M.E.; Schubert, C.R.; Hill, R.S.; Akawi, N.A.; Al-Younes, B.; et al. METTL23, a transcriptional partner of GABPA, is essential for human cognition. Hum. Mol. Genet. 2014, 23, 3456–3466. [Google Scholar] [CrossRef] [PubMed]
- Bernkopf, M.; Webersinke, G.; Tongsook, C.; Koyani, C.N.; Rafiq, M.A.; Ayaz, M.; Müller, D.; Enzinger, C.; Aslam, M.; Naeem, F.; et al. Disruption of the methyltransferase-like 23 gene METTL23 causes mild autosomal recessive intellectual disability. Hum. Mol. Genet. 2014, 23, 4015–4023. [Google Scholar] [CrossRef]
- Davydova, E.; Ho, A.Y.Y.; Malecki, J.; Moen, A.; Enserink, J.M.; Jakobsson, M.E.; Loenarz, C.; Falnes, P.Ø. Identification and characterization of a novel evolutionarily conserved lysine-specific methyltransferase targeting eukaryotic translation elongation factor 2 (eEF2). J. Biol. Chem. 2014, 289, 30499–30510. [Google Scholar] [CrossRef]
- Kaul, G.; Pattan, G.; Rafeequi, T. Eukaryotic elongation factor-2 (eEF2): Its regulation and peptide chain elongation. Cell Biochem. Funct. 2011, 29. [Google Scholar] [CrossRef]
- Cloutier, P.; Lavallée-Adam, M.; Faubert, D.; Blanchette, M.; Coulombe, B. Methylation of the DNA/RNA-binding protein Kin17 by METTL22 affects its association with chromatin. J. Proteom. 2014, 100, 115–124. [Google Scholar] [CrossRef]
- Jakobsson, M.E.; Moen, A.; Bousset, L.; Egge-Jacobsen, W.; Kernstock, S.; Melki, R.; Falnes, P.Ø. Identification and characterization of a novel human methyltransferase modulating Hsp70 protein function through lysine methylation. J. Biol. Chem. 2013, 288, 27752–27763. [Google Scholar] [CrossRef] [PubMed]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Gorlova, O.; Fedorov, A.; Logothetis, C.; Amos, C.; Gorlov, I. Genes with a large intronic burden show greater evolutionary conservation on the protein level. BMC Evol. Biol. 2014, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Deutsch, M. Association of intron phases with conservation at splice site sequences and evolution of spliceosomal introns. Mol. Biol. Evol. 1999, 16, 1528–1534. [Google Scholar] [CrossRef]
- Dinant, S.; Clark, A.M.; Zhu, Y.; Vilaine, F.; Palauqui, J.C.; Kusiak, C.; Thompson, G.A. Diversity of the superfamily of phloem lectins (phloem protein 2) in angiosperms. Plant. Physiol. 2003, 131, 114–128. [Google Scholar] [CrossRef]
- Bai, C.; Sen, P.; Hofmann, K.; Ma, L.; Goebl, M.; Harper, J.W.; Elledge, S.J. SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box. Cell 1996, 86, 263–274. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein ID | CDS ID | Gene ID | Chromosome | Gene Position | |
|---|---|---|---|---|---|---|
| Start | End | |||||
| GgMETTL21C1 | ENSGALP00000002756.3 | ENSGALT00000002759.6 | ENSGALG00000001790.6 | chr18 | 4,245,590 | 4,247,698 |
| GgMETTL21C2 | ENSGALP00000003173.1 | ENSGALT00000003178.2 | ENSGALG00000002044.5 | chr14 | 13,793,041 | 13,797,471 |
| GgMETTL21C3 | ENSGALP00000027192.5 | ENSGALT00000027243.7 | ENSGALG00000016864.7 | chr1 | 144,281,168 | 144,285,163 |
| GgMETTL21C4 | ENSGALP00000027185.6 | ENSGALT00000027236.7 | ENSGALG00000016859.7 | chr1 | 144,171,934 | 144,190,015 |
| GgMETTL21C5 | ENSGALP00000053406.2 | ENSGALT00000048292.2 | ENSGALG00000033603.3 | chr14 | 10,511,222 | 10,519,988 |
| GgMETTL21C6 | ENSGALP00000047914.1 | ENSGALT00000056245.2 | ENSGALG00000035963.2 | chr8 | 5,470,226 | 5,471,907 |
| GgMETTL21C7 | ENSGALP00000063245.1 | ENSGALT00000086206.2 | ENSGALG00000008493.7 | chr7 | 12,179,296 | 12,182,133 |
| GgMETTL21C8 | ENSGALP00000032767.3 | ENSGALT00000033408.5 | ENSGALG00000008493.7 | chr7 | 12,179,725 | 12,182,133 |
| GgMETTL21C9 | ENSGALP00000020014.7 | ENSGALT00000020041.7 | ENSGALG00000012269.7 | chr5 | 57,863,106 | 57,866,374 |
| GgMETTL21C10 | ENSGALP00000073336.1 | ENSGALT00000098371.1 | ENSGALG00000051460.1 | chr5 | 57,862,387 | 57,869,575 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Lu, H.; Wang, L.; Zhao, J.; Zeng, W.; Zhang, T. Genome-Wide Identification and Transcriptional Expression of the METTL21C Gene Family in Chicken. Genes 2019, 10, 628. https://doi.org/10.3390/genes10080628
Yang G, Lu H, Wang L, Zhao J, Zeng W, Zhang T. Genome-Wide Identification and Transcriptional Expression of the METTL21C Gene Family in Chicken. Genes. 2019; 10(8):628. https://doi.org/10.3390/genes10080628
Chicago/Turabian StyleYang, Ge, Hongzhao Lu, Ling Wang, Jiarong Zhao, Wenxian Zeng, and Tao Zhang. 2019. "Genome-Wide Identification and Transcriptional Expression of the METTL21C Gene Family in Chicken" Genes 10, no. 8: 628. https://doi.org/10.3390/genes10080628
APA StyleYang, G., Lu, H., Wang, L., Zhao, J., Zeng, W., & Zhang, T. (2019). Genome-Wide Identification and Transcriptional Expression of the METTL21C Gene Family in Chicken. Genes, 10(8), 628. https://doi.org/10.3390/genes10080628