Cloning and Functional Analysis of Lignin Biosynthesis Genes Cf4CL and CfCCoAOMT in Cryptomeria fortunei

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction of Total RNA and Synthesis of First Strand cDNA
2.3. Molecular Cloning and Genetic Transformation of Tobacco
2.4. Bioinformatics Analysis
2.5. Expression Analysis
2.6. Transgenic Plant Growth and Phenotyping
2.7. Determination of Lignin Content
3. Results
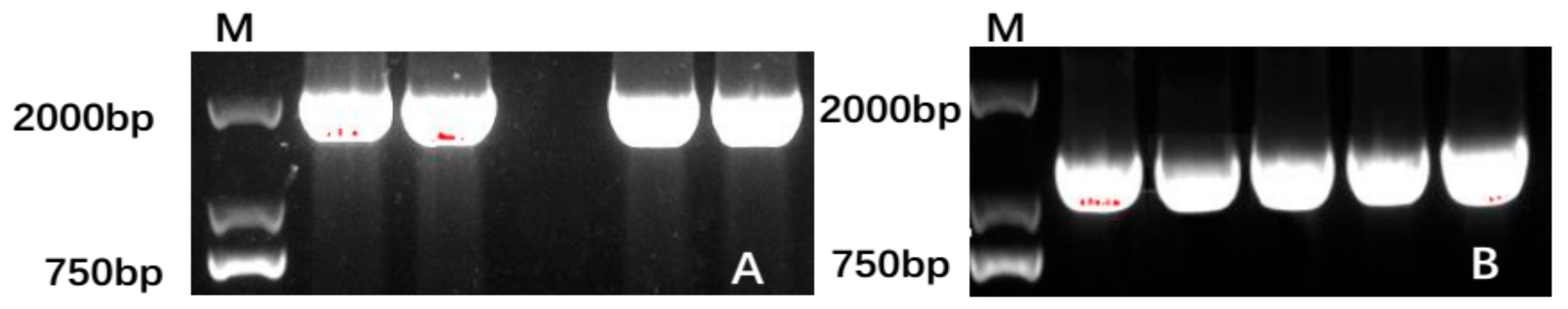
3.1. C. fortunei Contains Cf4CL and CfCCoAOMT Genes
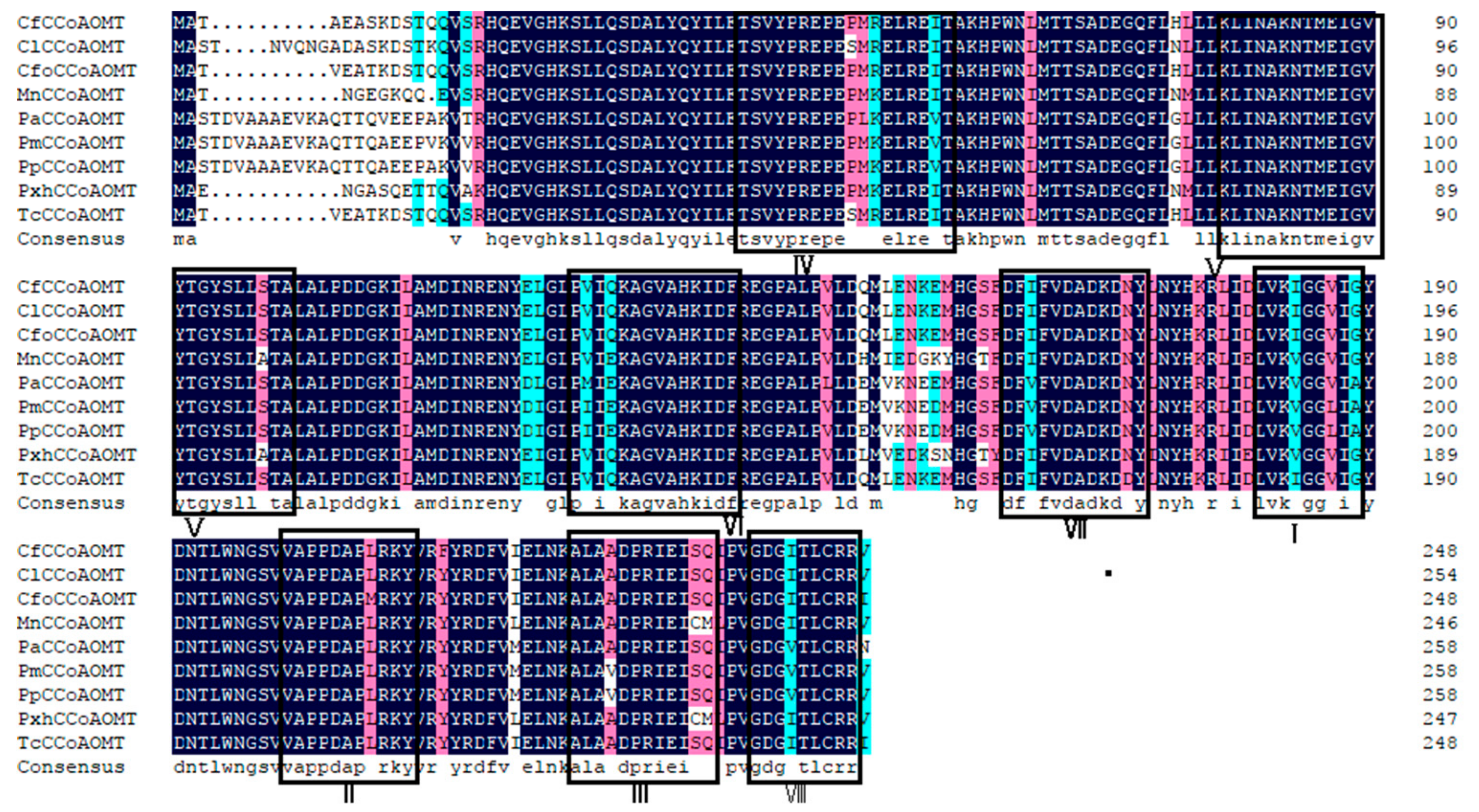
3.2. Cf4CL and CfCCoAOMT Sequences Are Conserved
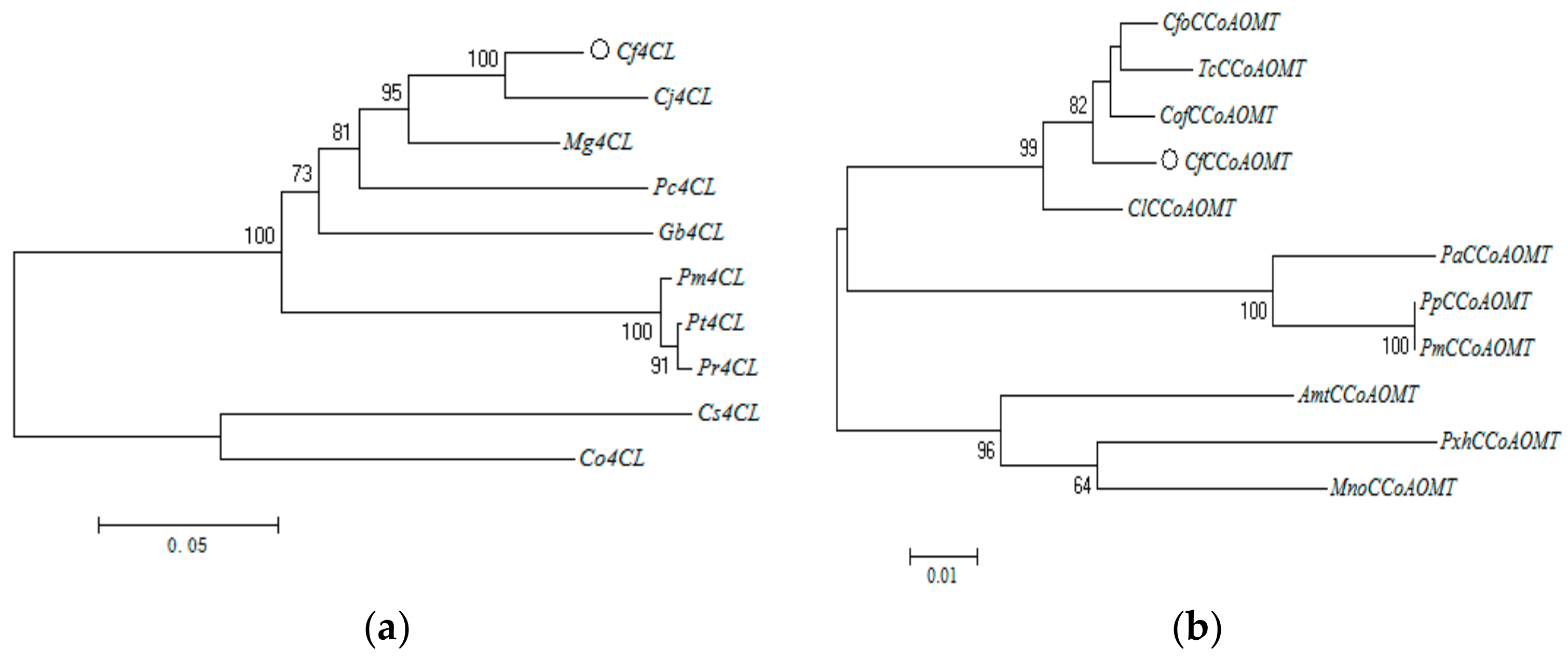
3.2.1. Protein Sequence Multiple Alignment and Phylogenetic Tree Construction
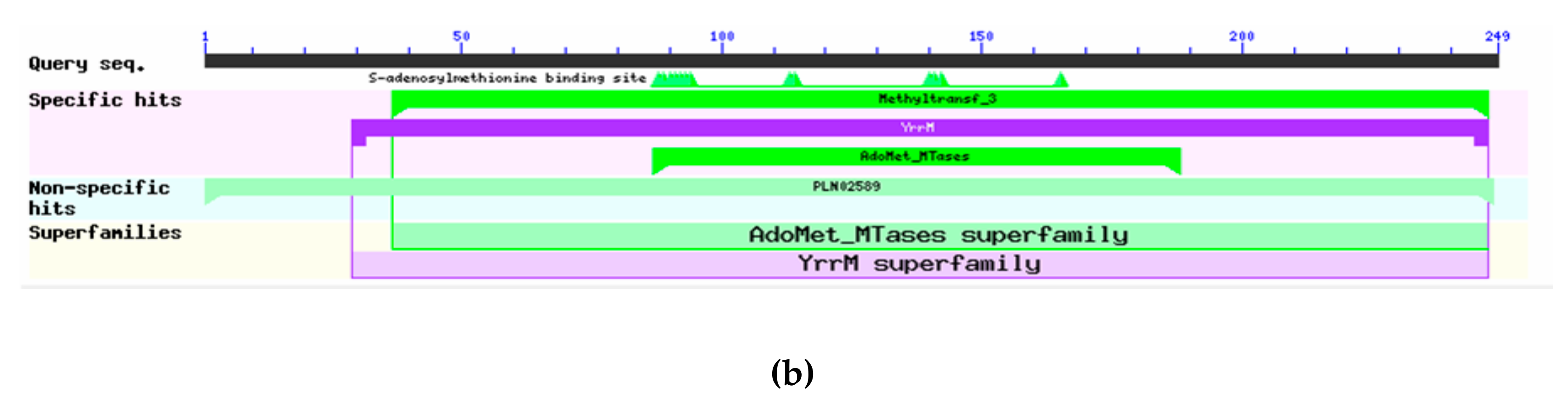
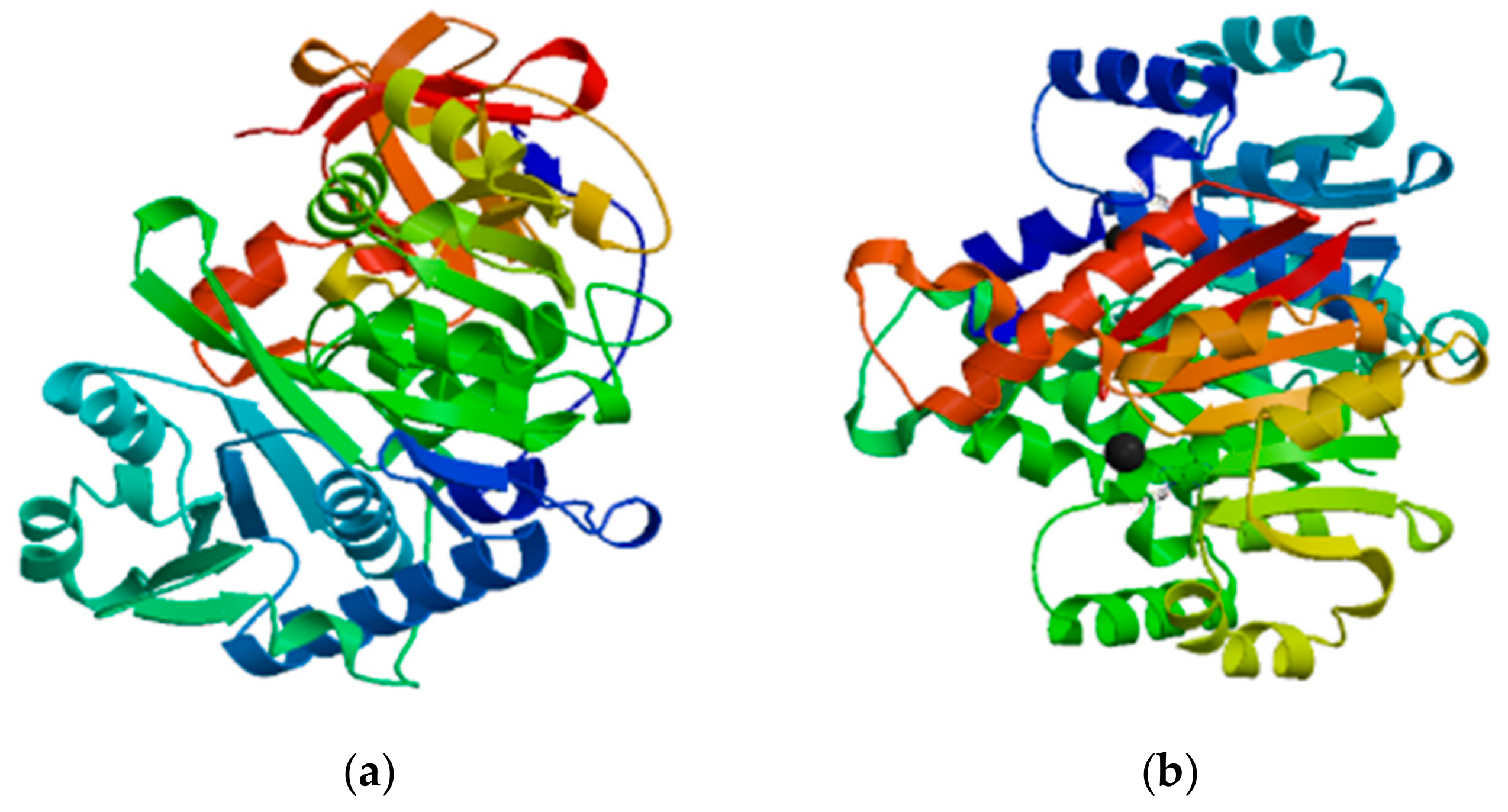
3.2.2. Cf4CL and CfCCoAOMT Protein Structure and Function Prediction
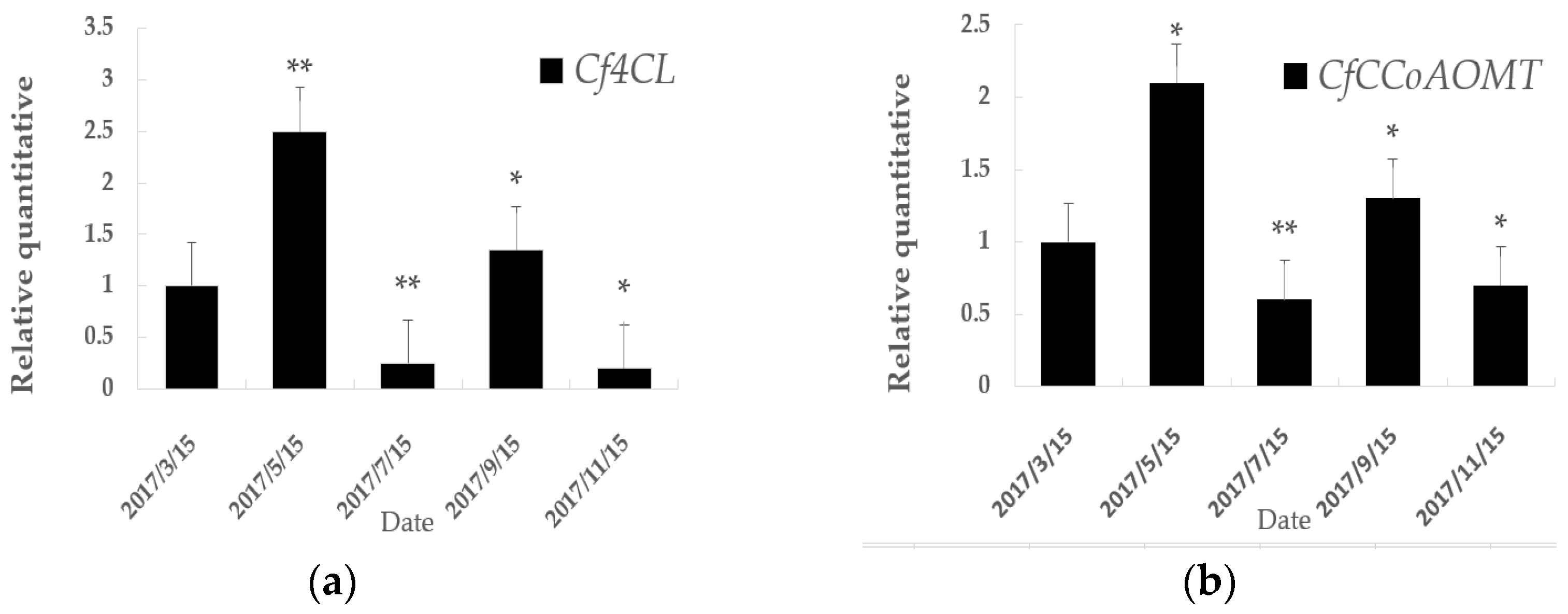
3.3. Quantitative Real-time PCR Analysis of Cf4CL and CfCCoAOMT
3.4. Functional Analysis of Cf4CL and CfCCoAOMT
3.4.1. Cf4CL and CfCCoAOMT May Increase Lignin Synthesis
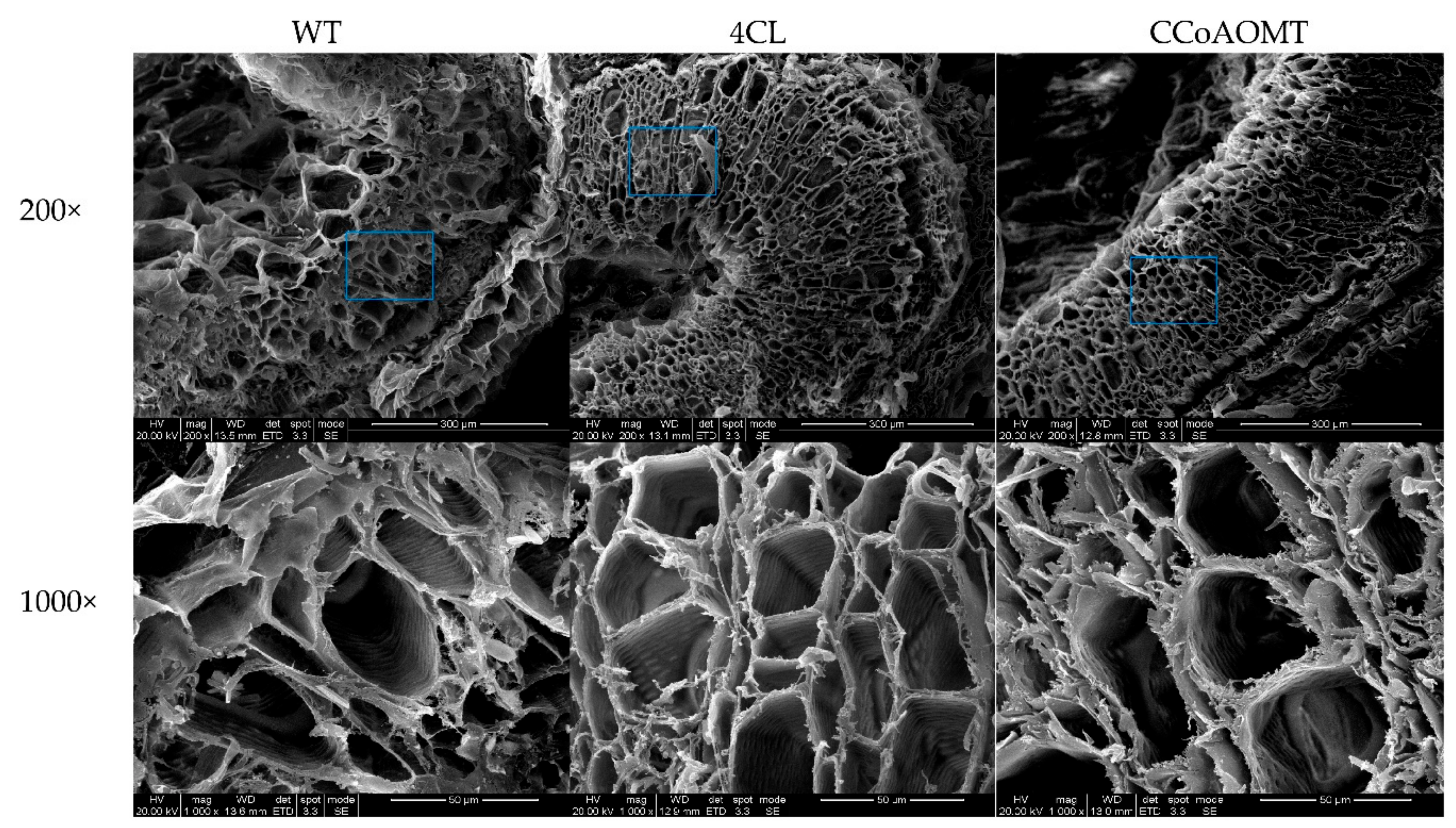
3.4.2. Cf4CL and CfCCoAOMT Expression Increases Lignin Content
4. Conclusions and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, Y.; Zou, M.; Tang, W.; Su, F.; Hu, X.; Ban, L.; Liu, W. Difference analyses of tracheid morphological characteristics among provenances of exotic loblolly pine in southern districts of Henan province in China. J. Nanjing. Univ. 2002, 26, 15–20. [Google Scholar]
- Li, L.; Cheng, X.F.; Leshkevich, J.; Umezawa, T.; Harding, S.A.; Chiang, V.L. The last step of syringyl monolignol biosynthesis in angiosperms is regulated by a novel gene encoding sinapyl alcohol dehydrogenase. Plant Cell 2001, 13, 1567–1585. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.K.; Chapple, C. The origin and evolution of lignin biosynthesis. New Phytol. 2010, 187, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, J.; Chapple, C. Rewriting the lignin roadmap. Curr. Opin. Plant Biol. 2002, 5, 224–229. [Google Scholar] [CrossRef]
- Mishima, K.; Fujiwara, T.; Iki, T.; Kuroda, K.; Yamashita, K.; Tamura, M.; Fujisawa, Y.; Watanabe, A. Transcriptome sequencing and profiling of expressed genes in cambial zone and differentiating xylem of Japanese cedar (Cryptomeria japonica). Bmc Genom. 2014, 15, 219. [Google Scholar] [CrossRef]
- Zhang, X.H.; Vincent, L.C. Molecular cloning of 4-coumarate: Coenzyme a ligase in loblolly pine and the role of this enzyme in the biosynthesis of lignin in compression wood. Plant Physiol. 1997, 113, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Douglas, C.J. Two divergent members of a tobacco 4-coumarate-coenzyme a ligase (4CL) gene family, cDNA structure, gene inheritance and expression, and peoperties if recombinant protein. Plant Physiol. 1996, 112, 193–205. [Google Scholar] [CrossRef]
- Pan, X. Study on the Structure and Function of the 4CL Gene Promoters and APX in Populus tomentosa. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2013. (In Chinese). [Google Scholar]
- Lee, D.; Meyer, K.; Chapple, C.; Douglas, C.J. Down-regulation of 4-coumarate: CoA ligase (4CL) in Arabidopsis: effect on lignin composition and implications for the control of monolignol biosynthesis. Plant Cell 1997, 9, 1985–1998. [Google Scholar]
- Lu, H.; Zhao, Y.; Jiang, X. Stable and specific expression of 4-coumarate: coenzyme A ligase gene (4CL1) driven by the xylem-specific Pto4CL1 promoter in the transgenic tobacco. Biotechnol. Lett. 2004, 26, 1147–1152. [Google Scholar] [CrossRef]
- Kajita, S.; Hishiyama, S.; Tomimura, Y. Structural characterization of modified lignin in transgenic tobacco plants in which the activity of 4-coumarate: Coenzyme A ligase is depressed. Plant Physiol. 1997, 114, 871–879. [Google Scholar] [CrossRef]
- Hua, Z.; Qing, S.; Shi, L.; Tai, W.; Yan, S. Expression analysis of Oryza sativa CCoAOMT gene. Chin. Sci. Bull. 2004, 14, 1390–1394. [Google Scholar]
- Ye, Z.H.; Kneusel, R.E.; Matern, U. An alternative methylation pathway in lignin biosynthesis in Zinnia. Plant Cell 1994, 6, 1427–1439. [Google Scholar] [PubMed]
- Li, L.G.; Osakabe, Y.K. Secondary Xylem-specific expressin of caffeoyl-coenzyme A 3-O-methyltransferase plays an important role in the methylation pathway associated with lignin biosynthetic in loblloly pine. Plant Mol. Biol. 1999, 40, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Sewalt, V.J.H.; Murray, B.G. Developmental expression and substrate specificities of Alfalfa caffeic acid 3-O-mehtyltransferase and coenzyme A-3-O-methyltransferase in relation to lignification. Plant Physiol. 1998, 117, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Meyermans, H.; Burggraeve, B.; Rycke, R.M.; Lnoue, K.; Vleesschauwer, V.; Steenackers, M.; Montagu, M.C.; Engler, G.J.; Boerjan, W.A. Cell-specific and conditional expression of caffeoyl-coenzyme A-3-O-methylytransferase in poplar. Plant Physiol. 2000, 123, 853–867. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. Transcriptome Sequencing and Profiling of Expression in Vascular Cambium of Cryptomeria Fortune. Master’s Thesis, Nanjing Forestry University, Nanjing, China, 2016. [Google Scholar]
- Jokipii-Lukkari, S.; Delhomme, N.; Schiffthaler, B.; Mannapperuma, C.; Prestele, J.; Nilsson, O.; Street, N.; Tuominen, H. Transcriptional roadmap to seasonal variation in wood formation of Norway spruce. Plant Physiol. 2018, 176, 2851–2870. [Google Scholar] [CrossRef] [PubMed]
- Curtis, I.; Davey, M.; Power, J. Leaf disk transformation. Meth. Mol. Biol. 1995, 44, 59–70. [Google Scholar]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.S.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Geourjon, C.; Deleage, G. SOPMA: significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, M.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Petersen, T.; Brunak, S.; Heijne, G.; Nielsen, H. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, W. OLIGO 7 Primer Analysis Software. In Methods in Molecular Biology Vol. 402: PCR Primer Design; Yuryev, A., Ed.; Humana Press: Totowa, NJ, USA, 2007; pp. 35–59. [Google Scholar]
- Liu, S.; Li, A.; Chen, C.; Xu, M.; Cai, G.; Zhang, L.; Guo, C. De novo transcriptome sequencing in Passiflora edulis sims to identify genes and signaling pathways involved in cold tolerance. Forests 2017, 8, 435. [Google Scholar] [CrossRef]
- Little, B.; Wagner, P.; Ray, R.; Pope, R.; Scheetz, R. Biofilms—An ESEM evaluation of artifacts introduced during SEM preparation. J. Ind. Microbiol. Biotechnol. 1991, 8, 213–221. [Google Scholar] [CrossRef]
- Rodrigues, J.; Faix, O.; Pereira, H. Improvement of the acetylbromide method for lignin determination within large scale screening programmes. Eur. J. Wood Wood Prod. 1999, 57, 341–345. [Google Scholar] [CrossRef]
- Li, J.; Fan, F.; Wang, L.; Zhan, Q.; Wu, P.; Du, J.; Yang, X.; Liu, Y. Cloning and expression analysis of cinnamoyl-CoA reductase (CCR) genes in sorghum. PeerJ 2016, 4, e2005. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wei, L.; Li, H.; Li, J. The analysis of genes and phytohormone metabolic pathways associated with leaf shape development in Liriodendron chinense via de novo transcriptome sequencing. Genes 2018, 9, 577. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, Y.; Fu, C.; Zhou, G.; Gao, H. Progress of a key enzyme-caffeoyl-co a 3-o-methyltransferase in lignin biosynthesis. Genom. Appl. Biol. 2014, 33, 458–466. [Google Scholar]
- Challis, G.L.; Ravel, J.; Townsend, C.A. Predictive, structure-based model of amino acid recognition by nonribosomal peptide synthetase adenylation domains. Chem. Biol. 2000, 7, 211–224. [Google Scholar] [CrossRef]
- Stuible, H.P.; Buttner, D.; Ehlting, J.; Hahlbrok, K.; Kombrink, E. Mutation alanalysis of 4-coumarate: CoA ligase identifies functionally important amino acids andverifies its close relationship to othe radenylate-form ingenzymes. Febs Lett. 2000, 467, 117–122. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/protein/ (accessed on 1 December 2006).
- Cronn, R.; Dolan, P.C.; Jogdeo, S.; Wegrzyn, J.; Neale, D.; Bradley, J.; Denver, D. Transcription through the eye of a needle: Daily and annual cyclic gene expression variation in Douglas-fir needles. Bmc Genom. 2017, 18, 558. [Google Scholar] [CrossRef]
- Cukovic, D.; Ehlting, J.; Vanziffle, J.A.; Douglas, C.J. Structureand evolution of 4-coumarate: Coenzyme A ligase (4CL) gene families. Biol. Chem. 2001, 382, 645–654. [Google Scholar] [PubMed]
- Huang, J.; Li, W.; Qu, C.; Liu, L.; Xu, X.; Wang, R.; Li, J. Expression characteristics of key genes in Lignin pathway among different lodging resistance lines of Brassica napus L. Acta Agron. Sin. 2013, 39, 1339–1344. [Google Scholar] [CrossRef]
- Pagadala, N.S.; Arha, M.; Reddy, P.; Kumar, R.; Sirisha, V.L.; Prashant, S.; Reddy, K.; Khan, B.; Rawal, S.K.; Kishor, P.B. Phylogenetic analysis, homology modelling, molecular dynamics and docking studies of caffeoyl-coa-o-methyl transferase (CCoAOMT1 and 2) isoforms isolated from subabul (Leucaena leucocephala). J. Mol. Model. 2009, 15, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Cheng, T.; Chen, J.; Lu, M.; Liu, G.; Li, M.; Shi, J.; Lu, Y.; Laux, T. Expansion and functional divergence of AP2 group genes in Spermatophytes determined by molecular evolution and Arabidopsis mutant analysis. Front. Plant Sci. 2016, 7, 1383. [Google Scholar] [CrossRef]
- Zhao, H.; Wei, J.; Lu, J.; Song, Y.; Wang, T.; Zhu, Z. cDNA cloning and functional analysis of 4-coumarate: CoA ligase (4CL) gene in Chinese white aspen. Prog. Nat. Sci. 2003, 12, 895–900. [Google Scholar] [CrossRef]
- Meng, H.; Campbell, W. Substrate profiles and expression of caffeoyl coenzyme A and caffeic acid O-methyltransferases in secondary xylem of aspen during seasonal development. Plant Mol. Biol. 1998, 38, 513–520. [Google Scholar] [CrossRef]
- Oribe, Y.; Funada, R.; Shibagaki, M.; Kubo, T. Cambial reactivation in locally heated stems of the evergreen conifer Abies sachalinensis (Schmidt) masters. Planta 2001, 212, 684–691. [Google Scholar] [CrossRef]
- O’Connell, A.; Holt, K.; Piquemal, J.; Grima, J.; Boudet, A.; Pollet, B.; Lapierre, C.; Petit, M.; Schuch, W.; Halpin, C. Improved paper pulp from plants with suppressed cinnamoyl-CoA reductase or cinnamyl alcohol dehydrogenase. Transgenic Res. 2002, 11, 495–503. [Google Scholar] [CrossRef]
- Lapierre, C.; Pollet, B.; Petit-Conil, M. Structural alteration of lignin in transgenic poplars with depressed cinnamyl alcohol dehydrogenase or caffeic acid O-methyl-transferase activity have an opposite impact on the efficiency of industrial kraft pulping. Plant Physiol 1999, 119, 153–163. [Google Scholar] [CrossRef]
- Song, F. Expression and Functional Analysis of Correlative Enzyme Genes of Lignin Phenylpropanoid Biosynthetic Pathway in Betula platphylla. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2009. (In Chinese). [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Application |
|---|---|---|
| Longer | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | Universal primer |
| Shorter | CTAATACGACTCACTATAGGGC | |
| 4CL-5′outer | GGCATTCGCTCTCATCCGCCTCAGTCAG | 4CL RACE reaction |
| 4CL-5′inner | GCTCCACCGGACATGATCAAGC | |
| 4CL-3′outer | ATAGCCCACCTGAAGGTTGCCTCCCGATT | |
| 4CL-3′inner | CTATTCCCTCAATTCGGTGCTC | |
| CCoAOMT-5′outer | GTATTGTCATAGCCGATCACTCCCCCAA | CCoAOMT RACE reaction |
| CCoAOMT-5′inner | CAATTTTGTGGGCAACACCTG | |
| CCoAOMT-3′outer | GCCCTGCCTGATGATGGAAAGATCCTAG | |
| CCoAOMT-3′inner | GATCGGCTATGACAATACTCTG | |
| 4CL-ALL-F | GGGTAAAGATCAATTACTGCTTC | Amplification of the conserved region |
| 4CL-ALL-R | CAAATTTATGTGTGCTGCGAGT | |
| CCoAOMT-ALL-F | GTTTCATTCCTCCAATCCAGT | |
| CCoAOMT-ALL-R | GGGCTGTTCTTAAATCACTCC | |
| RT-4CL-F | CTCCTTTGTGCGCTCCGAGT | RT-PCR |
| RT-4CL-R | GCTCCACCGGACATGATCAAGC | |
| RT-CCoAOMT-F | GCTCATCAATGCCAAGAACACCA | |
| RT-CCoAOMT-R | TCAATTTTGTGGGCAACACCT | |
| CE-4CL-F | ATGGCTTGTGTCGCACCTCT | Enzyme cutting |
| CE-4CL-R | TTAGGCTGCTGCAAGTCTGGC | |
| CE-CCoAOMT-F | ATGGCAACTGCAGAGGCTTC | |
| CE-CCoAOMT-R | AATAACTCTTCTGCAGAGAGTGATGC |
| Amino Acids/aa | Molecular Weight/kD | PI Value | Molecular Formula | Fat Coefficient | Hydrophilicity | Unstable Coefficient | |
|---|---|---|---|---|---|---|---|
| Cf4CL | 554 | 59.86 | 5.41 | C2700H4309N699O793S19 | 103.84 | 0.124 | 36.15 |
| CfCCoAOMT | 249 | 28.098 | 5.52 | C1261H1994N338O372S8 | 99.88 | −0.228 | 39.40 |
| Plant | WT | 4CL | CCoAOMT | ||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | ||
| Average lignin content % | 4.86 | 8.31 | 10.51 | 7.14 | 9.13 | 7.54 | 8.34 |
| SD | 0.983 | 1.457 | 2.819 | 0.404 | 2.318 | 0.507 | 1.365 |
| Sig | <0.05 | <0.01 | <0.05 | <0.01 | <0.05 | <0.01 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Hua, H.; Xu, J.; Mo, J.; Zhao, H.; Yang, J. Cloning and Functional Analysis of Lignin Biosynthesis Genes Cf4CL and CfCCoAOMT in Cryptomeria fortunei. Genes 2019, 10, 619. https://doi.org/10.3390/genes10080619
Guo Z, Hua H, Xu J, Mo J, Zhao H, Yang J. Cloning and Functional Analysis of Lignin Biosynthesis Genes Cf4CL and CfCCoAOMT in Cryptomeria fortunei. Genes. 2019; 10(8):619. https://doi.org/10.3390/genes10080619
Chicago/Turabian StyleGuo, Zhenhao, Hui Hua, Jin Xu, Jiaxing Mo, Hui Zhao, and Junjie Yang. 2019. "Cloning and Functional Analysis of Lignin Biosynthesis Genes Cf4CL and CfCCoAOMT in Cryptomeria fortunei" Genes 10, no. 8: 619. https://doi.org/10.3390/genes10080619
APA StyleGuo, Z., Hua, H., Xu, J., Mo, J., Zhao, H., & Yang, J. (2019). Cloning and Functional Analysis of Lignin Biosynthesis Genes Cf4CL and CfCCoAOMT in Cryptomeria fortunei. Genes, 10(8), 619. https://doi.org/10.3390/genes10080619