Contrasting Asymptomatic and Drug Resistance Gene Prevalence of Plasmodium falciparum in Ghana: Implications on Seasonal Malaria Chemoprevention

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
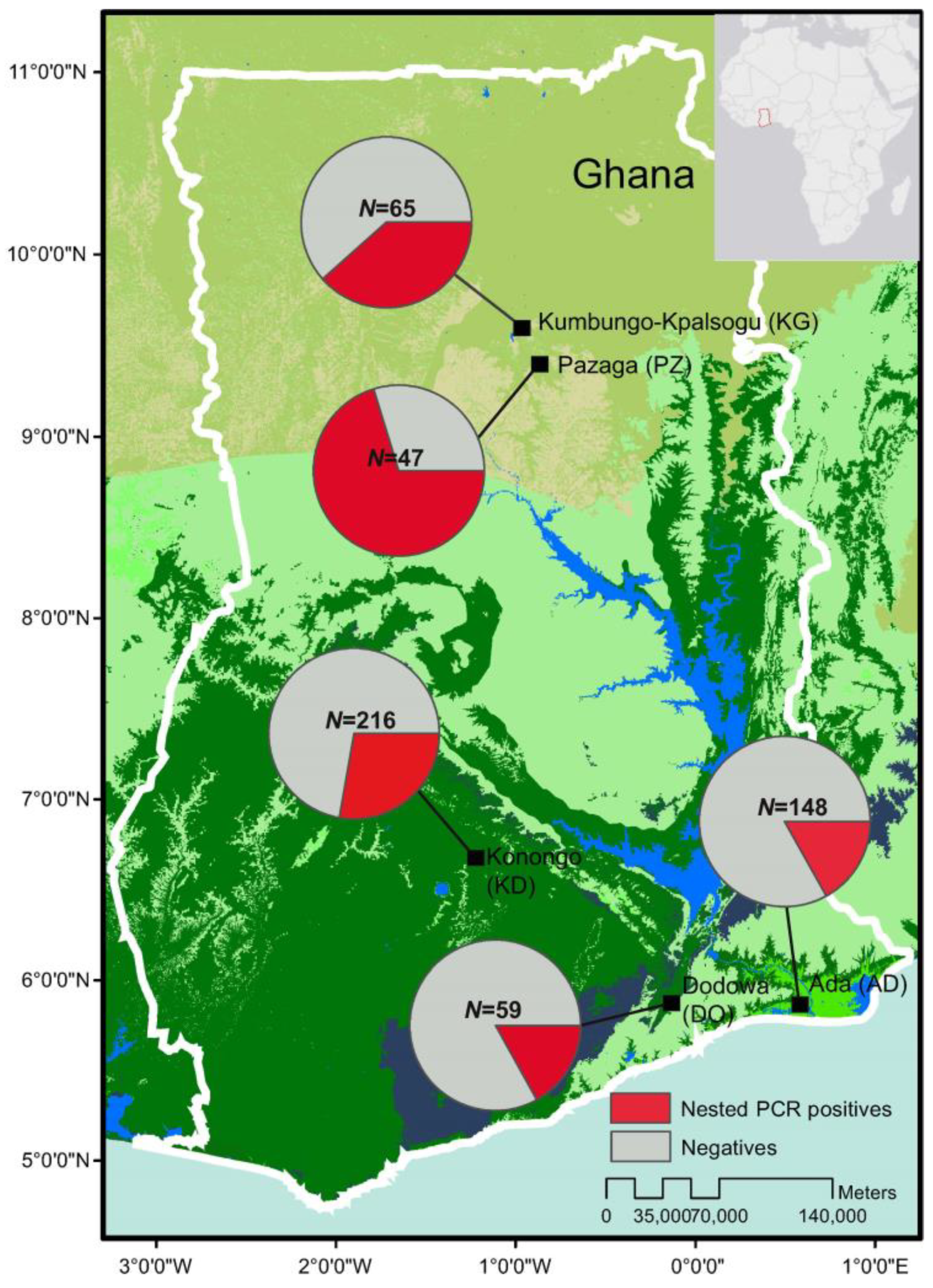
2.2. Sampling and Study Sites
2.3. Microscopic and Molecular Screening
2.4. Assessing Sensitivity and Specificity
2.5. Resistance gene Mutation Frequency
3. Results
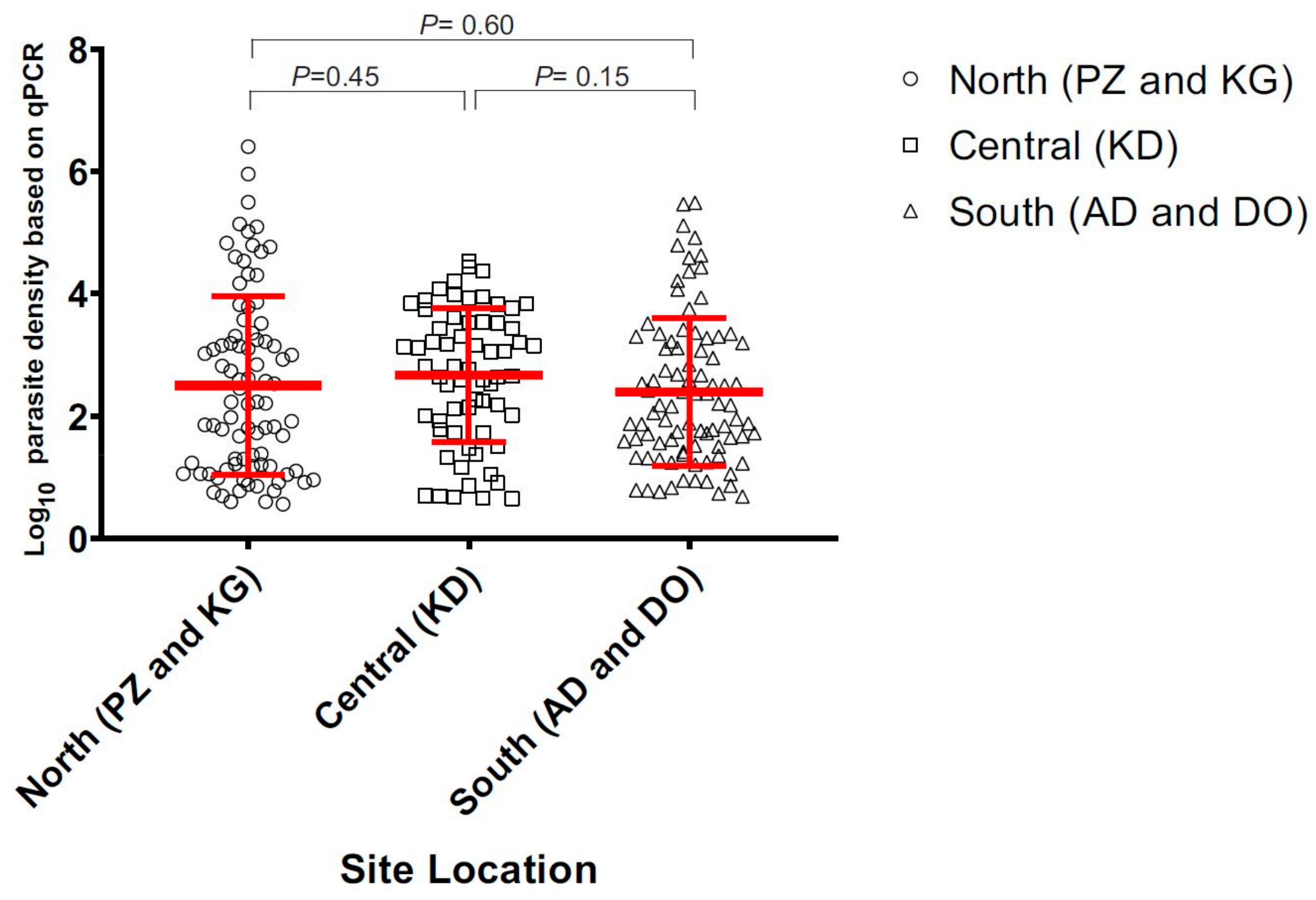
3.1. Asymptomic Prevalence across Ecological Zones
3.1.1. Sensitivity and Specificity of Diagnostic Methods
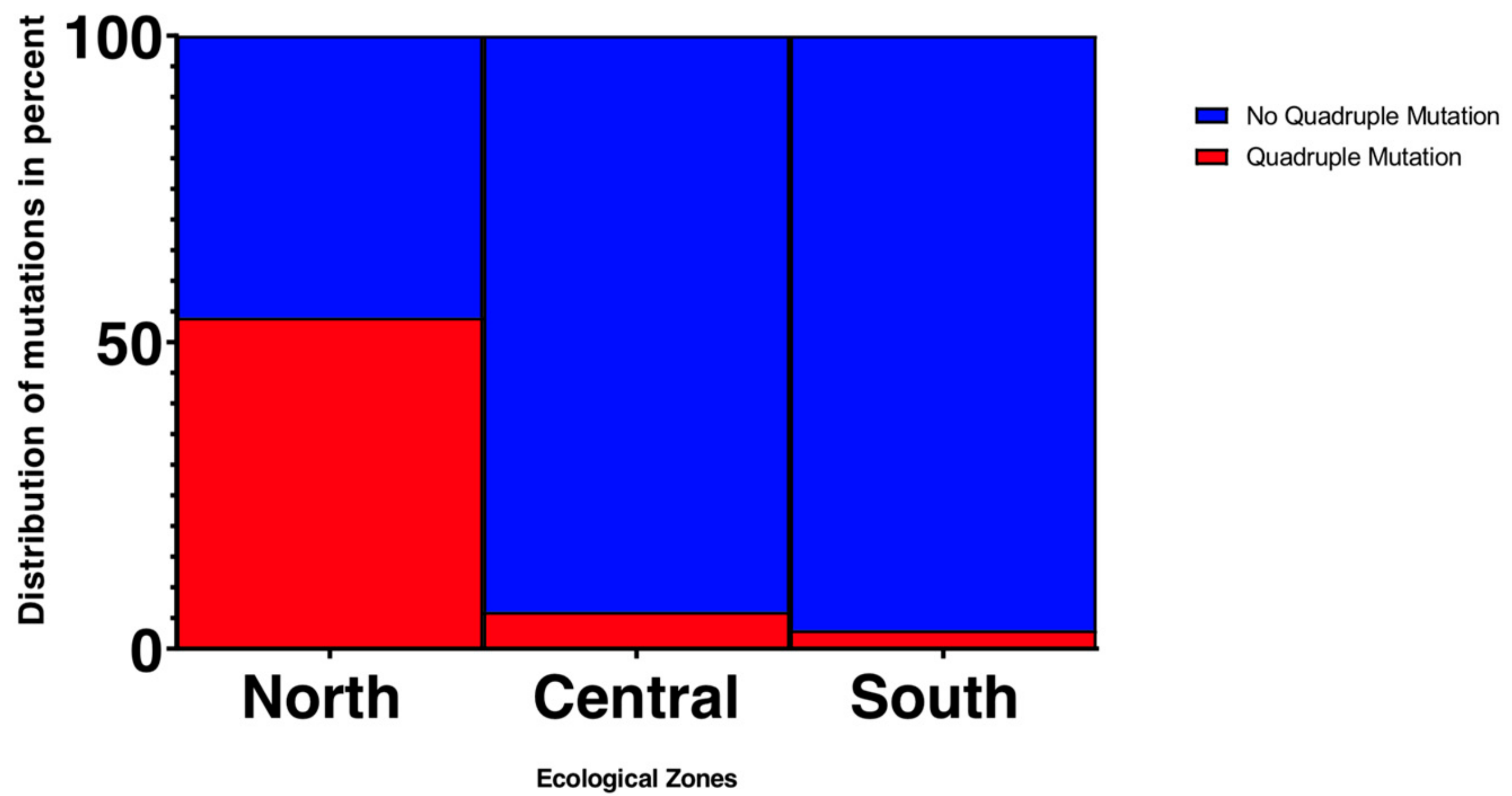
3.1.2. Frequency of Resistance Gene Mutations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Malaria Report 2019. Available online: https://www.cdc.gov/parasites/malaria/index.html (accessed on 25 April 2019).
- Snow, R.W.; Omumbo, J.A. Malaria. In Disease and Mortality in Sub-Saharan Africa, 2nd ed.; Jamison, D.T., Feachem, R.G., Makgoba, M.W., Bos, E.R., Baingana, F.K., Hofman, K.J., Rogo, K.O., Eds.; The International Bank for Reconstruction and Development: Washington, DC, USA, 2006; pp. 195–213. ISBN 978-0-8213-6397-3. [Google Scholar]
- Malaria Operational Plan FY 2018. President’s Malaria Initiative: Ghana. Available online: https://reliefweb.int/report/ghana/president-s-malaria-initiative-ghana-malaria-operational-plan-fy-2018/ (accessed on 6 June 2019).
- Unicef. World Malaria Day 2018. Available online: https://www.unicef.org/ghana/media_12060.html (accessed on 25 April 2019).
- Conway, D.J.; Roper, C.; Oduola, A.M.J.; Arnot, D.E.; Kremsner, P.G.; Grobusch, M.P.; Curtis, C.F.; Greenwood, B.M. High recombination rate in natural populations of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1999, 96, 4506–4511. [Google Scholar] [CrossRef] [PubMed]
- Ayanful-Torgby, R.; Quashie, N.B.; Boampong, J.N.; Williamson, K.C.; Amoah, L.E. Seasonal variations in Plasmodium falciparum parasite prevalence assessed by varying diagnostic tests in asymptomatic children in southern Ghana. PLoS ONE 2018. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; von Seidlein, L.; Nguyen, T.V.; Truong, P.N.; Hung, S.D.; Pham, H.T.; Nguyen, T.U.; Le, T.D.; Dao, V.H.; Mukaka, M.; et al. The persistence and oscillations of submicroscopic Plasmodium falciparum and Plasmodium vivax infections over time in Vietnam: An open cohort study. Lancet Infect. Dis. 2018, 18, 565–572. [Google Scholar] [CrossRef]
- Wangai, L.N.; Karau, M.G.; Njiruh, P.N.; Sabah, O.; Kimani, F.T.; Magoma, G.; Kiambo, N. Sensitivity of Microscopy Compared to Molecular Diagnosis of P. Falciparum: Implications on Malaria Treatment in Epidemic Areas in Kenya. Afr. J. Infect. Dis. 2011, 5, 1–6. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Andrews, L.; Andersen, R.F.; Webster, D.; Dunachie, S.; Walther, R.M.; Bejon, P.; Hunt-Cooke, A.; Bergson, G.; Sanderson, F.; Hill, A.V.; et al. A Quantitative real-time polymerase chain reaction for malaria diagnosis and its use in malaria vaccine clinical trials. Am. J. Trop. Med. Hyg. 2005, 73, 191–198. [Google Scholar] [CrossRef]
- Hofmann, N.; Mwingira, F.; Shekalaghe, S.; Robinson, L.J.; Mueller, I.; Felger, I. Ultra-sensitive detection of Plasmodium falciparum by amplification of multi-copy subtelomeric targets. PLoS Med. 2015, 12. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.M.; Pirrit, L.A.; Scherf, A.; Pirritt, L.A. Genomic organization and chromatin structure of Plasmodium falciparum chromosome ends. Mol. Biochem. Parasitol. 2000, 106, 169–174. [Google Scholar] [CrossRef]
- Hou, Y.; Zhang, H.; Miranda, L.; Lin, S. Serious overestimation in quantitative PCR by circular (supercoiled) plasmid standard: Microalgal pcna as the model gene. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Sidhu, A.B.; Verdier-Pinard, D.; Fidock, D.A. Chloroquine resistance in Plasmodium falciparum malaria parasites conferred by pfcrt mutations. Science 2002, 298, 210–213. [Google Scholar] [CrossRef]
- World Health Organization—Malaria Country Profile: Ghana. Available online: https://www.who.int/malaria/publications/country-profiles/profile_gha_en.pdf?ua=1 (accessed on 7 June 2019).
- World Health Organization. WHO Policy Recommendation: Seasonal Malaria Chemoprevention for Plasmodium falciparum Control in Highly Seasonal Transmission Areas of the Sahel Sub-Region in Africa. Available online: https://www.who.int/malaria/publications/atoz/who_smc_policy_recommendation/en/ (accessed on 25 April 2019).
- Druetz, T.; Corneau-Tremblay, N.; Millogo, T.; Kouanda, S.; Ly, A.; Bicaba, A.; Haddad, S. Impact Evaluation of Seasonal Malaria Chemoprevention under Routine Program Implementation: A Quasi-Experimental Study in Burkina Faso. Am. J. Trop. Med. Hyg. 2018, 98, 524–533. [Google Scholar] [CrossRef]
- Seasonal Malaria Chemoprevention (SMC). Available online: http://www.ghanahealthservice.org/malaria/subcategory.php?nmcpscid=115&nmcpcid=85 (accessed on 25 April 2019).
- Kupferschmidt, H.G.; Schröder, K.; Beltzner, B. Chloroquine and fansidar resistance of Plasmodium falciparum now also in Ghana. Angew. Parasitol. 1988, 29, 211–213. [Google Scholar] [PubMed]
- Fidock, D.A.; Nomura, T.; Talley, A.K.; Cooper, R.A.; Dzekunov, S.M.; Ferdig, M.T.; Ursos, L.M.; Sidhu, A.B.; Naudé, B.; Deitsch, K.W.; et al. Mutations in the P. falciparum digestive vacuole transmembrane protein PfCRT and evidence for their role in chloroquine resistance. Mol. Cell. 2000, 6, 861–871. [Google Scholar] [CrossRef]
- Gatton, M.L.; Martin, L.B.; Cheng, Q. Evolution of Resistance to Sulfadoxine-Pyrimethamine in Plasmodium falciparum. Antimicrob. Agents Chemother. 2004, 48, 2116–2123. [Google Scholar] [CrossRef] [PubMed]
- Juma, D.W.; Omondi, A.A.; Ingasia, L.; Opot, B.; Cheruiyot, A.; Yeda, R.; Okudo, C.; Cheruiyot, J.; Muiruri, P.; Ngalah, B.; et al. Trends in drug resistance codons in Plasmodium falciparum dihydrofolate reductase and dihydropteroate synthase genes in Kenyan parasites from 2008 to 2012. Malar. J. 2014, 13, 250. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, D.; Dieye, B.; Ndiaye, Y.D.; Van Tyne, D.; Daniels, R.; Bei, A.K.; Mbaye, A.; Valim, C.; Lukens, A.; Mboup, S.; et al. Polymorphism in dhfr/dhps genes, parasite density and ex vivo response to pyrimethamine in Plasmodium falciparum malaria parasites in Thies, Senegal. Int. J. Parasitol. Drugs Drug Resist. 2013, 3, 135–142. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miguel, R.B.; Coura, J.R.; Samudio, F.; Suárez-Mutis, M.C. Evaluation of three different DNA extraction methods from blood samples collected in dried filter paper in Plasmodium subpatent infections from the Amazon region in Brazil. Rev. Inst. Med. Trop. Sao Paulo 2013, 55, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Afoakwah, R.; Boampong, J.N.; Egyir-Yawson, A.; Nwaefuna, E.K.; Verner, O.N.; Asare, K.K. High prevalence of PfCRT K76T mutation in Plasmodium falciparum isolates in Ghana. Acta Trop. 2014, 136, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Duraisingh, M.T.; Curtis, J.; Warhurst, D. Plasmodium falciparum: Detection of polymorphisms in the Dihydrofolate reductase and Dihydropteroate synthetase genes by PCR and restriction digestion. Exp. Parasitol. 1998, 89, 1–8. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Talundzic, E.; Ravishankar, S.; Kelly, J.; Patel, D.; Plucinski, M.; Schmedes, S.; Ljolje, D.; Clemons, B.; Madison-Antenucci, S.; Arguin, P.M.; et al. A next-generation sequencing and bioinformaticsprotocol for Malaria drug Resistance marker Surveillance (MaRS). Am. Soc. Microbiol. 2018. [Google Scholar] [CrossRef]
- Lo, E.; Hemming-Schroeder, E.; Yewhalaw, D.; Nguyen, J.; Kebede, E.; Zemene, E.; Getachew, S.; Tushune, K.; Zhong, D.; Zhou, G.; et al. Transmission dynamics of co-endemic Plasmodium vivax and P. falciparum in Ethiopia and prevalence of antimalarial resistant genotypes. PLoS Negl. Trop. Dis. 2017, 11, e0005806. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Omosun, Y.; Lal, A.; Odero, C.; Gatei, W.; Otieno, K.; Shi, Y.P. Assessment of molecular markers for anti-malarial drug resistance after the introduction and scale-up of malaria control interventions in western Kenya. Malar. J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Mlambo, G.; Sullivan, D.; Mutambu, S.L.; SokoJoel, W.; Mbedzi, J.; Chivenga, J.; Gemperli, A.; Kuma, N. High prevalence of molecular markers for resistance to chloroquine and pyrimethamine in Plasmodium falciparum from Zimbabwe. Parasitol. Res. 2007, 101, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Microscopic Tests. Available online: https://www.malariasite.com/microscopic-tests/ (accessed on 25 April 2019).
- Joanny, F.; Löhr, S.J.; Engleitner, T.; Lell, B.; Mordmüller, B. Limit of blank and limit of detection of Plasmodium falciparum thick blood smear microscopy in a routine setting in Central Africa. Malar. J. 2014, 13, 234. [Google Scholar] [CrossRef] [PubMed]
- Rougemont, M.; Van Saanen, M.; Sahli, R.; Hinrikson, H.P.; Bille, J.; Jaton, K. Detection of four Plasmodium species in blood from humans by 18S rRNA gene subunit-based and species-specific real-time PCR assays. J. Clin. Microbiol. 2004, 42, 5636–5643. [Google Scholar] [CrossRef] [PubMed]
- Ohrt, C.; Purnom; Sutamihardja, M.A.; Tang, D.; Kain, K.C. Impact of microscopy error on estimates of protective efficacy in malaria-prevention trials. J. Infect. Dis. 2002, 186, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Dery, D.B.; Brown, C.; Asante, K.P.; Adams, M.; Dosoo, D.; Amenga-Etego, S.; Wilson, M.; Chandramohan, D.; Greenwood, B.; Owusu-Agyei, S. Patterns and Seasonality of Malaria Transmission in the Forest-Savannah Transitional Zones of Ghana. Malar. J. BioMed Central 2010. [Google Scholar] [CrossRef]
- Appawu, M.A.; Owusu-Agyei, S.; Dadzie, S.; Asoala, V.; Anto, F.; Koram, K.; Rogers, W.; Nkrumah, F.; Hoffman, S.L.; Fryauff, D.J. Malaria transmission dynamics at a site in northern Ghana proposed for testing of malaria vaccines. Trop. Med. Int. Health 2004, 9, 164–170. [Google Scholar] [CrossRef]
- Appawu, M.A.; Baffoe-Wilmot, A.; Afari, E.A.; Dunyo, S.; Koram, K.A.; Nkrumah, F.K. Malaria vector studies in two ecological zones in southern Ghana. Afr. Entomol. 2001, 9, 59–65. [Google Scholar]
- Russell, S. International Migration. In Demographic Change in Sub-Saharan Africa; Martin, L.G., Foote, K.A., Eds.; National Research Council: Washington, DC, USA, 1993; ISBN 978-0-309-04942-9. [Google Scholar]
- Tutu, K.A. Determinants of Internal Migration. In Migration Research Study in Ghana; Twum-Baah, K.A., Nabila, J.S., Aryee, A.F., Eds.; Ghana Statistical Survey: Accra, Ghana, 1995. [Google Scholar]
- Gatton, M.L.; Cheng, Q. Modeling the development of acquired clinical immunity to Plasmodium falciparum malaria. Infect Immun. 2004, 72, 6538–6545. [Google Scholar] [CrossRef]
- Schousboe, M.L.; Ranjitkar, S.; Rajakaruna, R.S.; Amerasinghe, P.H.; Morales, F.; Pearce, R.; Ord, R.; Leslie, T.; Rowland, M.; Gadalla, N.B.; et al. Multiple Origins of Mutations in the mdr1 Gene—A Putative Marker of Chloroquine Resistance in P. vivax. PLoS Negl. Trop. Dis. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Eijk, A.M.; Blokland, I.E.; Slutsker, L.; Odhiambo, F.; Ayisi, J.G.; Bles, H.M.; Rosen, D.H.; Adazu, K.; Lindblade, K.A. Use of intermittent preventive treatment for malaria in pregnancy in a rural area of western Kenya with high coverage of insecticide-treated bed nets. Trop. Med. Int. Health 2005, 10, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Dellicour, S.; Hill, J.; Bruce, J.; Ouma, P.; Marwanga, D.; Otieno, P.; Desai, M.; Hamel, M.J.; Kariuki, S.; Webster, J. Effectiveness of the delivery of interventions to prevent malaria in pregnancy in Kenya. Malar. J. 2016. [Google Scholar] [CrossRef] [PubMed]
- Duah, N.O.; Quashie, N.B.; Abuaku, B.K.; Sebeny, P.J.; Kronmann, K.C.; Koram, K.A. Surveillance of molecular markers of Plasmodium falciparum resistance to sulphadoxine-pyrimethamine 5 years after the change of malaria treatment policy in Ghana. Am. J. Trop. Med. Hyg. 2012, 87, 996–1003. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Method | Positive | Negative | Sensitivity (95% CI) | Specificity (95% CI) |
|---|---|---|---|---|
| Microscopy * | 35 (18.5%) | 100 (81.5%) | - | - |
| 18s Nested PCR | 62 (45.9%) | 73 (54.1%) | 63.8% (53.3–73.5%) | 91.3% (71.9–98.9%) |
| 18s qPCR | 76 (56.3%) | 59 (43.7%) | 80.9% (71.4–88.2%) | 75.5% (60.4–87.1%) |
| TARE-2 qPCR | 89 (65.9%) | 46 (34.1%) | 81.2% ((74.4–86.8%) | 48.3% (30.1–66.9%) |
| N | Gene | Codon | Total (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Region | |||||||||
| North | Central | South | |||||||
| KG | PZ | KD | AD | DO | |||||
| Pfcrt | |||||||||
| 165 | K76T | K | 38 (97) | 36 (100) | 45 (98) | 26 (96) | 16 (94) | 161 (99) | |
| T | 1 (3) | 0 | 1 (2) | 1 (4) | 1 (6) | 4 (1) | |||
| Pfmdr1 | |||||||||
| 216 | N86Y | N | 55 (100) | 61 (94) | 45 (100) | 29 (88) | 17 (94) | 207 (96) | |
| Y | 0 | 4 (6) | 0 | 4 (12) | 1 (6) | 9 (4) | |||
| 183 | Y184F | Y | 40 (74) | 32 (64) | 13 (28) | 6 (43) | 10 (56) | 101 (55) | |
| F | 14 (26) | 18 (36) | 34 (72) | 8 (57) | 8 (44) | 82 (45) | |||
| Pfdhps | |||||||||
| 109 | A437G | A | 3 (9) | 0 | 0 | 0 | 0 | 3 (3) | |
| G | 30 (91) | 33 (100) | 45 (100) | 24 (100) | 13 (100) | 106 (97) | |||
| 155 | K540E | K | 32 (100) | 40 (98) | 45 (100) | 24 (100) | 13 (100) | 154 (99) | |
| E | 0 | 1 (2) | 0 | 0 | 0 | 1 (1) | |||
| Pfdhfr | |||||||||
| 189 | N51I | N | 6 (12) | 4 (8) | 6 (12) | 4 (11) | 0 | 20 (11) | |
| I | 43 (88) | 47 (92) | 43 (88) | 33 (89) | 0 | 169 (89) | |||
| 209 | C59R | C | 13 (28) | 12 (23) | 4 (8) | 0 | 0 | 29 (14) | |
| R | 34 (72) | 41 (77) | 47 (92) | 35 (100) | 23 (100) | 180 (86) | |||
| 205 | S108N | S | 2 (4) | 0 | 3 (6) | 0 | 0 | 5 (2) | |
| N | 45 (94) | 52 (100) | 49 (94) | 35 (100) | 23 (100) | 204 (98) | |||
| Gene | Codon | ||||||
|---|---|---|---|---|---|---|---|
| West Africa | East Africa | Southern Africa | |||||
| Ghana | Gambia | Ethiopia | Kenya | Zimbabwe | |||
| Pfcrt | |||||||
| K76T | K | 161 (99) | 138 (57) | 116 (39) | 69 (27) | 22 (20) | |
| T | 4 (1) | 105 (43) | 189 (61) | 185 (73) | 90 (80) | ||
| Pfmdr1 | |||||||
| N86Y | N | 207 (96) | 202 (83) | 142 (93) | 78 (31) | 37 (33) | |
| Y | 9 (4) | 43 (17) | 10 (7) | 174 (69) | 75 (67) | ||
| Y184F | Y | 101 (55) | 88 (36) | 18 (10) | 195 (77) | - | |
| F | 82 (45) | 157 (64) | 181 (90) | 58 (23) | - | ||
| Pfdhps | |||||||
| A437G | A | 3 (3) | 180 (74) | 152 (85) | 3 (1) | 98 (87) | |
| G | 106 (97) | 63 (26) | 27 (15) | 250 (99) | 14 (13) | ||
| K540E | K | 154 (99) | 23 (10) | - | 0 (0) | 96 (85) | |
| E | 1 (1) | 222 (90) | - | 253 (100) | 16 (15) | ||
| Pfdhfr | |||||||
| N51I | N | 20 (11) | 240 (98) | 45 (23) | 8 (3) | 100 (89) | |
| I | 169 (89) | 5 (2) | 154 (77) | 245 (97) | 12 (11) | ||
| C59R | C | 29 (14) | 241 (98) | 100 (50) | 26 (10) | 102 (91) | |
| R | 180 (86) | 6 (2) | 99 (50) | 227 (90) | 10 (9) | ||
| S108N | S | 5 (2) | 243 (99) | 44 (22) | 0 (0) | 72 (64) | |
| N | 204 (98) | 2(1) | 155 (78) | 253 (100) | 40 (36) | ||
| Region | Study Site | N (%) | ||||
|---|---|---|---|---|---|---|
| Pfdhps | Pfdhfr | Pfdhfr/Pfdhps | Pfdhfr/Pfdhps | |||
| Total | N108 | I51R59N108 | I51R59N108/G437 | I51R59N109/G437E540 | ||
| North | ||||||
| KG | 47 | 45 (96) | 33 (70) | 17 (36) | 0 | |
| PZ | 52 | 52 (100) | 37 (71) | 28 (54) | 0 | |
| Central | ||||||
| KD | 52 | 49 (94) | 43 (83) | 3 (6) | 0 | |
| South | ||||||
| DO | 23 | 23 (100) | 0 | 0 | 0 | |
| AD | 36 | 35 (97) | 3 (8) | 1 (3) | 0 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dieng, C.C.; Gonzalez, L.; Pestana, K.; Dhikrullahi, S.B.; Amoah, L.E.; Afrane, Y.A.; Lo, E. Contrasting Asymptomatic and Drug Resistance Gene Prevalence of Plasmodium falciparum in Ghana: Implications on Seasonal Malaria Chemoprevention. Genes 2019, 10, 538. https://doi.org/10.3390/genes10070538
Dieng CC, Gonzalez L, Pestana K, Dhikrullahi SB, Amoah LE, Afrane YA, Lo E. Contrasting Asymptomatic and Drug Resistance Gene Prevalence of Plasmodium falciparum in Ghana: Implications on Seasonal Malaria Chemoprevention. Genes. 2019; 10(7):538. https://doi.org/10.3390/genes10070538
Chicago/Turabian StyleDieng, Cheikh Cambel, Lauren Gonzalez, Kareen Pestana, Shittu B. Dhikrullahi, Linda E. Amoah, Yaw A. Afrane, and Eugenia Lo. 2019. "Contrasting Asymptomatic and Drug Resistance Gene Prevalence of Plasmodium falciparum in Ghana: Implications on Seasonal Malaria Chemoprevention" Genes 10, no. 7: 538. https://doi.org/10.3390/genes10070538
APA StyleDieng, C. C., Gonzalez, L., Pestana, K., Dhikrullahi, S. B., Amoah, L. E., Afrane, Y. A., & Lo, E. (2019). Contrasting Asymptomatic and Drug Resistance Gene Prevalence of Plasmodium falciparum in Ghana: Implications on Seasonal Malaria Chemoprevention. Genes, 10(7), 538. https://doi.org/10.3390/genes10070538