Identification of a Missense Variant in MFSD12 Involved in Dilution of Phaeomelanin Leading to White or Cream Coat Color in Dogs

 , , , , , ,
, , , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
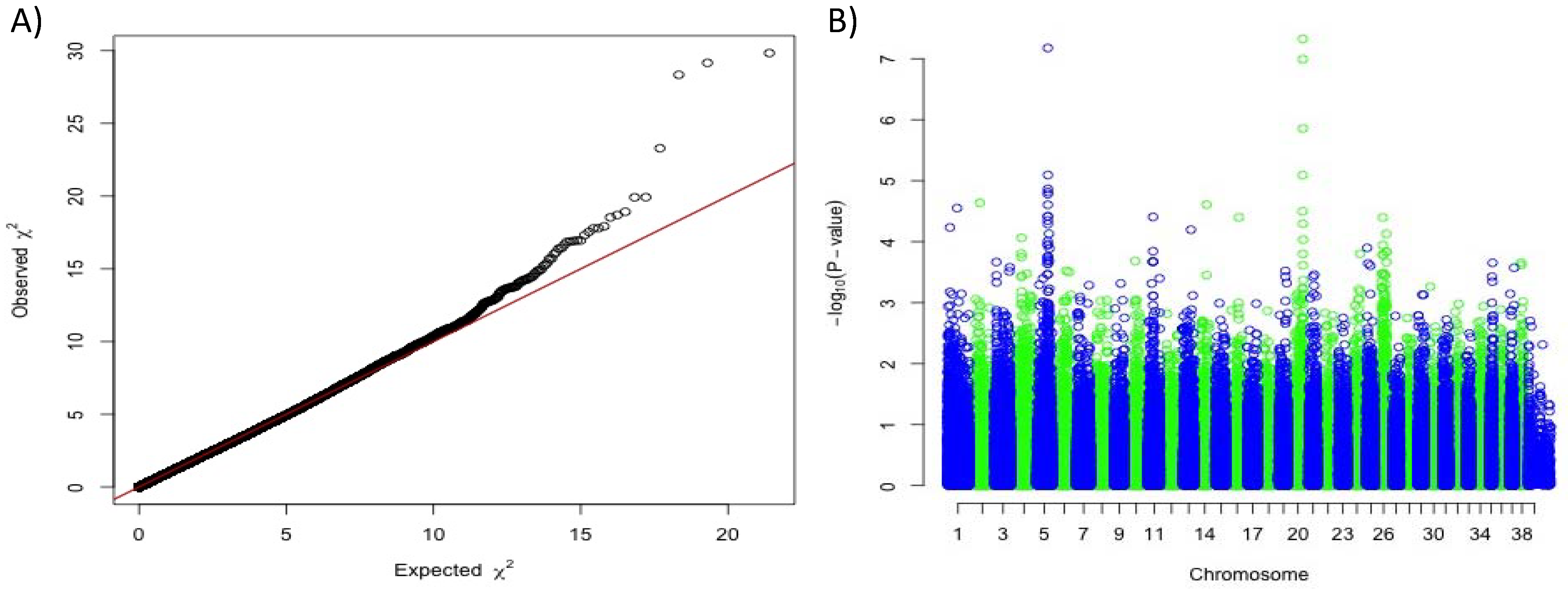
2.2. Genome-Wide Association Study (GWAS)
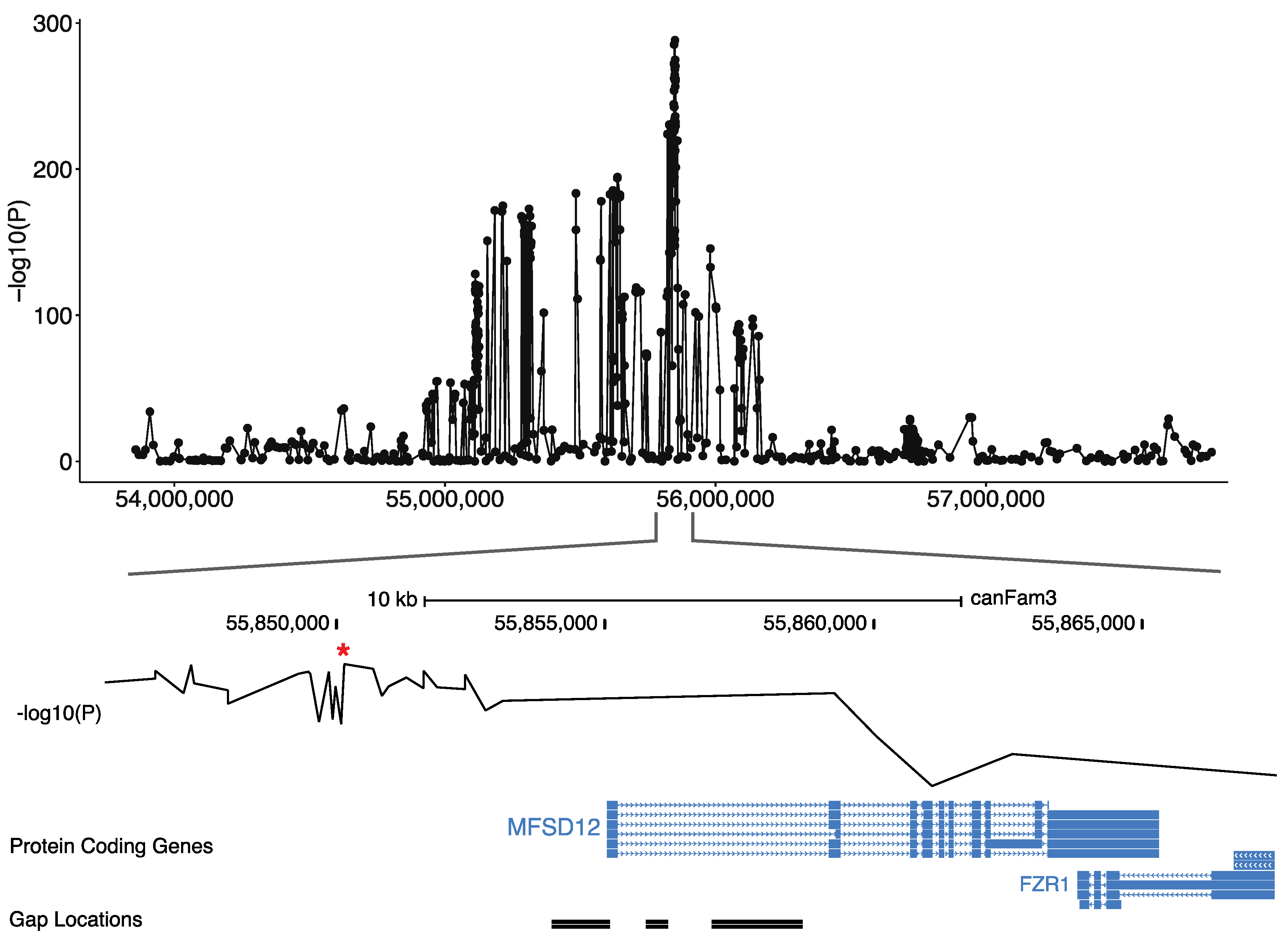
2.3. Filling Gap in the CanFam3 Sequence of MFSD12
2.4. Sequencing and RT-PCR on MFSD12
3. Results
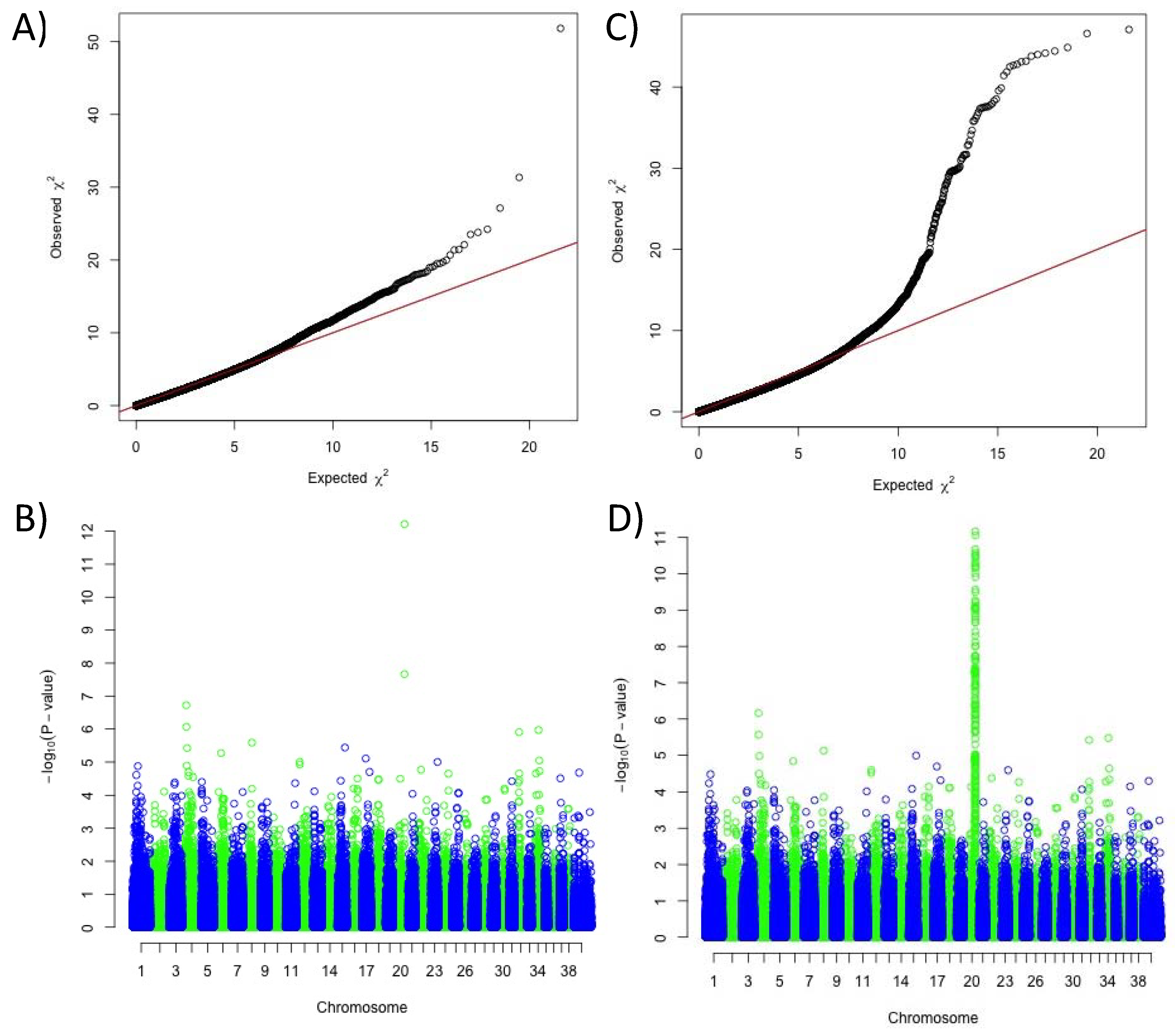
3.1. A Novel Association With Solid White on Canine Chromosome 20.
3.2. Identification of a Coding MFSD12 Variant Linked to Phaeomelanin Dilution
3.3. Validation of the MFSD12 Coding Variant
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Caduff, M.; Bauer, A.; Jagannathan, V.; Leeb, T. A single base deletion in the SLC45A2 gene in a Bullmastiff with oculocutaneous albinism. Anim. Genet. 2017, 48, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Wijesena, H.R.; Schmutz, S.M. A Missense Mutation in SLC45A2 Is Associated with Albinism in Several Small Long Haired Dog Breeds. J. Hered. 2015, 106, 285–288. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Winkler, P.A.; Gornik, K.R.; Ramsey, D.T.; Dubielzig, R.R.; Venta, P.J.; Petersen-Jones, S.M.; Bartoe, J.T. A partial gene deletion of SLC45A2 causes oculocutaneous albinism in Doberman pinscher dogs. PLoS ONE 2014, 9, e92127. [Google Scholar] [CrossRef] [PubMed]
- Caduff, M.; Bauer, A.; Jagannathan, V.; Leeb, T. OCA2 splice site variant in German Spitz dogs with oculocutaneous albinism. PLoS ONE 2017, 12, e0185944. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, M.F.; Van Cleave, P.S.; Glenn, K.L.; Carlstrom, L.P.; Ellinwood, N.M. Association of MITF with white spotting in Beagle crosses and Newfoundland dogs. Anim. Genet. 2006, 37, 606–607. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.K.; Baranowska, I.; Wade, C.M.; Salmon Hillbertz, N.H.C.; Zody, M.C.; Anderson, N.; Biagi, T.M.; Patterson, N.; Pielberg, G.R.; Kulbokas, E.J., III; et al. Efficient mapping of mendelian traits in dogs through genome-wide association. Nat. Genet. 2007, 39, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Leegwater, P.A.; van Hagen, M.A.; van Oost, B.A. Localization of white spotting locus in Boxer dogs on CFA20 by genome-wide linkage analysis with 1500 SNPs. J. Hered. 2007, 98, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.K.; Ruhe, A.L.; Robertson, K.R.; Loew, E.R.; Williams, D.C.; Neff, M.W. A de novo mutation in KIT causes white spotting in a subpopulation of German Shepherd dogs. Anim. Genet. 2013, 44, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Little, C.C. The Inheritance of Coat Color in Dogs, 1st US ed.; Comstock Publishing Associates: Ithaca, NY, USA, 1957. [Google Scholar]
- Sponenberg, D.P.; Rothschild, M.F. Genetics of coat colour and hair texture. In Genetics of the Dog; Ruvinsky, A., Sampson, J., Eds.; CABI Press: New York, NY, USA, 2001. [Google Scholar]
- Schmutz, S.M.; Berryere, T.G. The genetics of cream coat color in dogs. J. Hered. 2007, 98, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Ulve, R.; Rault, M.; Bahin, M.; Lagoutte, L.; Abadie, J.; De Brito, C.; Coindre, J.-M.; Botherel, N.; Rousseau, A.; Wucher, V.; et al. Discovery of Human-Similar Gene Fusions in Canine Cancers. Cancer Res. 2017, 77, 5721–5727. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Willer, C.J.; Ding, J.; Scheet, P.; Abecasis, G.R. MaCH: Using sequence and genotype data to estimate haplotypes and unobserved genotypes. Genet. Epidemiol. 2010, 34, 816–834. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan, V.; Drögemüller, C.; Leeb, T. Dog Biomedical Variant Database Consortium; A Comprehensive Biomedical Variant Catalogue Based on Whole Genome Sequencing of 6 Wolves and 584 Dogs. Anim. Genet. (submitted).
- Adhikari, K.; Mendoza-Revilla, J.; Sohail, A.; Fuentes-Guajardo, M.; Lampert, J.; Chacon-Duque, J.C.; Hurtado, M.; Villegas, V.; Granja, V.; Acuña-Alonzo, V.; et al. A GWAS in Latin Americans highlights the convergent evolution of lighter skin pigmentation in Eurasia. Nat. Commun. 2019, 10, 358. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.G.; Kelly, D.E.; Hansen, M.E.B.; Beltrame, M.H.; Fan, S.; Bowman, S.L.; Jewett, E.; Ranciaro, A.; Thompson, S.; Lo, Y.; et al. Loci associated with skin pigmentation identified in African populations. Science 2017, 358. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, J.; Leeb, T.; Mack, M.; Jagannathan, V.; Flury, C.; Bachmann, I.; McDonnell, S.M.; Maria, P.M.C.T.; Bellone, R. A Frameshift Variant in MFSD12 Explains the Mushroom Coat Color Dilution in Shetland Ponies [Internet]. PAG. 2019. Available online: https://plan.core-apps.com/pag_2019/abstract/343fb67f-5123-40b8-8885-76ad3141b62e (accessed on 15 May 2019).
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Coat Color | T/T | T/C or C/C |
|---|---|---|---|
| Poodle * | white | 8 | 0 |
| apricot/cream/red | 2 | 57 | |
| Old German Shepherd ** | white phaeomelanin | 44 | 5 |
| red or sable phaeomelanin | 4 | 54 | |
| Leonberger *** | diluted phaeomelanin | 7 | 0 |
| fawn sable | 0 | 118 | |
| All 3 breeds **** | diluted phaeomelanin | 59 | 5 |
| red or sable phaeomelanin | 6 | 229 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hédan, B.; Cadieu, E.; Botherel, N.; Dufaure de Citres, C.; Letko, A.; Rimbault, M.; Drögemüller, C.; Jagannathan, V.; Derrien, T.; Schmutz, S.; et al. Identification of a Missense Variant in MFSD12 Involved in Dilution of Phaeomelanin Leading to White or Cream Coat Color in Dogs. Genes 2019, 10, 386. https://doi.org/10.3390/genes10050386
Hédan B, Cadieu E, Botherel N, Dufaure de Citres C, Letko A, Rimbault M, Drögemüller C, Jagannathan V, Derrien T, Schmutz S, et al. Identification of a Missense Variant in MFSD12 Involved in Dilution of Phaeomelanin Leading to White or Cream Coat Color in Dogs. Genes. 2019; 10(5):386. https://doi.org/10.3390/genes10050386
Chicago/Turabian StyleHédan, Benoit, Edouard Cadieu, Nadine Botherel, Caroline Dufaure de Citres, Anna Letko, Maud Rimbault, Cord Drögemüller, Vidhya Jagannathan, Thomas Derrien, Sheila Schmutz, and et al. 2019. "Identification of a Missense Variant in MFSD12 Involved in Dilution of Phaeomelanin Leading to White or Cream Coat Color in Dogs" Genes 10, no. 5: 386. https://doi.org/10.3390/genes10050386
APA StyleHédan, B., Cadieu, E., Botherel, N., Dufaure de Citres, C., Letko, A., Rimbault, M., Drögemüller, C., Jagannathan, V., Derrien, T., Schmutz, S., Leeb, T., & André, C. (2019). Identification of a Missense Variant in MFSD12 Involved in Dilution of Phaeomelanin Leading to White or Cream Coat Color in Dogs. Genes, 10(5), 386. https://doi.org/10.3390/genes10050386