Cloning of pcB and pcA Gene from Gracilariopsis lemaneiformis and Expression of a Fluorescent Phycocyanin in Heterologous Host

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Genomic DNA Extraction and Gene Cloning of pcB and pcA
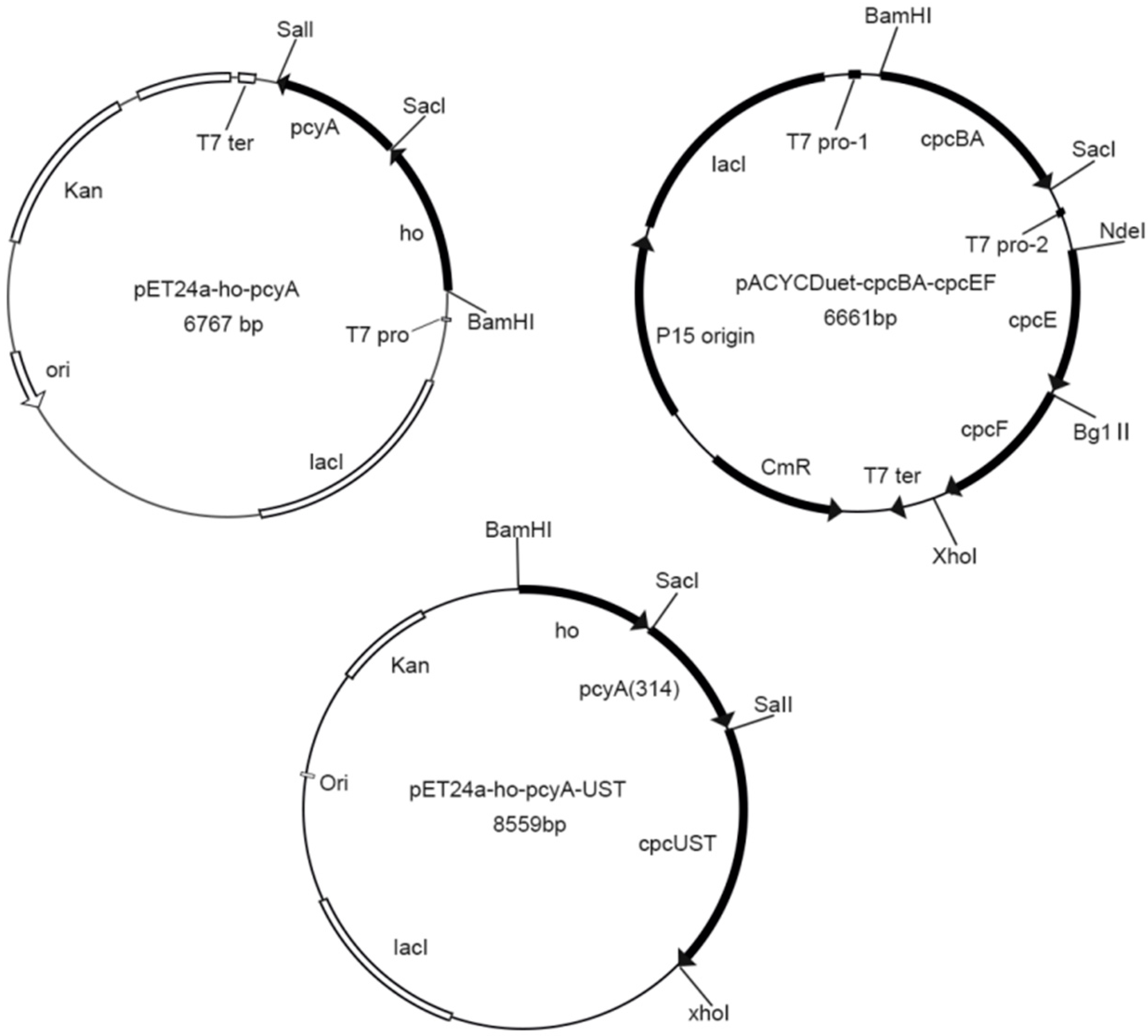
2.3. Vector Construction
2.4. Plasmid Transformation and Protein Expression
2.5. Recombinant Protein Analysis
2.6. Fluorescence Emission Spectra Analysis
3. Results
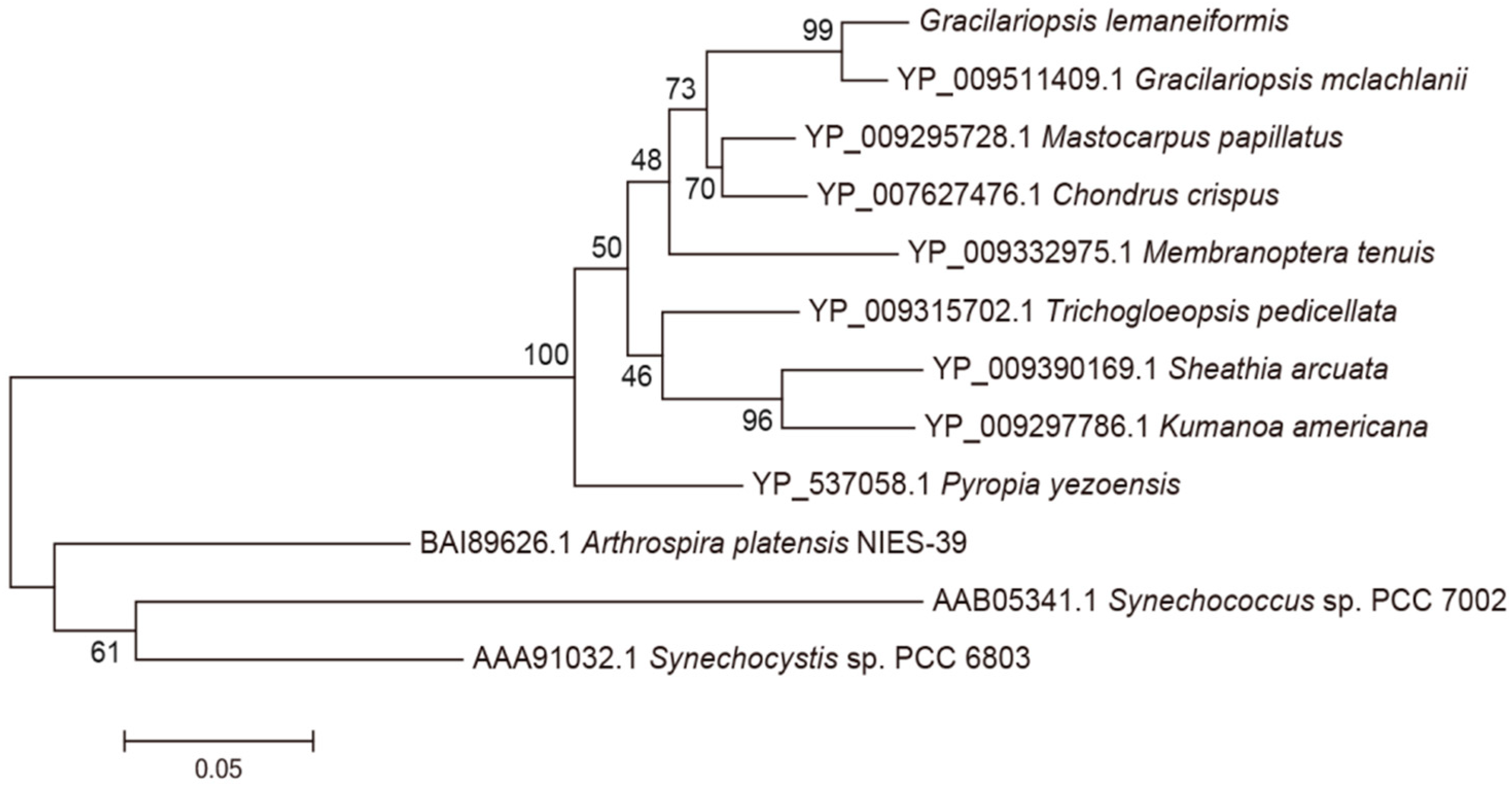
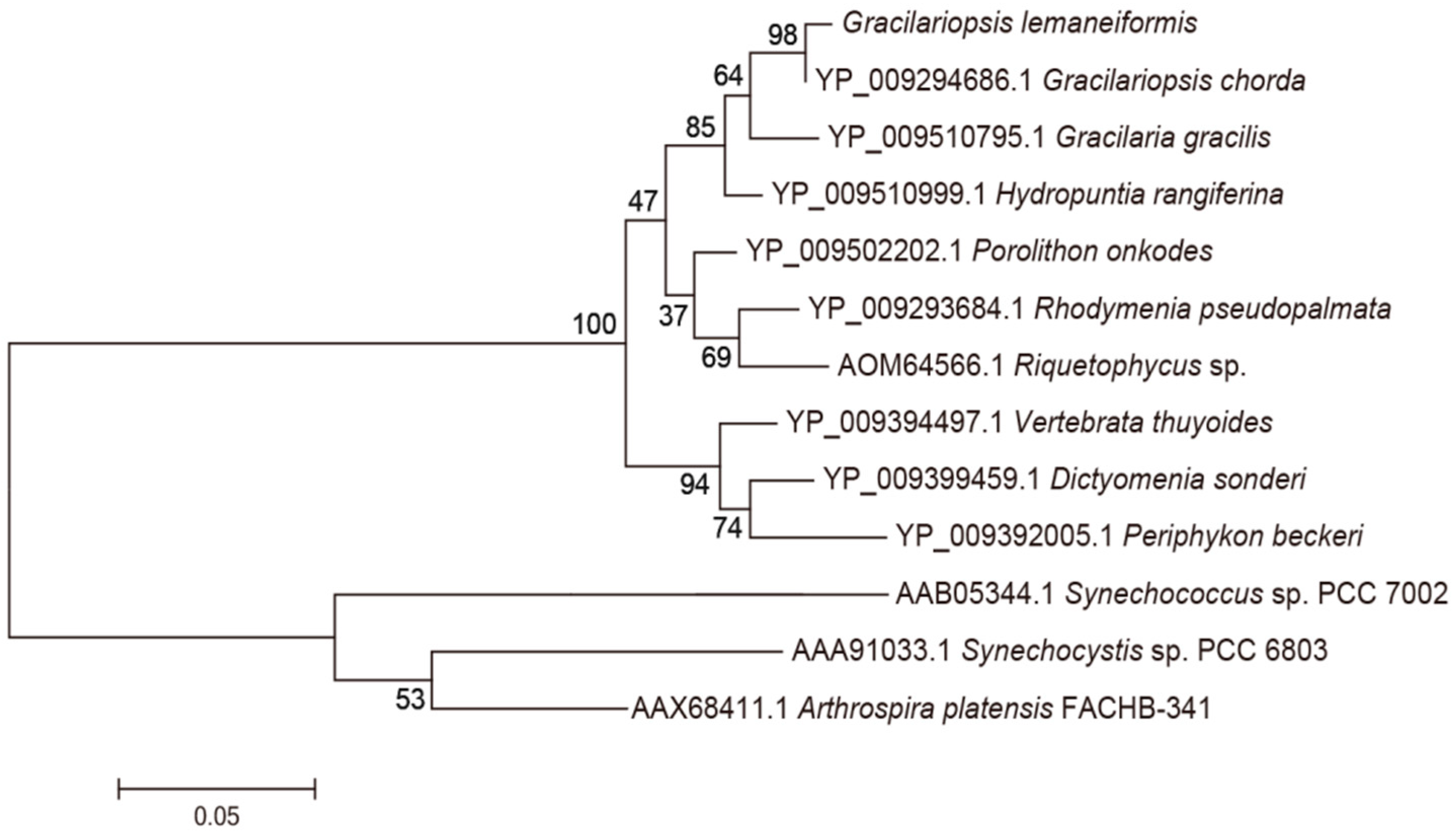
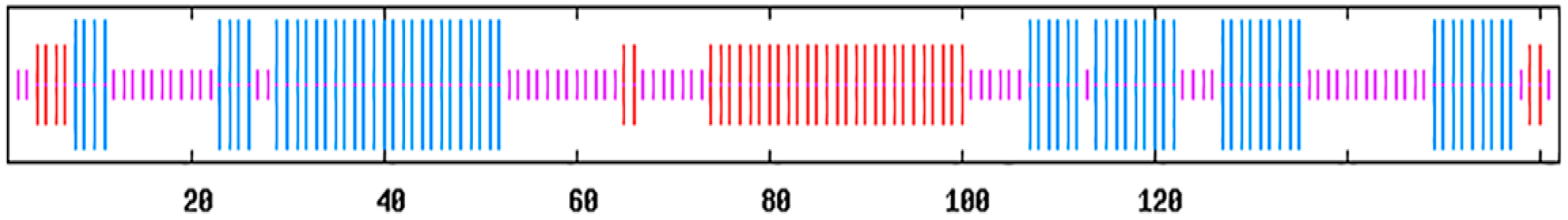
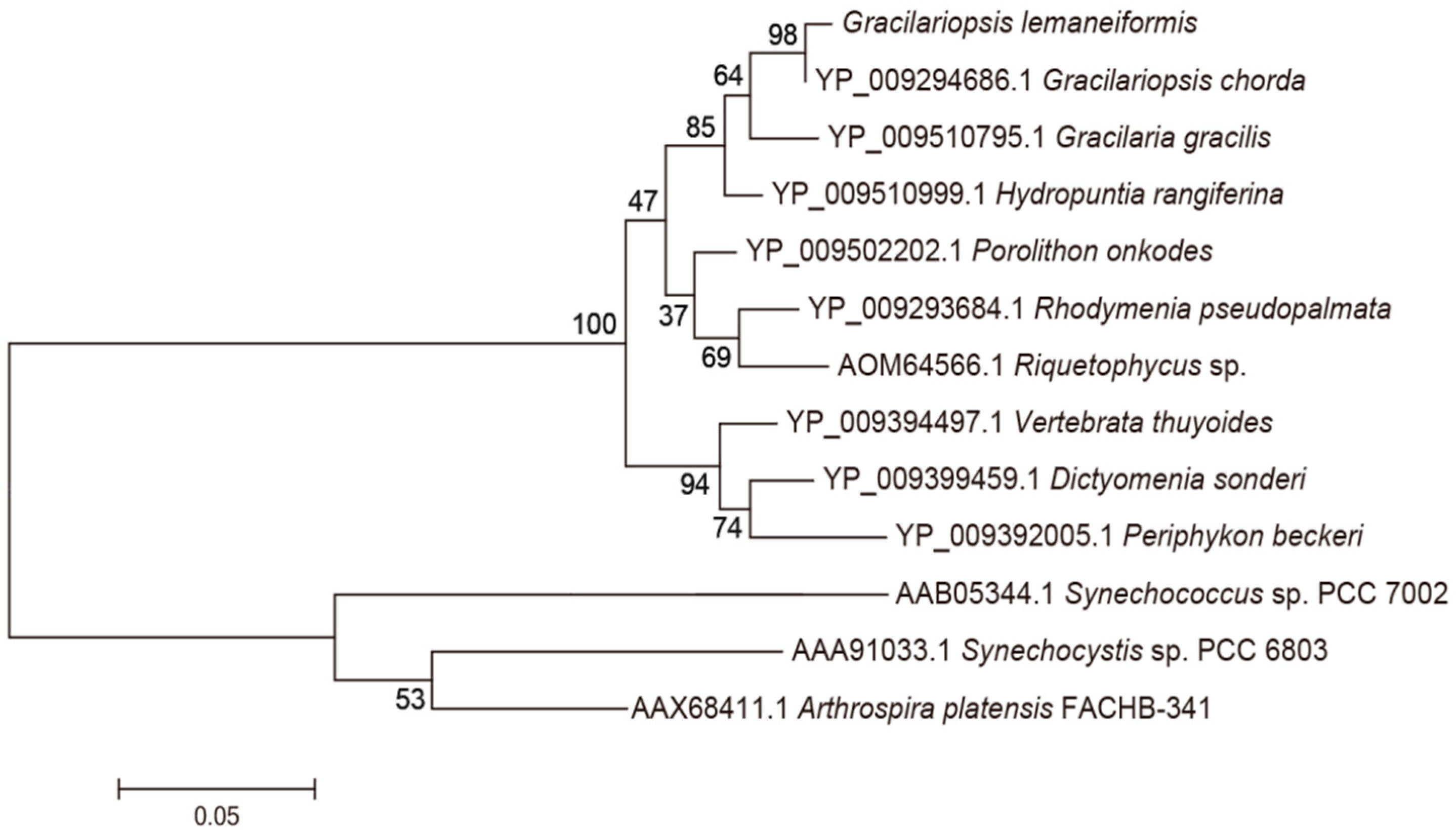
3.1. Analysis of the Genes pcB and pcA
3.2. Construction of Recombinant Strain
3.3. Expression of the Recombinant Proteins
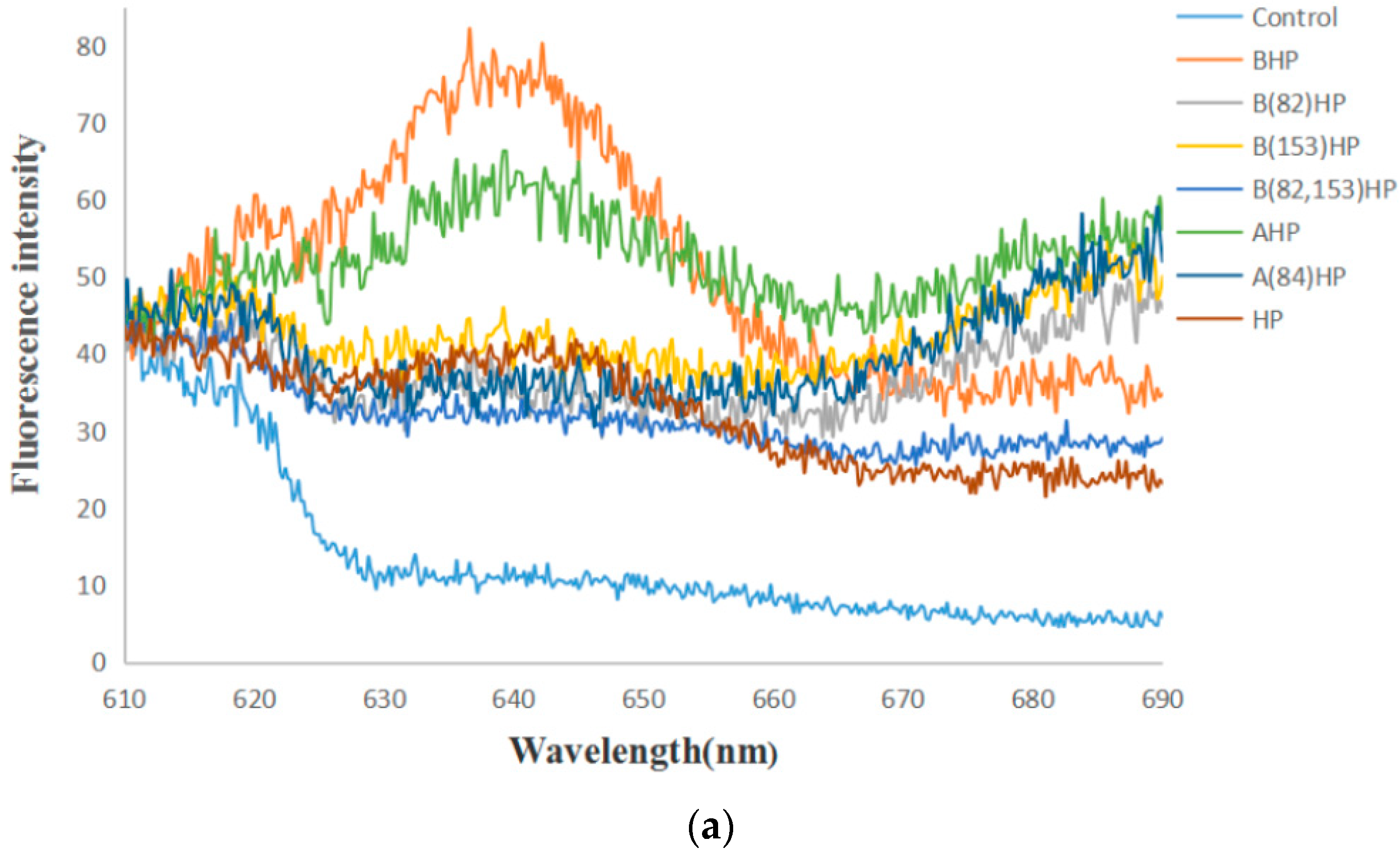
3.4. Fluorescence Emission Spectra
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Glazer, A.N. Light guides. Directional energy transfer in a photosynthetic antenna. J. Biol. Chem. 1989, 264, 1–4. [Google Scholar] [PubMed]
- Green, B.R.; Parson, W.W. Light-Harvesting Antennas in Photosynthesis; Advances in Photosynthesis & Respiration; Springer: Amsterdam, The Netherlands, 2003; Volume 13. [Google Scholar]
- Glazer, A.N. Adaptive variations in phycobilisome structure. Adv. Mol. Cell Biol. 1994, 10, 119–149. [Google Scholar]
- Bhat, V.B.; Madyastha, K.M. C-phycocyanin: A potent peroxyl radical scavenger in vivo and in vitro. Biochem. Biophys. Res. Commun. 2000, 275, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Nyman, E.S.; Hynninen, P.H. Research advances in the use of tetrapyrrolic photosensitizers for photodynamic therapy. J. Photochem. Photobiol. B Biol. 2004, 73, 1–28. [Google Scholar] [CrossRef]
- Shih, C.M.; Cheng, S.N.; Wong, C.S.; Kuo, Y.L.; Chou, T.C. Antiinflammatory and antihyperalgesic activity of C-phycocyanin. Anesth. Analg. 2009, 108, 1303–1310. [Google Scholar] [CrossRef]
- Beale, S.I. Biosynthesis of cyanobacterial tetrapyrrole pigments hemes, chlorophylls, and phycobilins. Mol. Biol. Cyanobacteria 1994, 1, 519–558. [Google Scholar]
- Sidler, W.A. Phycobilisome and phycobiliprotein structures, molecular biology of cyanobacteria. In The Molecular Biology of Cyanobacteria; Springer: Dordrecht, The Netherlands, 1994; Volume 1, pp. 139–216. [Google Scholar]
- Wiethaus, J.; Busch, A.W.U.; Dammeyer, T.; Frankenberg-Dinkel, N. Phycobiliproteins in Prochlorococcus marinus: Biosynthesis of pigments and their assembly into proteins. Eur. J. Cell Biol. 2010, 89, 1005–1010. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, D.; Zhang, L.; Zhou, M.; Bohm, S.; Bubenzer, C.; Scheer, H. Chromophore attachment in phycocyanin. Functional amino acids of phycocyanobilin-alpha-phycocyanin lyase and evidence for chromophore binding. FEBS J. 2010, 273, 1262–1274. [Google Scholar] [CrossRef]
- Biswas, A.; Vasquez, Y.M.; Dragomani, T.M.; Kronfel, M.L.; Williams, S.R.; Alvey, R.M.; Bryant, D.A.; Schluchter, W.M. Biosynthesis of cyanobacterial phycobiliproteins in Escherichia coli: Chromophorylation efficiency and specificity of all bilin lyases from Synechococcus sp. strain PCC 7002. Appl. Environ. Microbiol. 2010, 76, 2729. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kang, Z.; Chen, J.; Du, G. Optimization of the heme biosynthesis pathway for the production of 5-aminolevulinic acid in Escherichia coli. Sci. Rep. 2015, 5, 8584. [Google Scholar] [CrossRef]
- Frankenberg, N.; Mukougawa, K.; Kohchi, T.; Lagarias, J.C. Functional genomic analysis of the HY2 family of ferredoxin-dependent bilin reductases from oxygenic photosynthetic organisms. Plant Cell 2001, 13, 965–978. [Google Scholar] [CrossRef]
- Zhao, K.H.; Su, P.; Tu, J.M.; Wang, X.; Liu, H.; Plöscher, M.; Eichacker, L.; Yang, B.; Zhou, M.; Scheert, H. Phycobilin:Cystein-84 biliprotein lyase, a near-universal lyase for cysteine-84-binding sites in cyanobacterial phycobiliproteins. Proc. Natl. Acad. Sci. USA 2007, 104, 14300–14305. [Google Scholar] [CrossRef]
- Shen, G.; Schluchter, W.M.; Bryant, D.A. Biogenesis of phycobiliproteins: I. cpcS-I and cpcU mutants of the cyanobacterium Synechococcus sp. PCC 7002 define a heterodimeric phyococyanobilin lyase specific for β-phycocyanin and allophycocyanin subunits. J. Biol. Chem. 2008, 283, 7503–7512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Feng, X.T.; Wu, F.; Ding, Y.; Zang, X.N.; Zhang, X.C.; Yuan, D.Y.; Zhao, B.R. Molecular cloning and expression analysis of a new bilin lyase: The cpcT gene encoding a bilin lyase responsible for attachment of phycocyanobilin to Cys-153 on the β-subunit of phycocyanin in Arthrospira platensis FACHB314. Gene 2014, 544, 191–197. [Google Scholar] [CrossRef]
- Tooley, A.J.; Glazer, A.N. Biosynthesis of the cyanobacterial light-harvesting polypeptide phycoerythrocyanin holo-α subunit in a heterologous host. J. Bacteriol. 2002, 184, 4666–4671. [Google Scholar] [CrossRef]
- Tooley, A.J.; Cai, Y.A.; Glazer, A.N. Biosynthesis of a fluorescent cyanobacterial C-phycocyanin holo-α subunit in a heterologous host. Proc. Natl. Acad. Sci. USA 2001, 98, 10560–10565. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ge, B.; Guan, X.; Zhang, W.; Qin, S. Combinational biosynthesis of a fluorescent cyanobacterial holo-α-allophycocyanin in Escherichia coli. Biotechnol. Lett. 2008, 30, 1001. [Google Scholar] [CrossRef]
- Yi, J.; Xu, D.; Zang, X.; Yuan, D.; Zhao, B.; Tang, L.; Tan, Y.; Zhang, X. Lyase activities of heterologous CpcS and CpcT for phycocyanin holo-β-subunit from Arthrospira platensis in Escherichia coli. J. Ocean Univ. China 2014, 13, 497–502. [Google Scholar] [CrossRef]
- Wu, F.; Zang, X.; Zhang, X.; Zhang, R.; Huang, X.; Hou, L.; Jiang, M.; Liu, C.; Pang, C. Molecular cloning of cpcU and heterodimeric bilin lyase activity analysis of CpcU and CpcS for attachment of phycocyanobilin to Cys-82 on the β-subunit of phycocyanin in Arthrospira platensis FACHB314. Molecules 2016, 21, 357. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zang, X.; Wu, F.; Jin, Y.; Wang, H.; Liu, C.; Ding, Y.; He, B.; Xiao, D.; Song, X. Transcriptome sequencing of Gracilariopsis lemaneiformis to analyze the genes related to optically active phycoerythrin synthesis. PLoS ONE 2017, 12, e0170855. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.L.; Church, D.M.; Federhen, S.; Lash, A.E.; Madden, T.L.; Pontius, J.U.; Schuler, G.D.; Schriml, L.M.; Sequeira, E.; Tatusova, T.A. Database resources of the national center for biotechnology. Nucleic Acids Res. 2003, 31, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.; Saunée, N.A.; Williams, S.R.; Gallo, E.F.; Schluchter, W.M.; Bryant, D.A. Identification and characterization of a new class of bilin lyase the cpcT gene encodes a bilin lyase responsible for attachment of phycocyanobilin to CYS-153 on the β-subunit of phycocyanin in Synechococcus sp. PCC 7002. J. Biol. Chem. 2006, 281, 17768–17778. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.G.; Chen, X.L.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z. Spectral changes of C-phycocyanin with different molar ratios of SPDP. Spectrosc. Spectr. Anal. 2008, 28, 1115–1117. [Google Scholar]
- Wiltshire, K.H.; Moldaenke, C. In vivo fluorescence of phycocyanin in combination with multispectral chlorophyll fluorescence spectroscopy—A powerful tool for the analysis of cyanobacteria and algae. Sci. Access 2001, 3, 1. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain and Plasmid | Source | Application |
|---|---|---|
| asapMD19-T | TaKaRa (Dalian, China) | Cloning vector |
| pACYCDuet-1 | Novagen (Germany) | Expression vector |
| pET24a | Novagen (Germany) | Expression vector |
| pET24a-ho-pcyA | Our laboratory | Express heme oxygenase and ferredoxin oxidoreductase (HO and PcyA) |
| pACYCDuet-cpcBA-cpcEF | Our laboratory | Expression of apo-phycocyanin and chromophore lyase (CpcE and CpcF) |
| pET24a-ho-pcyA-UST | Our laboratory | Express heme oxygenase, ferredoxin oxidoreductase and chromophore lyase (CpcU, CpcS and CpcT) |
| Primer Names | Primer Sequences | Annealing Temperature (°C) |
|---|---|---|
| pcB primers | pcB-F:5′-GGATCCATGCTAGATGCATTTGCTAAAGTTGT-3′ | 57.8 |
| pcB-R:5′-GAGCTCTTATACTACTGCAACTGCTG-3′ | 55.7 | |
| pcA primers | pcA-F:5′-GGATCCATGAAAACACCTATAACAGA-3′ | 52.6 |
| pcA-R:5′-GAGCTCTTAAGTTAAAGCATTAATGGCATAGTCT-3′ | 56.7 | |
| pcBA primers | pcB-F:5′-GGATCCATGCTAGATGCATTTGCTAAAGTTGT-3′ | 57.8 |
| pcA-R:5′-GAGCTCTTAAGTTAAAGCATTAATGGCATAGTCT-3′ | 56.7 |
| Primer Names | Primer Sequences | Annealing Temperature (°C) |
|---|---|---|
| pcB mutation primers 1 | pcB(82)-F:5′-GCAGCTGCTTTAAGAGATATGGAGATTGTTTTAA-3′ | 56.7 |
| pcB(82)-R:5′-CATTCTACGACTCGTATATGCATTACCACCT-3′ | 57.7 | |
| pcB mutation primers 2 | pcB(153)-F:5′-GGAGATGCTAGCTCATTAATCGCTGAATTAGGA-3′ | 59.1 |
| pcB(153)-R:5′-TAGAGTTACACCATTTGTATTATTAGCTAAACTTA-3′ | 53.3 | |
| pcA mutation primers | pcA(84)-F:5′-AAAAGCTGCACGTGA CATAGGATAT-3′ | 52.3 |
| pcA(84)-R:5′-GCTTTTCCTATTGCACTAGATG-3′ | 49.4 |
| Names of the Transformed | Expression Vectors | E. coli Strains |
|---|---|---|
| Control | pACYCDuet-1 | pET-24a (+) |
| BAHP | pACYCDuet-pcBA | pET24a-ho-pcyA |
| BHP | pACYCDuet-pcB | pET24a-ho-pcyA |
| B(82)HP | pACYCDuet-pcB(C82A) | pET24a-ho-pcyA |
| B(153)HP | pACYCDuet-pcB(C153A) | pET24a-ho-pcyA |
| B(82,153)HP | pACYCDuet-pcB(C82A,C153A) | pET24a-ho-pcyA |
| AHP | pACYCDuet-pcA | pET24a-ho-pcyA |
| A(84)HP | pACYCDuet-pcA(C84A) | pET24a-ho-pcyA |
| BAEFHPUST | pACYCDuet-pcBA-cpcEF | pET24a-ho-pcyA-cpcUST |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, D.; Zang, X.; Guo, Y.; Xiao, D.; Cao, X.; Liu, Z.; Zhang, F.; Jin, Y.; Shi, J.; Wang, Z.; et al. Cloning of pcB and pcA Gene from Gracilariopsis lemaneiformis and Expression of a Fluorescent Phycocyanin in Heterologous Host. Genes 2019, 10, 322. https://doi.org/10.3390/genes10050322
Sun D, Zang X, Guo Y, Xiao D, Cao X, Liu Z, Zhang F, Jin Y, Shi J, Wang Z, et al. Cloning of pcB and pcA Gene from Gracilariopsis lemaneiformis and Expression of a Fluorescent Phycocyanin in Heterologous Host. Genes. 2019; 10(5):322. https://doi.org/10.3390/genes10050322
Chicago/Turabian StyleSun, Deguang, Xiaonan Zang, Yalin Guo, Dongfang Xiao, Xuexue Cao, Zhu Liu, Feng Zhang, Yuming Jin, Jiawei Shi, Zhendong Wang, and et al. 2019. "Cloning of pcB and pcA Gene from Gracilariopsis lemaneiformis and Expression of a Fluorescent Phycocyanin in Heterologous Host" Genes 10, no. 5: 322. https://doi.org/10.3390/genes10050322
APA StyleSun, D., Zang, X., Guo, Y., Xiao, D., Cao, X., Liu, Z., Zhang, F., Jin, Y., Shi, J., Wang, Z., Li, R., & Yangzong, Z. (2019). Cloning of pcB and pcA Gene from Gracilariopsis lemaneiformis and Expression of a Fluorescent Phycocyanin in Heterologous Host. Genes, 10(5), 322. https://doi.org/10.3390/genes10050322