A Magnesium Transport Protein Related to Mammalian SLC41 and Bacterial MgtE Contributes to Circadian Timekeeping in a Unicellular Green Alga


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ostreococcus tauri Cell Lines
2.2. Verification of Overexpressing Lines
2.3. Bioinformatics Analysis
3. Results
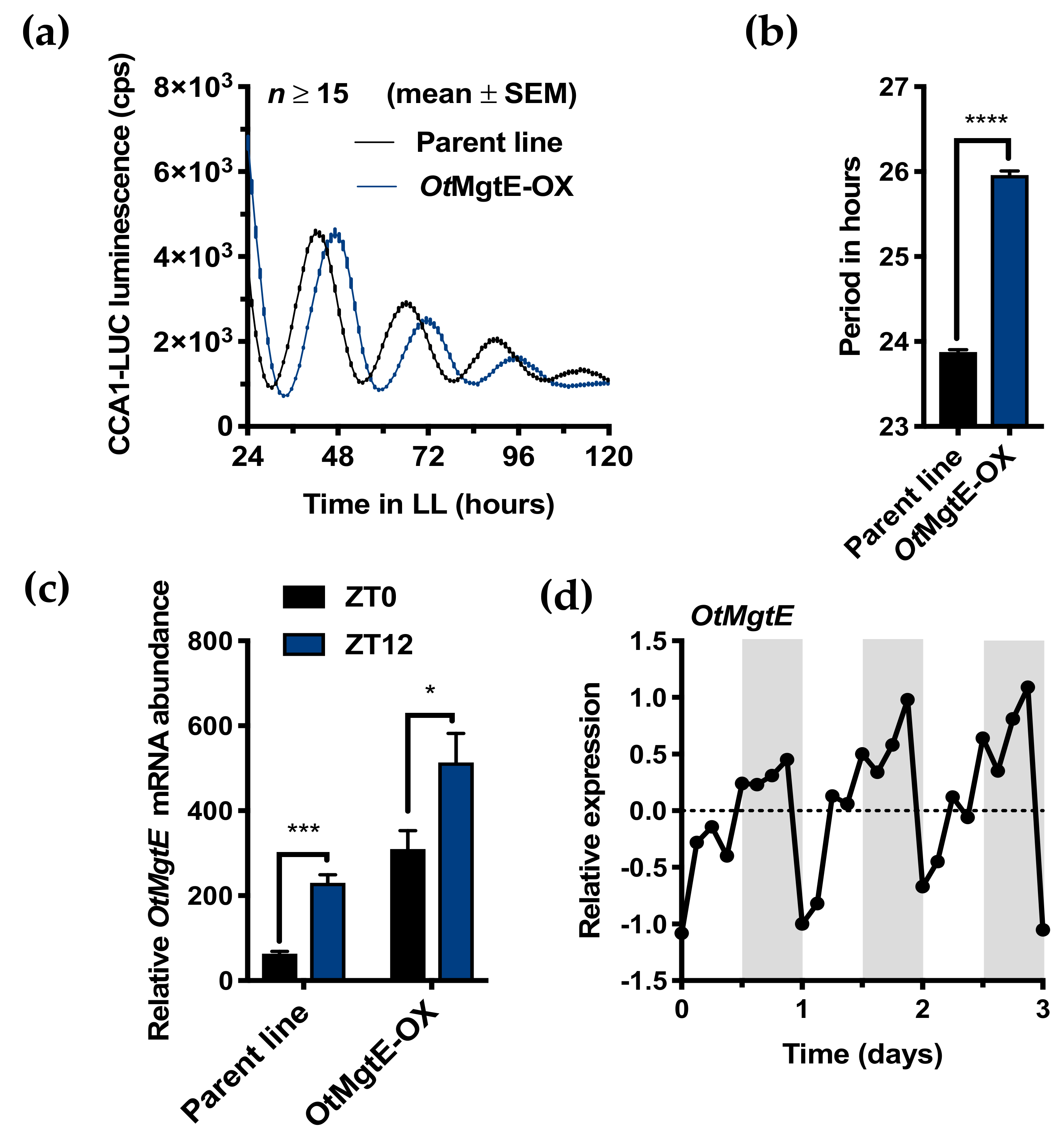
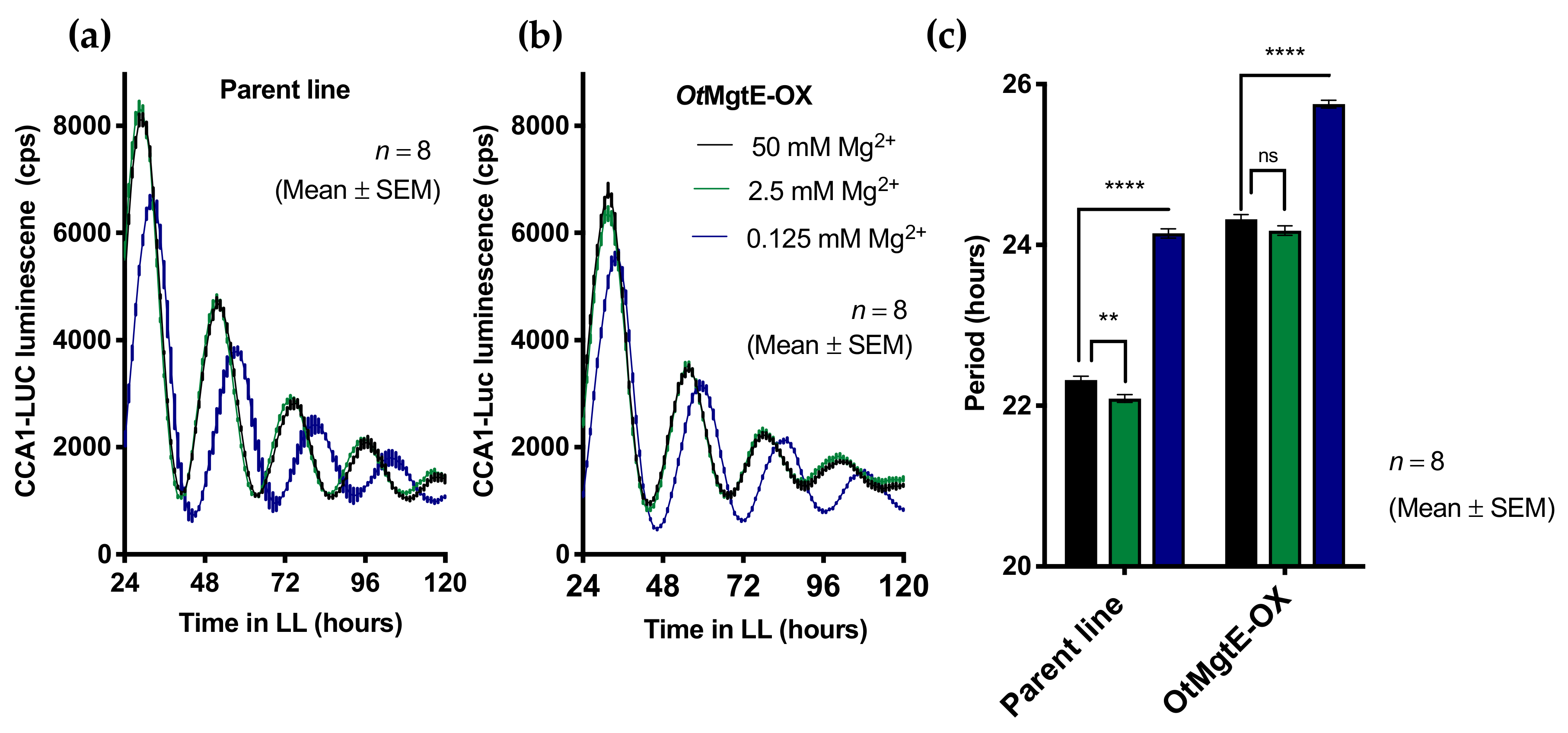
3.1. OtMgtE Overexpression Lengthens Circadian Period
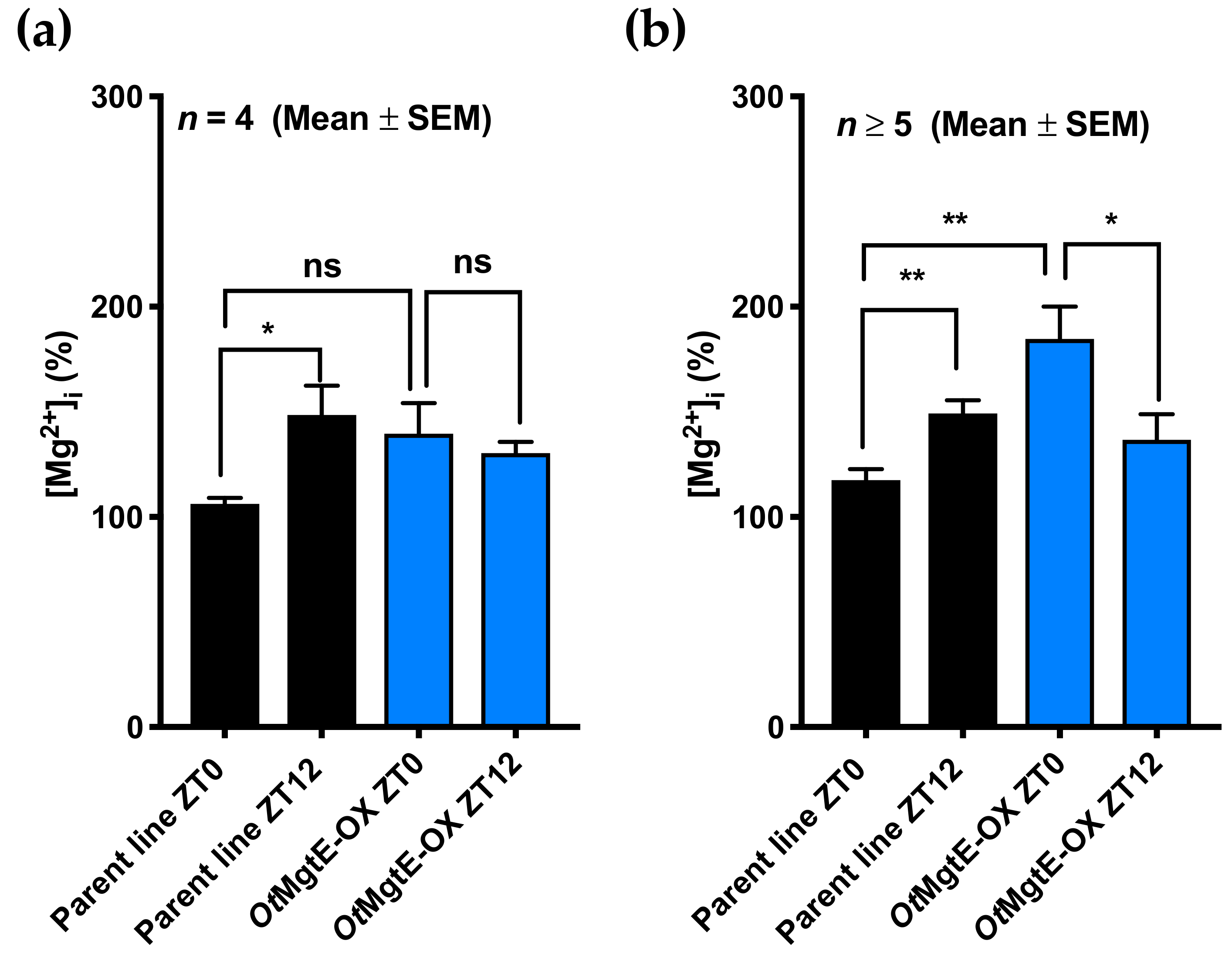
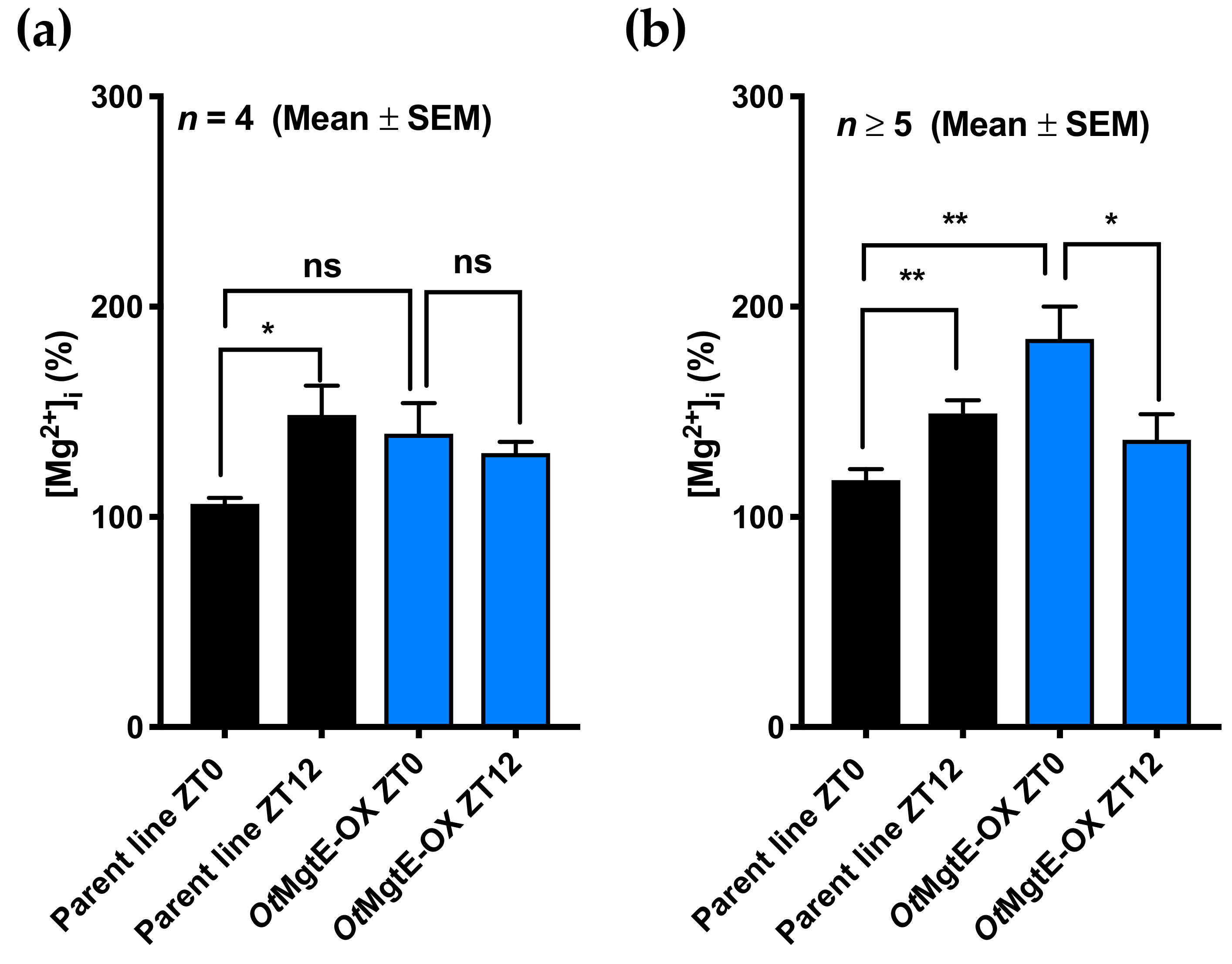
3.2. Effect of OtMgtE Overexpression on Intracellular Magnesium
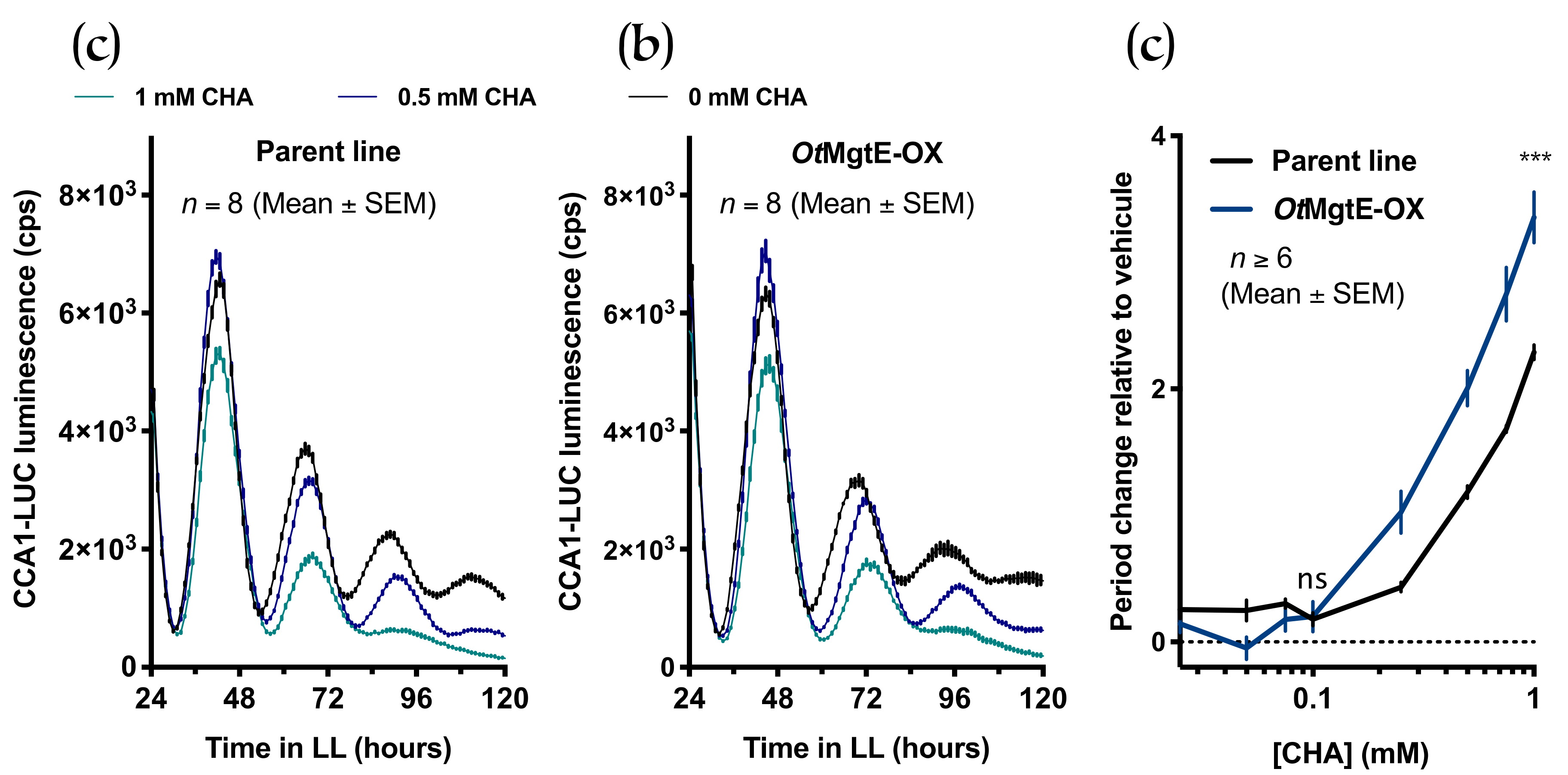
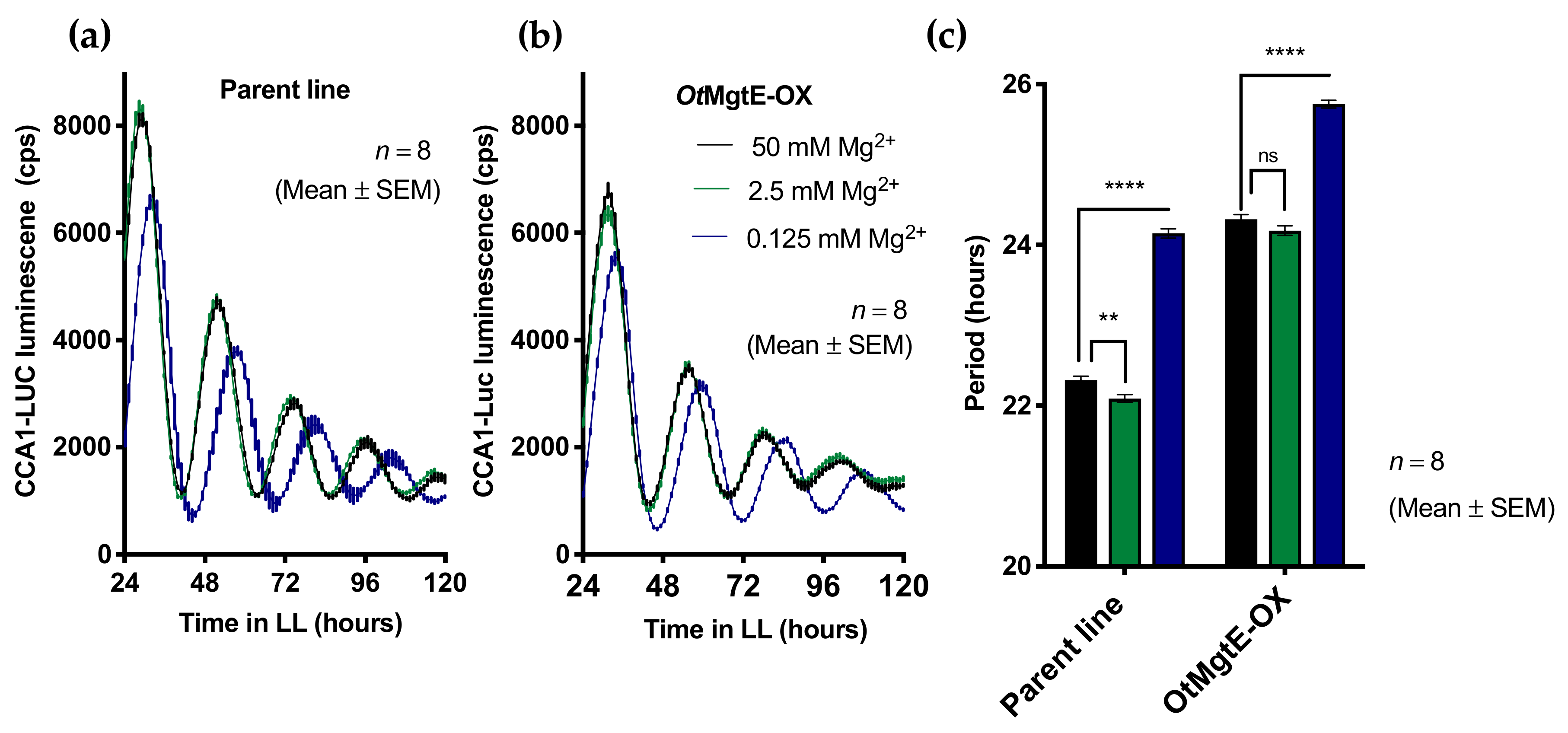
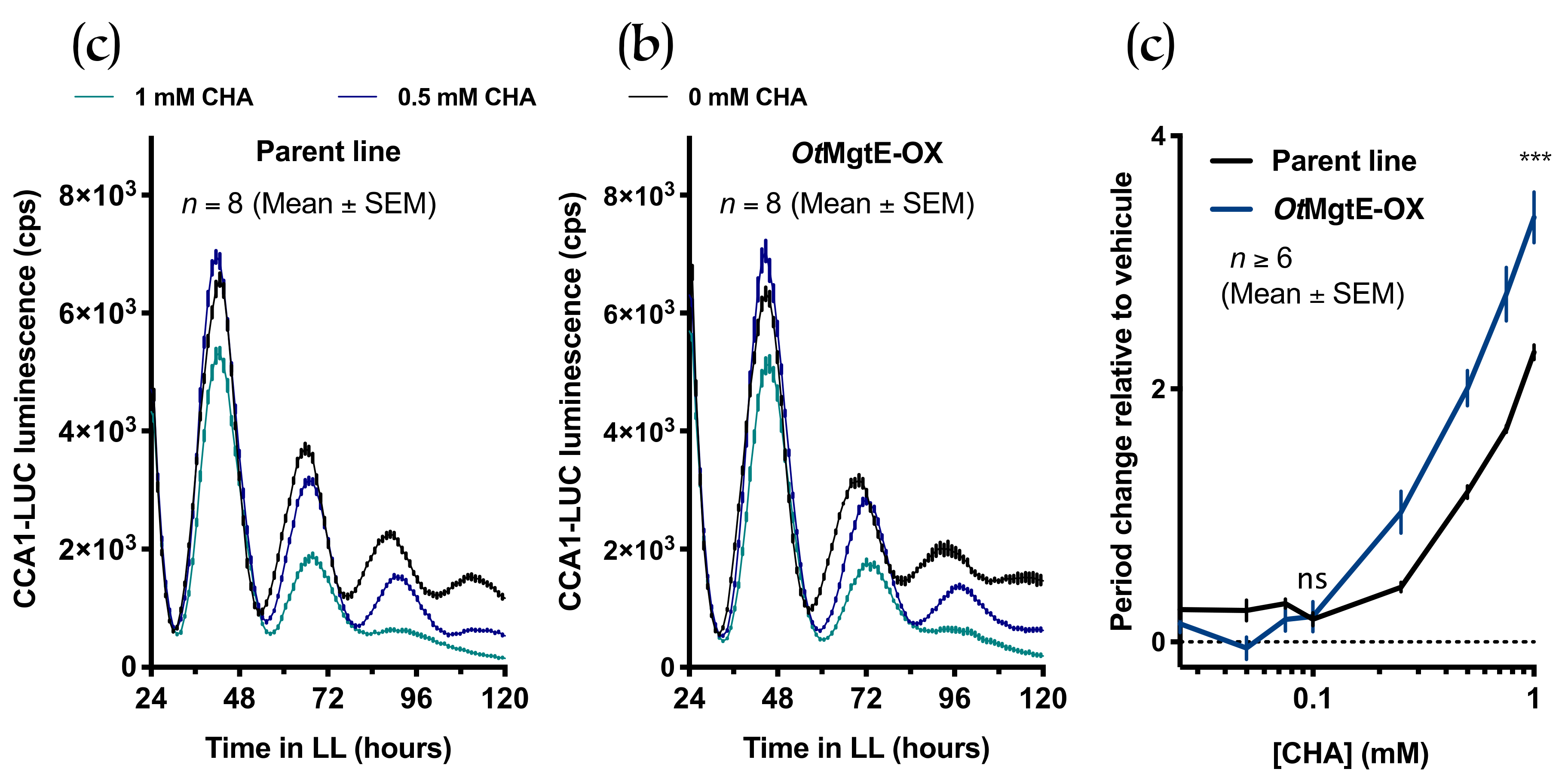
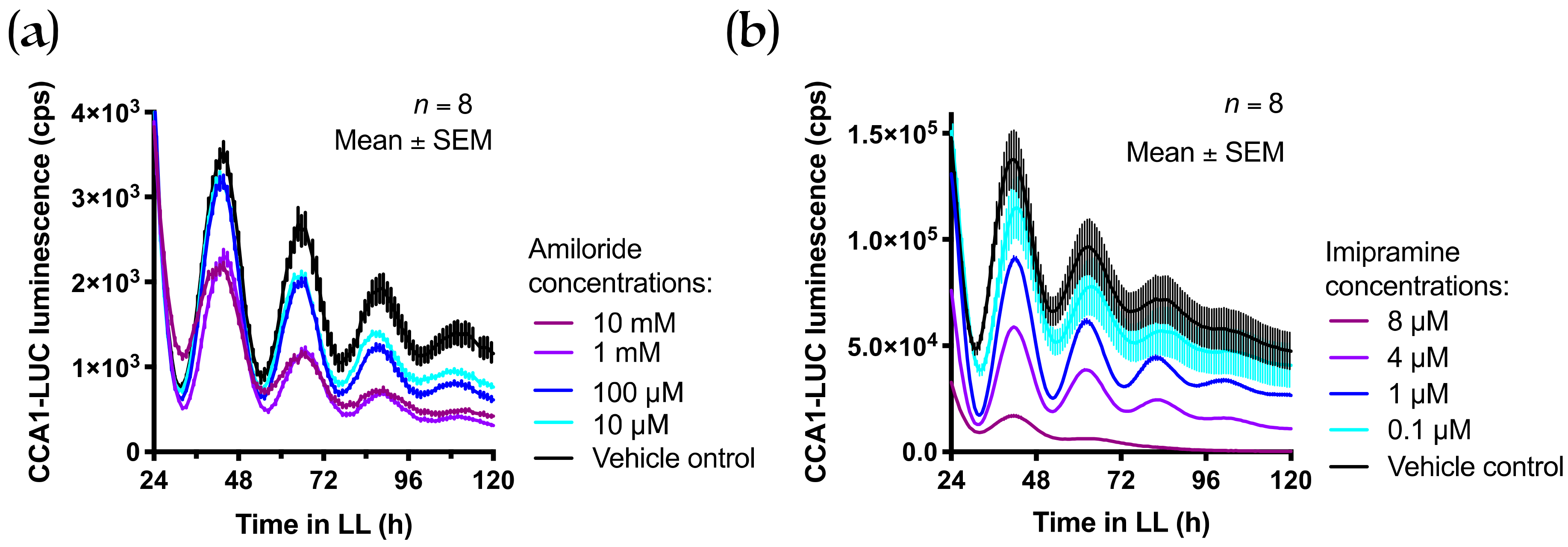
3.3. Treatment with Cobalt(III)hexamine and Low Extracellular Mg2+
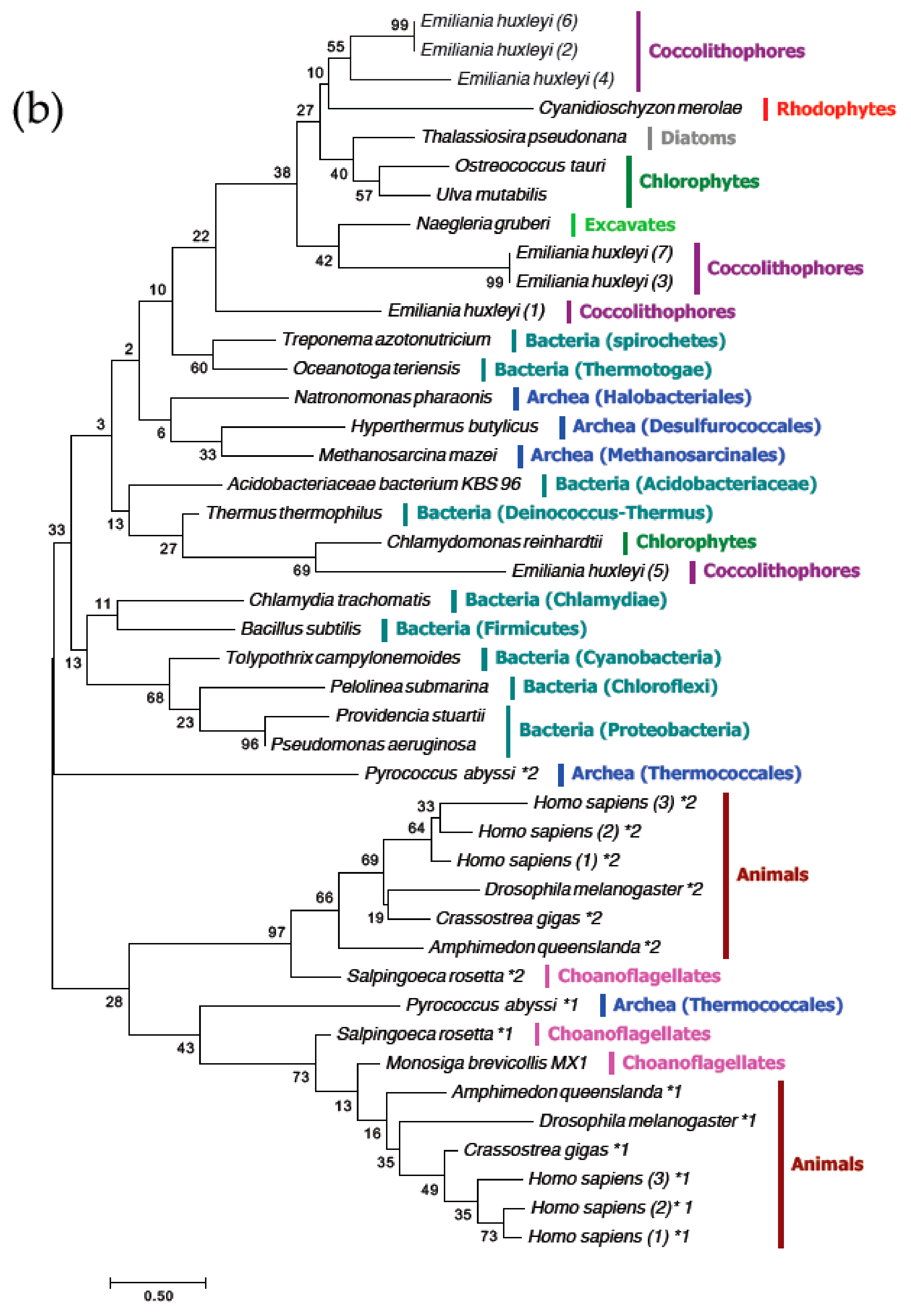
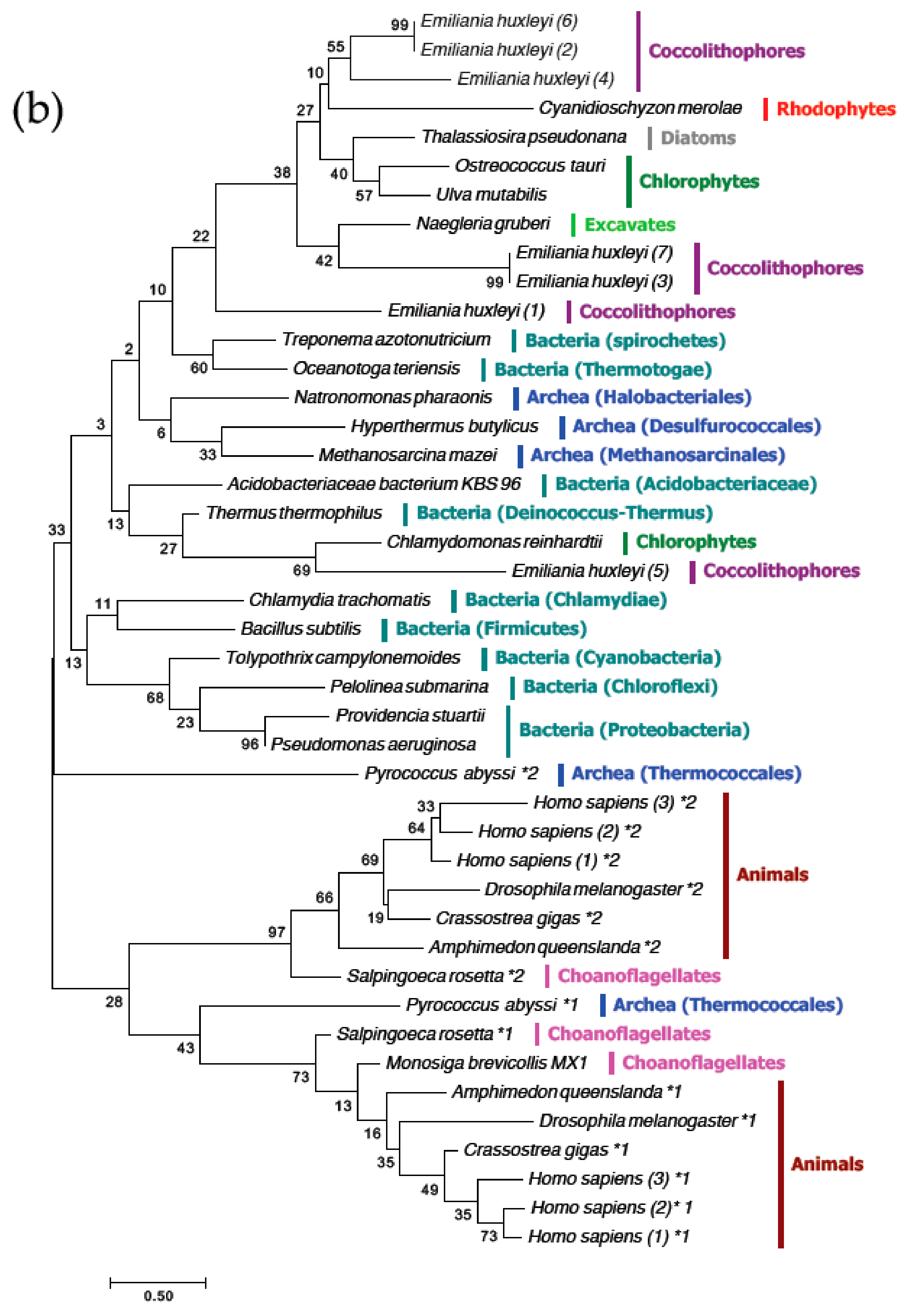
3.4. The MgtE Domains in OtMgtE and Animal SLC41 are Distinct
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dunlap, J.C. Molecular bases for circadian clocks. Cell 1999, 96, 271–290. [Google Scholar] [CrossRef]
- Bell-Pedersen, D.; Cassone, V.M.; Earnest, D.J.; Golden, S.S.; Hardin, P.E.; Thomas, T.L.; Zoran, M.J. Circadian rhythms from multiple oscillators: Lessons from diverse organisms. Nat. Rev. Drug. Discov. 2005, 4, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Pittendrigh, S. Circadian rhythms and the circadian organization of living systems. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 159–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.E.; Kay, S.A. Clocks not winding down: unravelling circadian networks. Nat. Rev. Mol. Cell Biol. 2010, 11, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Van Ooijen, G.; Millar, A.J. Non-transcriptional oscillators in circadian timekeeping. Trends Biochem. Sci. 2012, 37, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.C.; O’Neill, J.S. Non-transcriptional processes in circadian rhythm generation. Curr. Opin. Physiol. 2018, 5, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Tomita, J.; Nakajima, M.; Kondo, T.; Iwasaki, H. No transcription-translation feedback in circadian rhythm of kaic phosphorylation. Science 2005, 307, 251–254. [Google Scholar] [CrossRef]
- Nakajima, M.; Imai, K.; Ito, H.; Nishiwaki, T.; Murayama, Y.; Iwasaki, H.; Oyama, T.; Kondo, T. Reconstitution of circadian oscillation of cyanobacterial kaic phosphorylation in vitro. Science 2005, 308, 414–415. [Google Scholar] [CrossRef]
- O-Neill, J.S.; Reddy, A.B. Circadian clocks in human red blood cells. Nature 2011, 469, 498–504. [Google Scholar] [CrossRef]
- O’Neill, J.S.; van Ooijen, G.; Dixon, L.E.; Troein, C.; Corellou, F.; Bouget, F.-Y.; Reddy, A.B.; Millar, A.J. Circadian rhythms persist without transcription in a eukaryote. Nature 2011, 469, 554–558. [Google Scholar] [CrossRef]
- Edgar, R.S.; Green, E.W.; Zhao, Y.; van Ooijen, G.; Olmedo, M.; Qin, X.; Xu, Y.; Pan, M.; Valekunja, U.K.; Maywood, E.S.; et al. Peroxiredoxins are conserved markers of circadian rhythms. Nature 2012, 485, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Feeney, K.A.; Hansen, L.L.; Putker, M.; Olivares-yañez, C.; Day, J.; Eades, L.J.; Larrondo, L.F.; Neill, J.S.O.; Van Ooijen, G. Daily magnesium fluxes regulate cellular timekeeping and energy balance. Nature 2016, 532, 375–379. [Google Scholar] [CrossRef] [PubMed]
- van Ooijen, G.; O’Neill, J.S. Intracellular magnesium and the rhythms of life. Cell Cycle 2016, 15, 2997–2998. [Google Scholar] [CrossRef] [PubMed]
- Uetani, N.; Hardy, S.; Gravel, S.-P.; Kiessling, S.; Pietrobon, A.; Wong, N.N.; Chénard, V.; Cermakian, N.; St-Pierre, J.; Tremblay, M.L. PRL2 links magnesium flux and sex-dependent circadian metabolic rhythms. JCI Insight 2017, 2, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.P.; Gan, L.; Jensen, G.J. 3-D ultrastructure of O. tauri: Electron cryotomography of an entire Eukaryotic cell. PLoS ONE 2007, 2. [Google Scholar] [CrossRef] [PubMed]
- Corellou, F.; Schwartz, C.; Motta, J.; Djouani-tahri, E.B. Clocks in the green lineag: Comparative Functional analysis of the circadian architecture of the Picoeukaryote ostreococcus. Plant Cell 2009, 21, 3436–3449. [Google Scholar] [CrossRef] [PubMed]
- Derelle, E.; Ferraz, C.; Rombauts, S.; Rouze, P.; Worden, A.Z.; Robbens, S.; Partensky, F.; Degroeve, S.; Echeynie, S.; Cooke, R.; et al. Genome analysis of the smallest free-living eukaryote Ostreococcus tauri unveils many unique features. Proc. Natl. Acad. Sci. USA 2006, 103, 11647–11652. [Google Scholar] [CrossRef]
- Hindle, M.M.; Martin, S.F.; Noordally, Z.B.; van Ooijen, G.; Barrios-Llerena, M.E.; Simpson, T.I.; Le Bihan, T.; Millar, A.J. The reduced kinome of Ostreococcus tauri: Core eukaryotic signalling components in a tractable model species. BMC Genom. 2014, 15. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Dixon, L.E.; Troein, C.; Millar, A.J. Proteasome function is required for biological timing throughout the twenty-four hour cycle. Curr. Biol. 2011, 21, 869–875. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Hindle, M.; Martin, S.F.; Barrios-Llerena, M.; Sanchez, F.; Bouget, F.Y.; O’Neill, J.S.; Le Bihan, T.; Millar, A.J. Functional analysis of casein Kinase 1 in a minimal circadian system. PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef]
- Le Bihan, T.; Hindle, M.; Martin, S.F.; Barrios-Llerena, M.E.; Krahmer, J.; Kis, K.; Millar, A.J.; van Ooijen, G. Label-free quantitative analysis of the casein kinase 2-responsive phosphoproteome of the marine minimal model species Ostreococcus tauri. Proteomics 2015, 15, 4135–4144. [Google Scholar] [CrossRef]
- Troein, C.; Corellou, F.; Dixon, L.E.; van Ooijen, G.; O’Neill, J.S.; Bouget, F.Y.; Millar, A.J. Multiple light inputs to a simple clock circuit allow complex biological rhythms. Plant J. 2011, 66, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, M.L.; Akman, O.E.; van Ooijen, G. Stochastic models of cellular circadian rhythms in plants help to understand the impact of noise on robustness and clock structure. Front Plant Sci. 2014, 5, 1–6. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Knox, K.; Kis, K.; Bouget, F.-Y.; Millar, A.J. Genomic transformation of the picoeukaryote Ostreococcus tauri. J. Vis. Exp. 2012, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.L.; van den Burg, H.A.; van Ooijen, G. Sumoylation contributes to timekeeping and temperature compensation of the plant circadian clock. J. Biol. Rhythms 2017, 32, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Sterck, L.; Billiau, K.; Abeel, T.; Rouzé, P.; Van de Peer, Y. ORCAE: Online resource for community annotation of eukaryotes. Nat. Methods 2012, 9, 1041. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucl. Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Hofmann, K.; Stoffel, W. TMbase-A database of membrane spanning proteins segments. Biol. Chem. Hoppe Seyler 1993, 347, 166. [Google Scholar]
- Monnier, A.; Liverani, S.; Bouvet, R.; Jesson, B.; Smith, J.Q.; Mosser, J.; Corellou, F.; Bouget, F.-Y. Orchestrated transcription of biological processes in the marine picoeukaryote Ostreococcus exposed to light/dark cycles. BMC Genom. 2010, 11, 192. [Google Scholar] [CrossRef] [PubMed]
- Kolisek, M.; Launay, P.; Beck, A.; Sponder, G.; Serafini, N.; Brenkus, M.; Froschauer, E.M.; Martens, H.; Fleig, A.; Schweigel, M. SLC41A1 is a novel mammalian Mg2+ carrier. J. Biol. Chem. 2008, 283, 16235–16247. [Google Scholar] [CrossRef] [PubMed]
- Wabakken, T.; Rian, E.; Kveine, M.; Aasheim, H.C. The human solute carrier SLC41A1 belongs to a novel eukaryotic subfamily with homology to prokaryotic MgtE Mg2+ transporters. Biochem. Biophys. Res. Commun. 2003, 306, 718–724. [Google Scholar] [CrossRef]
- Romani, A.M.P. Intracellular magnesium homeostasis. Magnes Cent. Nerv. Syst. 2011, 512, 13–58. [Google Scholar] [CrossRef]
- Günther, T. Na+ /Mg2+ antiport in non-erythrocyte vertebrate cells. Magnes Res. 2007, 20, 89–99. [Google Scholar] [PubMed]
- Sahni, J.; Nelson, B.; Scharenberg, A.M. SLC41A2 encodes a plasma-membrane Mg2+ transporter. Biochem. J. 2007, 401, 505–513. [Google Scholar] [CrossRef]
- Mastrototaro, L.; Smorodchenko, A.; Aschenbach, J.R.; Kolisek, M.; Sponder, G. Solute carrier 41A3 encodes for a mitochondrial Mg2+ efflux system. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Hattori, M.; Iwase, N.; Furuya, N.; Tanaka, Y.; Tsukazaki, T.; Ishitani, R.; Maguire, M.E.; Ito, K.; Maturana, A.; Nureki, O. Mg2+-dependent gating of bacterial MgtE channel underlies Mg2+ homeostasis. EMBO J. 2009, 28, 3602–3612. [Google Scholar] [CrossRef]
- Kolisek, M.; Nestler, A.; Vormann, J.; Schweigel-Rontgen, M. Human gene SLC41A1 encodes for the Na+/Mg2+ exchanger. Am. J. Physiol. Cell Physiol. 2012, 302, C318–326. [Google Scholar] [CrossRef]
- Payandeh, J.; Pfoh, R.; Pai, E.F. The structure and regulation of magnesium selective ion channels. Biochim. Biophys. Acta Biomembr. 2013, 1828, 2778–2792. [Google Scholar] [CrossRef]
- Bateman, A. The structure of a domain common to archeobacteria and the homocystinuria disease protein. Trends Biochem. Sci. 1997, 0004, 1–2. [Google Scholar] [CrossRef]
- Tomita, A.; Zhang, M.; Jin, F.; Zhuang, W.; Takeda, H.; Maruyama, T.; Osawa, M.; Hashimoto, K.I.; Kawasaki, H.; Ito, K.; et al. ATP-dependent modulation of MgtE in Mg2+ homeostasis. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dalmas, O.; Sandtner, W.; Medovoy, D.; Frezza, L.; Bezanilla, F.; Perozo, E. A repulsion mechanism explains magnesium permeation and selectivity in CorA. Proc. Natl. Acad. Sci. USA 2014, 111. [Google Scholar] [CrossRef] [PubMed]
- Knoop, V.; Groth-Malonek, M.; Gebert, M.; Eifler, K.; Weyand, K. Transport of magnesium and other divalent cations: Evolution of the 2-TM-GxN proteins in the MIT superfamily. Mol. Genet Genom. 2005, 274, 205–216. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feord, H.K.; Dear, F.E.G.; Obbard, D.J.; van Ooijen, G. A Magnesium Transport Protein Related to Mammalian SLC41 and Bacterial MgtE Contributes to Circadian Timekeeping in a Unicellular Green Alga. Genes 2019, 10, 158. https://doi.org/10.3390/genes10020158
Feord HK, Dear FEG, Obbard DJ, van Ooijen G. A Magnesium Transport Protein Related to Mammalian SLC41 and Bacterial MgtE Contributes to Circadian Timekeeping in a Unicellular Green Alga. Genes. 2019; 10(2):158. https://doi.org/10.3390/genes10020158
Chicago/Turabian StyleFeord, Helen K., Frederick E.G. Dear, Darren J. Obbard, and Gerben van Ooijen. 2019. "A Magnesium Transport Protein Related to Mammalian SLC41 and Bacterial MgtE Contributes to Circadian Timekeeping in a Unicellular Green Alga" Genes 10, no. 2: 158. https://doi.org/10.3390/genes10020158
APA StyleFeord, H. K., Dear, F. E. G., Obbard, D. J., & van Ooijen, G. (2019). A Magnesium Transport Protein Related to Mammalian SLC41 and Bacterial MgtE Contributes to Circadian Timekeeping in a Unicellular Green Alga. Genes, 10(2), 158. https://doi.org/10.3390/genes10020158