An Integrative Systems Perspective on Plant Phosphate Research


Abstract
:1. Introduction
2. Global Perspectives
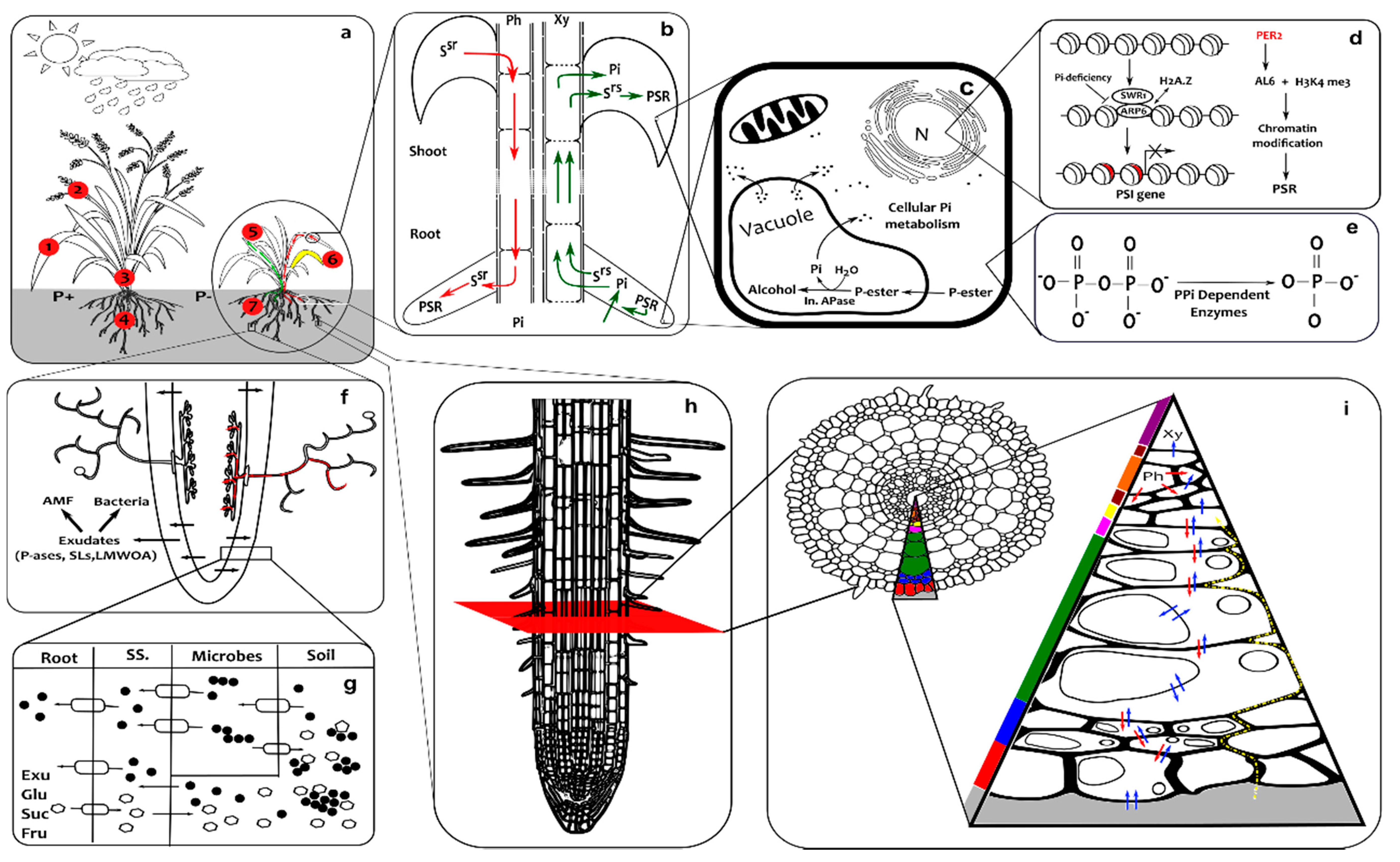
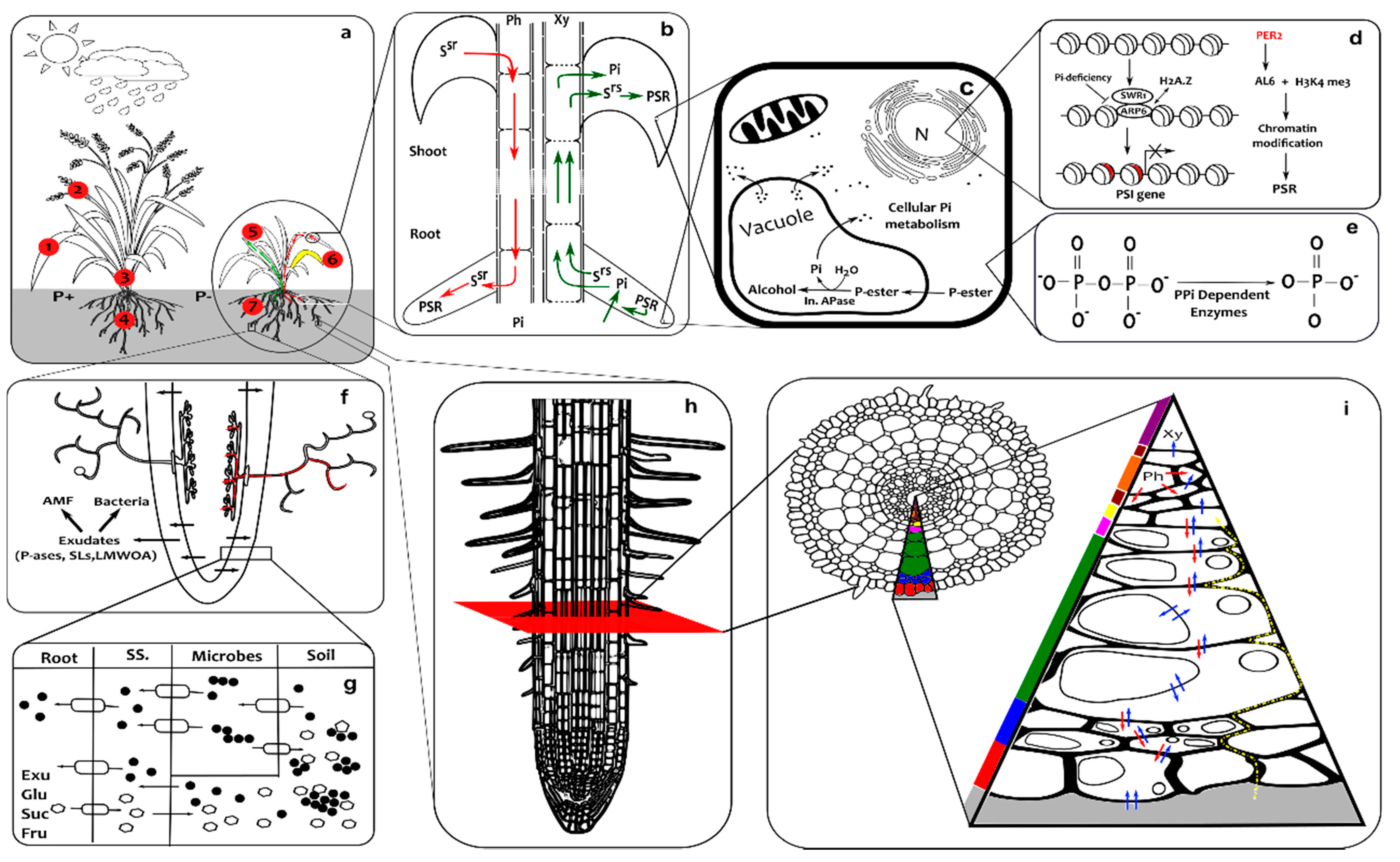
3. Response to Phosphate Starvation
3.1. Shoot
3.2. Root
3.3. Rhizosphere
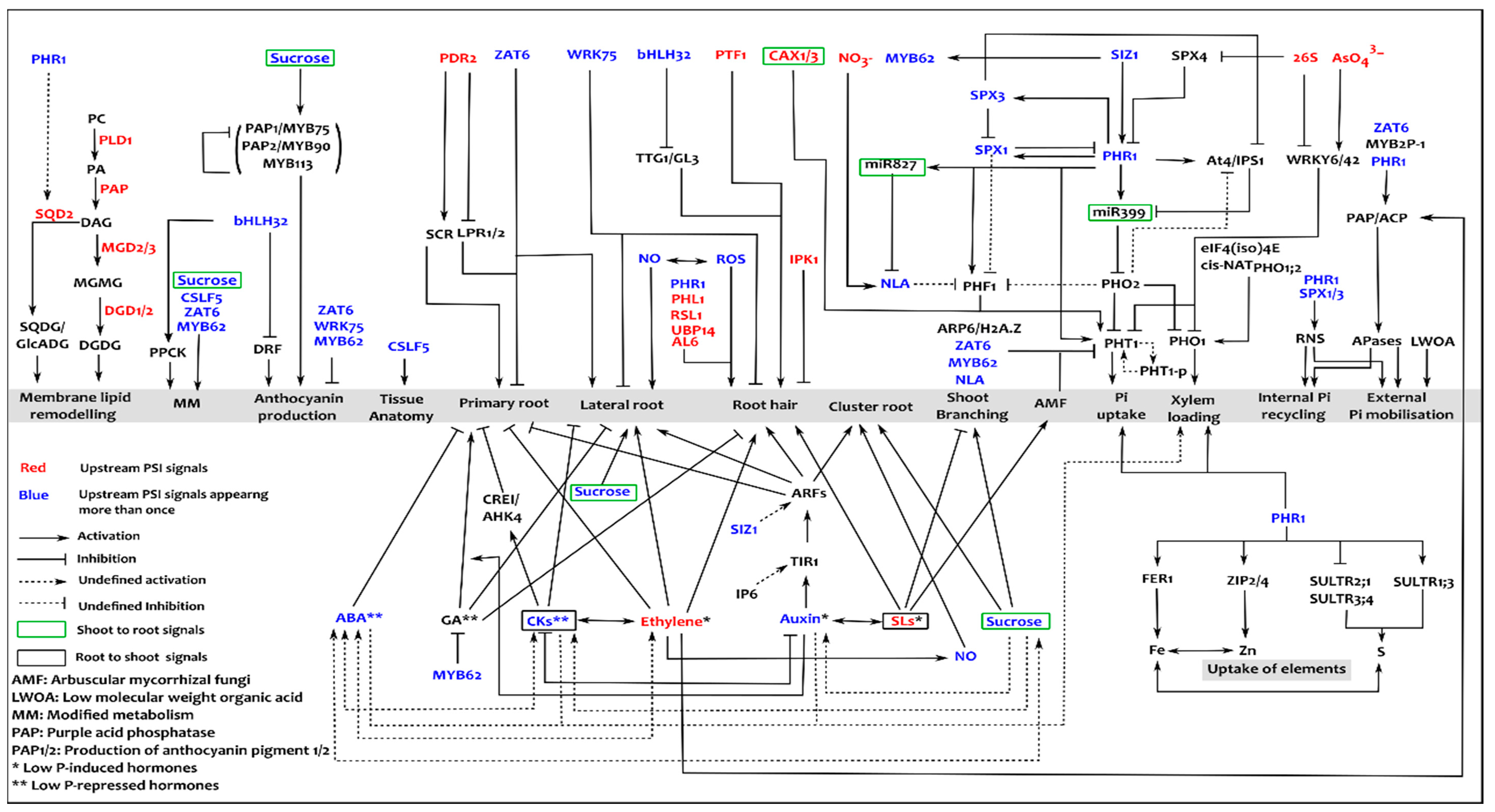
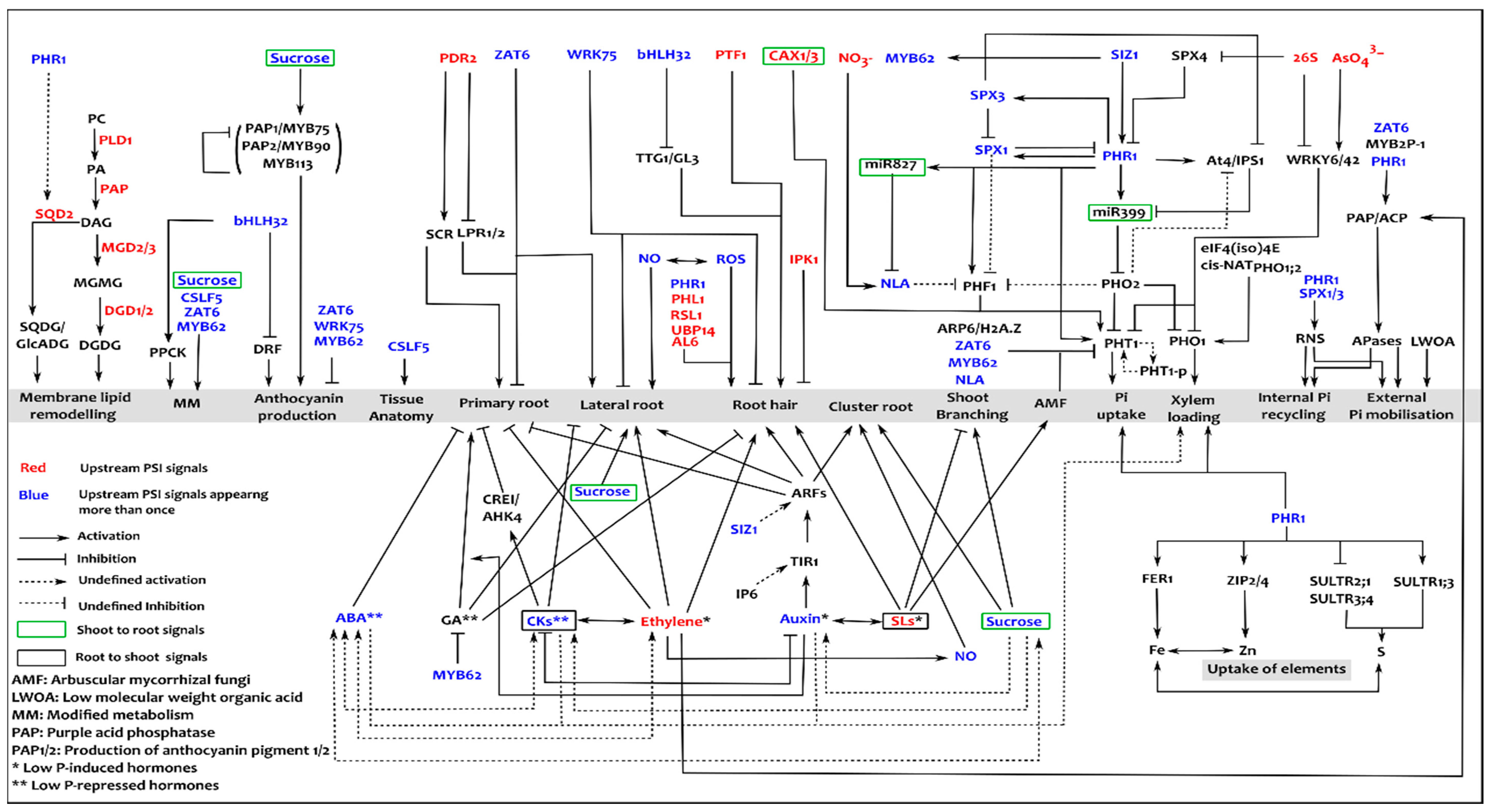
4. Spatio-Temporal Interaction between Phosphate Starvation Responses (PSRs)
5. Current Strategies and Challenges Towards Improving PUE in Plants
6. Need for Integrative Systems Approaches
7. Current Systems Activities in Plant P Research
8. Next Steps
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Gregory, P.J.; George, T.S. Feeding nine billion: The challenge to sustainable crop production. J. Exp. Bot. 2011, 62, 5233–5239. [Google Scholar] [CrossRef]
- Ray, D.K.; Ramankutty, N.; Mueller, N.D.; West, P.C.; Foley, J.A. Recent patterns of crop yield growth and stagnation. Nat. Commun. 2012, 3, 1293. [Google Scholar] [CrossRef]
- Bruinsma, J. World Agriculture: Towards 2015/2030: An FAO Perspective; Routledge: London, UK, 2017. [Google Scholar]
- Gilbert, N. Environment: The disappearing nutrient. Nat. News 2009, 461, 716–718. [Google Scholar] [CrossRef]
- Fixen, P.E.; Johnston, A.M. World fertilizer nutrient reserves: A view to the future. J. Sci. Food Agric. 2012, 92, 1001–1005. [Google Scholar] [CrossRef]
- Johnston, A.E.; Poulton, P.R.; Fixen, P.E.; Curtin, D. Phosphorus: Its efficient use in agriculture. In Advances in Agronomy; Elsevier: Oxford, UK, 2014; Volume 123, Chapter 5; pp. 177–228. [Google Scholar]
- Elser, J.J. Phosphorus: A limiting nutrient for humanity? Curr. Opin. Biotechnol. 2012, 23, 833–838. [Google Scholar] [CrossRef]
- Johnston, A.; Steen, I. Understanding Phosphorus and Its Use in Agriculture; European Fertilizer Manufacturers Association: Brussels, Belgium, 2000. [Google Scholar]
- Syers, J.; Johnston, A.; Curtin, D. Efficiency of Soil and Fertiliser Phosphorus Use: Reconciling Changing Concepts of Soil Phosphorus Behaviour with Agronomic Information; Food and Agriculture Organization of the United Nations: Rome, Italy, 2008. [Google Scholar]
- Faucon, M.-P.; Houben, D.; Reynoird, J.-P.; Mercadal-Dulaurent, A.-M.; Armand, R.; Lambers, H. Advances and perspectives to improve the phosphorus availability in cropping systems for agroecological phosphorus management. In Advances in Agronomy; Elsevier: Oxford, UK, 2015; Volume 134, pp. 51–79. [Google Scholar]
- Gaxiola, R.A.; Edwards, M.; Elser, J.J. A transgenic approach to enhance phosphorus use efficiency in crops as part of a comprehensive strategy for sustainable agriculture. Chemosphere 2011, 84, 840–845. [Google Scholar] [CrossRef]
- Fischer, R.A.; Connor, D.J. Issues for cropping and agricultural science in the next 20 years. Field Crop. Res. 2018, 222, 121–142. [Google Scholar] [CrossRef]
- Maathuis, F.J. Physiological functions of mineral macronutrients. Curr. Opin. Plant Boil. 2009, 12, 250–258. [Google Scholar] [CrossRef]
- Mehra, P.; Pandey, B.K.; Verma, L.; Giri, J. A Novel Glycerophosphodiester Phosphodiesterase Improves Phosphate Deficiency Tolerance. Plant Cell Environ. 2018. [Google Scholar] [CrossRef] [PubMed]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Pierre, W.; Parker, F. Soil phosphorus studies: II. The concentration of organic and inorganic phosphorus in the soil solution and soil extracts and the availability of the organic phosphorus to plants. Soil Sci. 1927, 24, 119–128. [Google Scholar] [CrossRef]
- Raghothama, K. Phosphate acquisition. Annu. Rev. Plant Boil. 1999, 50, 665–693. [Google Scholar] [CrossRef] [PubMed]
- Pratt, J.; Boisson, A.-M.; Gout, E.; Bligny, R.; Douce, R.; Aubert, S. Phosphate (Pi) starvation effect on the cytosolic Pi concentration and Pi exchanges across the tonoplast in plant cells: An in vivo 31P-nuclear magnetic resonance study using methylphosphonate as a Pi analog. Plant Physiol. 2009, 151, 1646–1657. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Plaxton, W.C. Phosphorus: Back to the Roots. Annu. Plant Rev. 2015, 48, 3–22. [Google Scholar]
- Mehra, P.; Pandey, B.; Giri, J. Improvement in phosphate acquisition and utilization by a secretory purple acid phosphatase (OsPAP21b) in rice. Plant Biotechnol. J. 2017, 15, 1054–1067. [Google Scholar] [CrossRef]
- Foyer, C.; Spencer, C. The relationship between phosphate status and photosynthesis in leaves. Planta 1986, 167, 369–375. [Google Scholar] [CrossRef]
- Kennelly, M.; O’Mara, J.; Rivard, C.; Miller, G.L.; Smith, D. Introduction to abiotic disorders in plants. Plant Health Instr. 2012, 10, 1094. [Google Scholar] [CrossRef]
- Hernández, I.; Munné-Bosch, S. Linking phosphorus availability with photo-oxidative stress in plants. J. Exp. Bot. 2015, 66, 2889–2900. [Google Scholar] [CrossRef]
- Smith, F.W. The phosphate uptake mechanism. In Food Security in Nutrient-Stressed Environments: Exploiting Plants’ Genetic Capabilities; Springer: Dordrecht, The Netherlands, 2002; pp. 235–244. [Google Scholar]
- Jeschke, W.D.; Kirkby, E.A.; Peuke, A.D.; Pate, J.S.; Hartung, W. Effects of P deficiency on assimilation and transport of nitrate and phosphate in intact plants of castor bean (Ricinus communis L.). J. Exp. Bot. 1997, 48, 75–91. [Google Scholar] [CrossRef]
- Jouhet, J.; Maréchal, E.; Baldan, B.; Bligny, R.; Joyard, J.; Block, M.A. Phosphate deprivation induces transfer of DGDG galactolipid from chloroplast to mitochondria. J. Cell Biol. 2004, 167, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liao, H.; Lucas, W.J. Molecular mechanisms underlying phosphate sensing, signaling, and adaptation in plants. J. Integr. Plant Boil. 2014, 56, 192–220. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.P.; Bennett, M.J.; Bowen, H.C.; Broadley, M.R.; Eastwood, D.C.; May, S.T.; Clive, R.; Ranjan, S.; Woolaway, K.E.; White, P.J. Changes in gene expression in Arabidopsis shoots during phosphate starvation and the potential for developing smart plants. Plant Physiol. 2003, 132, 578–596. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.K.; Mehra, P.; Verma, L.; Bhadouria, J.; Giri, J. OsHAD1, a Haloacid Dehalogenase-Like APase, Enhances Phosphate Accumulation. Plant Physiol. 2017, 174, 2316–2332. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2013, 112, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Benjamin, P.; Mathilde, C.; Laurent, N.; Thierry, D. Root developmental adaptation to phosphate starvation: Better safe than sorry. Trends Plant Sci. 2011, 16, 442–450. [Google Scholar]
- Haling, R.E.; Brown, L.K.; Stefanski, A.; Kidd, D.R.; Ryan, M.H.; Sandral, G.A.; George, T.S.; Lambers, H.; Simpson, R.J. Differences in nutrient foraging among Trifolium subterraneum cultivars deliver improved P-acquisition efficiency. Plant Soil 2018, 424, 539–554. [Google Scholar] [CrossRef]
- Jung, J.K.H.; Susan, M.C. Getting to the roots of it: Genetic and hormonal control of root architecture. Front. Plant Sci. 2013, 4, 186. [Google Scholar] [CrossRef]
- Lai, F.; Jennifer, T.; Li, Y.; Peter, D. Cell division activity determines the magnitude of phosphate starvation responses in Arabidopsis. Plant J. 2007, 50, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Ticconi, C.A.; Lucero, R.D.; Sakhonwasee, S.; Adamson, A.W.; Creff, A.; Nussaume, L.; Desnos, T.; Abel, S.; Amasino, R.M. ER-Resident Proteins PDR2 and LPR1 Mediate the Developmental Response of Root Meristems to Phosphate Availability. Proc. Natl. Acad. Sci. USA 2009, 106, 14174–14179. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Calderón, L.; López-Bucio, J.; Chacón-López, A.; Cruz-Ramírez, A.; Nieto-Jacobo, F.; Dubrovsky, J.G.; Herrera-Estrella, L. Phosphate starvation induces a determinate developmental program in the roots of Arabidopsis thaliana. Plant Cell Physiol. 2005, 46, 174–184. [Google Scholar] [CrossRef] [PubMed]
- López-Bucio, J.; Hernández-Abreu, E.; Sánchez-Calderón, L.; Nieto Jacobo, M.; Simpson, J.; Herrera-Estrella, L. Phosphate availability alters architecture and causes changes in hormone sensitivity in the Arabidopsis root system. Plant Physiol. 2002, 129, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Williamson, L.C.; Ribrioux, S.P.C.P.; Fitter, A.H.; Leyser, H.M.O. Phosphate availability regulates root system architecture in Arabidopsis. Plant Physiol. 2001, 126, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Liang, W.; Sturrock, C.J.; Pandey, B.K.; Giri, J.; Mairhofer, S.; Wang, D.; Muller, L.; Tan, H.; York, L.M. Rice actin binding protein RMD controls crown root angle in response to external phosphate. Nat. Commun. 2018, 9, 2346. [Google Scholar] [CrossRef]
- Schiefelbein, J.W.; Somerville, C. Genetic control of root hair development in Arabidopsis thaliana. Plant Cell 1990, 2, 235–243. [Google Scholar] [CrossRef]
- Foreman, J.; Dolan, L. Root Hairs as a Model System for Studying Plant Cell Growth. Ann. Bot. 2001, 88, 1–7. [Google Scholar] [CrossRef]
- Ma, Z.; DG, B.; Brown, K.M.; Lynch, J.P. Regulation of root hair density by phosphorus availability in Arabidopsis thaliana. Plant Cell Environ. 2001, 24, 459–467. [Google Scholar] [CrossRef]
- Bhosale, R.; Giri, J.; Pandey, B.K. A mechanistic framework for auxin dependent Arabidopsis root hair elongation to low external phosphate. Nat. Commun. 2018, 9, 1409. [Google Scholar] [CrossRef]
- Giri, J.; Bhosale, R.; Huang, G.; Pandey, B.; Parker, H.; Zappala, S.; Yang, J.; Dievart, A.; Bureau, C.; Ljung, K.; et al. Rice auxin influx carrier OsAUX1 facilitates root hair elongation in response to low external phosphate. Nat. Commun. 2018, 9, 1408. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.R.; Lynch, J.P. The efficiency of Arabidopsis thaliana (Brassicaceae) root hairs in phosphorus acquisition. Am. J. Bot. 2000, 87, 964–970. [Google Scholar] [CrossRef] [PubMed]
- Shane, M.W.; Lambers, H. Cluster Roots: A Curiosity in Context. Plant Soil 2005, 274, 101–125. [Google Scholar] [CrossRef]
- He, C.J.; Morgan, P.W.; Drew, M.C. Enhanced sensitivity to ethylene in nitrogen- or phosphate-starved roots of Zea mays L. during aerenchyma formation. Plant Physiol. 1992, 98, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Baskin, T.; Brown, K.; Lynch, J. Regulation of root elongation under phosphorus stress involves changes in ethylene responsiveness. Plant Physiol. 2003, 131, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Javot, H.; Pumplin, N.; Harrison, M.J. Phosphate in the arbuscular mycorrhizal symbiosis: transport properties and regulatory roles. Plant Cell Environ. 2007, 30, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Dakora, F.D.; Phillips, D.A. Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant Soil 2002, 245, 201–213. [Google Scholar] [CrossRef]
- Tomscha, J.L.; Trull, M.C.; Jill, D.; Lynch, J.P.; Guiltinan, M.J. Phosphatase under-producer mutants have altered phosphorus relations. Plant Physiol. 2004, 135, 334–345. [Google Scholar] [CrossRef]
- McNear, M.N., Jr. The Rhizosphere—Roots, Soil and Everything In Between. Nat. Educ. Knowl. 2003, 4, 1. [Google Scholar]
- Czarnecki, O.; Yang, J.; Weston, D.; Tuskan, G.; Chen, J.-G. A dual role of strigolactones in phosphate acquisition and utilization in plants. Int. J. Mol. Sci. 2013, 14, 7681–7701. [Google Scholar] [CrossRef]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Hayashi, H. Strigolactones: Chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann. Bot. 2006, 97, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Besserer, A.; Puech-Pages, V.; Kiefer, P.; Gomez-Roldan, V.; Jauneau, A.; Roy, S.; Portais, J.C.; Roux, C.; Becard, G.; Sejalon-Delmas, N. Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol. 2006, 4, e226. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, P.F.; Robson, A.D.; Barrow, N.J. Root hair length determines beneficial effect of a Glomus species on shoot growth of some pasture species. New Phytol. 1995, 131, 247–254. [Google Scholar] [CrossRef]
- Chen, W.; Li, J.; Zhu, H.; Xu, P.; Chen, J.; Yao, Q. Arbuscular Mycorrhizal Fungus Enhances Lateral Root Formation in Poncirus trifoliata (L.) as Revealed by RNA-Seq Analysis. Front. Plant Sci. 2017, 8, 2039. [Google Scholar] [CrossRef] [PubMed]
- George, E.; Marschner, H.; Jakobsen, I. Role of arbuscular mycorrhizal fungi in uptake of phosphorus and nitrogen from soil. Crit. Rev. Biotechnol. 1995, 15, 257–270. [Google Scholar] [CrossRef]
- Gutjahr, C.; Paszkowski, U. Multiple control levels of root system remodeling in arbuscular mycorrhizal symbiosis. Front. Plant Sci. 2013, 4, 204. [Google Scholar] [CrossRef] [PubMed]
- Svistoonoff, S.; Creff, A.; Reymond, M.; Sigoillot-Claude, C.; Ricaud, L.; Blanchet, A.; Nussaume, L.; Desnos, T. Root tip contact with low-phosphate media reprograms plant root architecture. Nat. Genet. 2007, 39, 792–796. [Google Scholar] [CrossRef]
- Bonnot, C.; Pinson, B.; Clément, M.; Bernillon, S.; Chiarenza, S.; Kanno, S.; Kobayashi, N.; Delannoy, E.; Nakanishi, T.M.; Nussaume, L.; et al. A chemical genetic strategy identify the PHOSTIN, a synthetic molecule that triggers phosphate starvation responses in Arabidopsis thaliana. New Phytol. 2016, 209, 161–176. [Google Scholar] [CrossRef]
- Chiou, T.-J.; Lin, S.-I. Signaling network in sensing phosphate availability in plants. Annu. Rev. Plant Boil. 2011, 62, 185–206. [Google Scholar] [CrossRef]
- Lin, W.Y.; Huang, T.K.; Leong, S.J.; Chiou, T.J. Long-distance call from phosphate: Systemic regulation of phosphate starvation responses. J. Exp. Bot. 2014, 65, 1817–1827. [Google Scholar] [CrossRef] [PubMed]
- Panigrahy, M.; Rao, D.N.; Sarla, N. Molecular mechanisms in response to phosphate starvation in rice. Biotechnol. Adv. 2009, 27, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.I.; Chiou, T.J. Long-distance movement and differential targeting of microRNA399s. Plant Signal. Behav. 2008, 3, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.Y.; Lin, S.T. Molecular regulators of phosphate homeostasis in plants. J. Exp. Bot. 2009, 60, 1427–1438. [Google Scholar] [CrossRef]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Pazares, J. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes Dev. 2001, 15, 2122–2133. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Rus, A.; Sharkhuu, A.; Yokoi, S.; Karthikeyan, A.S.; Raghothama, K.G.; Baek, D.; Koo, Y.D.; Jin, J.B.; Bressan, R.A.; et al. The Arabidopsis SUMO E3 ligase SIZ1 controls phosphate deficiency responses. Proc. Natl. Acad. Sci. USA 2005, 102, 7760–7765. [Google Scholar] [CrossRef] [PubMed]
- Qundan, L.; Zhong, Y.; Yuguang, W.; Wang, Z.; Li, Z.; Shi, J.; Wu, Z.; Yu, L.; Mao, C.; Yi, K.; et al. SPX4 negatively regulates phosphate signaling and homeostasis through its interaction with PHR2 in rice. Plant Cell 2014, 26, 1586–1597. [Google Scholar]
- María Isabel, P.; Isabel, M.; Rajulu, C.; Zhiye, W.; Franco-Zorrilla, J.M.; Laura, D.L.; Irigoyen, M.L.; Simona, M.; Regla, B.; José, R. SPX1 is a phosphate-dependent inhibitor of Phosphate Starvation Response 1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 14947–14952. [Google Scholar]
- Liu, T.-Y.; Wei-Yi, L.; Teng-Kuei, H.; Tzyy-Jen, C. MicroRNA-mediated surveillance of phosphate transporters on the move. Trends Plant Sci. 2014, 19, 647–655. [Google Scholar] [CrossRef]
- Briat, J.F.; Rouached, H.; Tissot, N.; Gaymard, F.; Dubos, C. Integration of P, S, Fe, and Zn nutrition signals in Arabidopsis thaliana: Potential involvement of PHOSPHATE STARVATION RESPONSE 1 (PHR1). Front. Plant Sci. 2015, 6, 125–132. [Google Scholar] [CrossRef]
- Pant, B.D.; Burgos, A.; Pant, P.; Cuadrosinostroza, A.; Willmitzer, L.; Scheible, W. The transcription factor PHR1 regulates lipid remodeling and triacylglycerol accumulation in Arabidopsis thaliana during phosphorus starvation. J. Exp. Bot. 2015, 66, 1907–1918. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C.; Lambers, H. ’Omics’ Approaches Towards Understanding Plant Phosphorus Acquisition and Use; John Wiley & Sons, Inc.: Chichester, UK, 2015; Chapter 3; pp. 65–97. [Google Scholar]
- Plaxton, W.C.; Shane, M.W. The Role of Post-Translational Enzyme Modifications in the Metabolic Adaptations of Phosphorus-Deprived Plants. In Annual Plant Reviews; John Wiley & Sons, Inc.: Chichester, UK, 2015; Volume 48, Chapter 4; pp. 99–124. [Google Scholar]
- Secco, D.; Wang, C.; Shou, H.; Schultz, M.D.; Chiarenza, S.; Nussaume, L.; Ecker, J.R.; Whelan, J.; Lister, R. Stress induced gene expression drives transient DNA methylation changes at adjacent repetitive elements. Elife 2015, 4, e09343. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Mehdi, J.; Hayden, W.; Huixia, S.; Ping, W.; Yves, P.; James, W. Spatio-temporal transcript profiling of rice roots and shoots in response to phosphate starvation and recovery. Plant Signal. Behav. 2013, 25, 4285–4304. [Google Scholar]
- Yong-Villalobos, L.; Cervantes-Pérez, S.A.; Gutiérrez-Alanis, D.; Gonzáles-Morales, S.; Martínez, O.; Herrera-Estrella, L. Phosphate starvation induces DNA methylation in the vicinity of cis-acting elements known to regulate the expression of phosphate-responsive genes. Plant Signal. Behav. 2016, 11, e1173300. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C.; Tran, H.T. Metabolic adaptations of phosphate-starved plants. Plant Physiol. 2011, 156, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C.; Lambers, H. Metabolomics of Plant Phosphorus-Starvation Response. In Annual Plant Reviews; John Wiley & Sons, Inc.: Chichester, UK, 2015; Volume 48, Chapter 8; pp. 215–236. [Google Scholar]
- Tudzynski, B. Plant Responses to Environmental Stresses: From Phytohormones to Genome Reorganization: H.R. Lerner (Ed.). Marcel Dekker, New York, Basel, 1999, 730 pp., ISBN 0-8247-0044-9. Phytochemistry 2001, 56, 641–642. [Google Scholar] [CrossRef]
- Morcuende, R.; Bari, R.; Gibon, Y.; Zheng, W.; Pant, B.D.; Bl Sing, O.; Usadel, B.R.; Czechowski, T.; Udvardi, M.K.; Stitt, M. Genome-wide reprogramming of metabolism and regulatory networks of Arabidopsis in response to phosphorus. Plant Cell Environ. 2010, 30, 85–112. [Google Scholar] [CrossRef]
- Pant, B.D.; Pant, P.; Erban, A.; Huhman, D.; Kopka, J.; Scheible, W.R. Identification of primary and secondary metabolites with phosphorus status-dependent abundance in Arabidopsis, and of the transcription factor PHR1 as a major regulator of metabolic changes during phosphorus limitation. Plant Cell Environ. 2015, 38, 172–187. [Google Scholar] [CrossRef]
- ricexPro.dna.affrc.go.jp. Available online: http://ricexpro.dna.affrc.go.jp/RXP_4001/index.php (accessed on 11 September 2019).
- Hammond, J.P.; White, P.J. Sucrose transport in the phloem: Integrating root responses to phosphorus starvation. J. Exp. Bot. 2008, 59, 93. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Solfanelli, C.; Perata, P. Sucrose-Specific Induction of the Anthocyanin Biosynthetic Pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, A.; Varadarajan, D.; Jain, A.; Held, M.; Carpita, N.; Raghothama, K. Phosphate starvation responses are mediated by sugar signaling in Arabidopsis. Planta 2007, 225, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.P.; Broadley, M.R.; White, P.J. Genetic responses to phosphorus deficiency. Ann. Bot. 2004, 94, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.C.; Zakhleniuk, O.V. Responses of primary and secondary metabolism to sugar accumulation revealed by microarray expression analysis of the Arabidopsis mutant, pho3. J. Exp. Bot. 2004, 55, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Müller, R.; Morant, M.; Jarmer, H.; Nilsson, L.; Nielsen, T.H. Genome-Wide Analysis of the Arabidopsis Leaf Transcriptome Reveals Interaction of Phosphate and Sugar Metabolism. Plant Physiol. 2007, 143, 156–171. [Google Scholar] [CrossRef]
- Ticconi, C.A.; Abel, S. Short on phosphate: Plant surveillance and countermeasures. Trends Plant Sci. 2004, 9, 548–555. [Google Scholar] [CrossRef]
- Hernández, G.; Oswaldo, V.L.; Mario, R.; Nicolas, G.; Georg, W.; Rosaura, A.F.; Sara Isabel, F.; Alexander, E.; Joachim, K.; Udvardi, M.K. Global changes in the transcript and metabolic profiles during symbiotic nitrogen fixation in phosphorus-stressed common bean plants. Plant Physiol. 2009, 151, 1221–1238. [Google Scholar] [CrossRef]
- Obata, T.; Fernie, A.R. The use of metabolomics to dissect plant responses to abiotic stresses. Cell. Mol. Life Sci. 2012, 69, 3225–3243. [Google Scholar] [CrossRef]
- Lahner, B.; Gong, J.M.; Smith, E.L.; Abid, K.B.; Rogers, E.E. Genomic scale profiling of nutrient and trace elements in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 1215–1221. [Google Scholar] [CrossRef]
- Salt, D.E. Update on plant ionomics. Plant Physiol. 2004, 136, 2451–2456. [Google Scholar] [CrossRef]
- Baxter, I. Ionomics: Studying the social network of mineral nutrients. Curr. Opin. Plant Biol. 2009, 12, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Baxter, I.; Springer, N.; Jackson, S. Ionomics: The functional genomics of elements. Brief. Funct. Genom. 2010, 9, 149. [Google Scholar] [CrossRef]
- Conn, S.; Gilliham, M. Comparative physiology of elemental distributions in plants. Ann. Bot. 2010, 105, 1081–1102. [Google Scholar] [CrossRef] [PubMed]
- Baxter, I.R.; Vitek, O.; Lahner, B.; Muthukumar, B.; Borghi, M.; Morrissey, J.; Guerinot, M.L.; Salt, D.E. The leaf ionome as a multivariable system to detect a plant’s physiological status. Proc. Natl. Acad. Sci. USA 2008, 105, 12081–12086. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, U.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Weber, A.P.; Zellerhoff, N.; Bucher, M.; Fahnenstich, H.; Sonnewald, U. Adaptation of maize source leaf metabolism to stress related disturbances in carbon, nitrogen and phosphorus balance. BMC Genom. 2013, 14, 442. [Google Scholar] [CrossRef] [PubMed]
- Naoko, O.O.; Jun, W. Recent progress in plant nutrition research: Cross-talk between nutrients, plant physiology and soil microorganisms. Plant Cell Physiol. 2010, 51, 1255–1264. [Google Scholar]
- Baxter, I.; Dilkes, B.P. Elemental profiles reflect plant adaptations to the environment. Science 2012, 336, 1661. [Google Scholar] [CrossRef]
- Dai, X.; Wang, Y.; Zhang, W.-H. OsWRKY74, a WRKY transcription factor, modulates tolerance to phosphate starvation in rice. J. Exp. Bot. 2015, 67, 947–960. [Google Scholar] [CrossRef]
- Kawa, D.; Julkowska, M.; Montero Sommerfeld, H.; ter Horst, A.; Haring, M.A.; Testerink, C. Phosphate-dependent root system architecture responses to salt stress. Plant Physiol. 2016, 172. [Google Scholar] [CrossRef]
- Wild, R.; Gerasimaite, R.; Jung, J.Y.; Truffault, V.; Pavlovic, I.; Schmidt, A.; Saiardi, A.; Jessen, H.J.; Poirier, Y.; Hothorn, M. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 2016, 352, 986–990. [Google Scholar] [CrossRef]
- Yue, W.; Ying, Y.; Wang, C.; Zhao, Y.; Dong, C.; Whelan, J.; Shou, H. OsNLA1, a RING-type ubiquitin ligase, maintains phosphate homeostasis in Oryza sativa via degradation of phosphate transporters. Plant J. Cell Mol. Boil. 2017, 90, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.J.; Stefanski, A.; Marshall, D.J.; Moore, A.D.; Richardson, A.E. Management of soil phosphorus fertility determines the phosphorus budget of a temperate grazing system and is the key to improving phosphorus efficiency. Agric. Ecosyst. Environ. 2015, 212, 263–277. [Google Scholar] [CrossRef]
- Jia, X.; Liu, P.; Lynch, J.P. Greater lateral root branching density in maize improves phosphorus acquisition from low phosphorus soil. J. Exp. Bot. 2018, 69, 4961–4970. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Castaneda, T.; Brown, K.M.; Lynch, J.P. Reduced root cortical burden improves growth and grain yield under low phosphorus availability in maize. Plant Cell Environ. 2018, 41, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Strock, C.F.; Morrow de la Riva, L.; Lynch, J.P. Reduction in Root Secondary Growth as a Strategy for Phosphorus Acquisition. Plant Physiol. 2018, 176, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Rose, T.; Liu, L.; Wissuwa, M. Improving phosphorus efficiency in cereal crops: Is breeding for reduced grain phosphorus concentration part of the solution? Front. Plant Sci. 2013, 4, 444. [Google Scholar] [CrossRef]
- Ha, S.; Tran, L.S. Understanding plant responses to phosphorus starvation for improvement of plant tolerance to phosphorus deficiency by biotechnological approaches. Crit. Rev. Biotechnol. 2014, 34, 16–30. [Google Scholar] [CrossRef]
- Lópezarredondo, D.L.; Leyvagonzález, M.A.; Gonzálezmorales, S.I.; Lópezbucio, J.; Herreraestrella, L. Phosphate Nutrition: Improving Low-Phosphate Tolerance in Crops. Annu. Rev. Plant Boil. 2014, 65, 95–123. [Google Scholar] [CrossRef]
- Lambers, H.; Finnegan, P.M.; Jost, R.; Plaxton, W.C.; Shane, M.W.; Stitt, M. Phosphorus nutrition in Proteaceae and beyond. Nat. Plants 2015, 1, 15109. [Google Scholar] [CrossRef]
- Brown, L.K.; George, T.S.; Dupuy, L.X.; White, P.J. A conceptual model of root hair ideotypes for future agricultural environments: What combination of traits should be targeted to cope with limited P availability? Ann. Bot. 2013, 112, 317–330. [Google Scholar] [CrossRef]
- Scheible, W.R.; Rojas-Triana, M. Sensing, Signaling and Control of Phosphate Starvation in Plants: Molecular Players and Applications. In Annual Plant Reviews; John Wiley & Sons, Inc.: Chichester, UK, 2015; Volume 48, Chapter 2; pp. 23–63. [Google Scholar]
- Vance, C.P. Quantitative trait loci, epigenetics, sugars, and microRNAs: Quaternaries in phosphate acquisition and use. Plant Physiol. 2010, 154, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Chin, J.H.; Rico, G.; Cheryl, D.; Masdiar, B.; Joko, P.; Sugiono, M.; Matthias, W.; Sigrid, H. Developing rice with high yield under phosphorus deficiency: Pup1 sequence to application. Plant Physiol. 2011, 156, 1202–1216. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Wissuwa, M.; Wegner, J.; Ae, N.; Yano, M. Substitution mapping of Pup1: A major QTL increasing phosphorus uptake of rice from a phosphorus-deficient soil. Theor. Appl. Genet. 2002, 105, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Gamuyao, R.; Chin, J.H.; Pariascatanaka, J.; Pesaresi, P.; Catausan, S.; Dalid, C.; Slametloedin, I.; Tecsonmendoza, E.M.; Wissuwa, M.; Heuer, S. The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature 2012, 488, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E. Regulating the phosphorus nutrition of plants: Molecular biology meeting agronomic needs. Plant Soil 2009, 322, 17–24. [Google Scholar] [CrossRef]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Boil. 2010, 63, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Ramaekers, L.; Remans, R.; Rao, I.M.; Blair, M.W.; Vanderleyden, J. Strategies for improving phosphorus acquisition efficiency of crop plants. Field Crop. Res. 2010, 117, 169–176. [Google Scholar] [CrossRef]
- Tian, J.; Xiurong, W.; Yiping, T.; Xinping, C.; Hong, L. Bioengineering and management for efficient phosphorus utilization in crops and pastures. Curr. Opin. Biotechnol. 2012, 23, 866–871. [Google Scholar] [CrossRef]
- Pérez-Clemente, R.M.; Vives, V.; Zandalinas, S.I.; López-Climent, M.F.; Muñoz, V.; Gómez-Cadenas, A. Biotechnological approaches to study plant responses to stress. BioMed Res. Int. 2013, 2013, 654120. [Google Scholar] [CrossRef]
- Péret, B.; Li, G.; Zhao, J.; Band, L.R.; Voß, U.; Postaire, O.; Luu, D.T.; Da Ines, O.; Casimiro, I.; Lucas, M.; et al. Auxin regulates aquaporin function to facilitate lateral root emergence. Nat. Cell Biol. 2012, 14, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Marbach, D.; Costello, J.; Küffner, R.; Vega, N.; Prill, R.; Camacho, D.; Allison, K.; Kellis, M.; Collins, J.; Stolovitzky, G. Wisdom of crowds for robust gene network inference. Nat. Methods 2012, 9, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Glyn, B.; Kevin, P.; Graham, B.; Chungui, L.; Rupert, F.; Alexandra, M.; Subhalai, J.; Charles, B.; Rik, V.W. Network inference analysis identifies an APRR2-like gene linked to pigment accumulation in tomato and pepper fruits. Plant Physiol. 2013, 161, 1476–1485. [Google Scholar] [CrossRef] [PubMed]
- Tak, L.; Taeyun, O.; Sunmo, Y.; Junha, S.; Sohyun, H.; Yeong, K.C.; Hyojin, K.; Hongseok, S.; Jung Eun, S.; Ronald, P.C. RiceNet v2: An improved network prioritization server for rice genes. Nucleic Acids Res. 2015, 43, W122. [Google Scholar]
- Lucas, M.; Laplaze, L.; Bennett, M.J. Plant Systems Biology: Network Matters. Plant Cell Environ. 2011, 34, 535–553. [Google Scholar] [CrossRef]
- Lavenus, J.; Middleton, A.; Wilson, M.; Lucas, M.; Laplaze, L.; Bennett, M.; Crespi, M. Toward a Virtual Root: Interaction of Genomics and Modeling to Develop Predictive Biology Approaches. In Root Genomics and Soil Interactions; John Wiley & Sons, Inc.: Chichester, UK, 2012; Chapter 5; pp. 79–92. [Google Scholar]
- Kristine, H.; Silvana, P.; Guillaume, L.; Susan, Z.; Sacha, M.; Xavier, D.; Bennett, M.J. Root Systems Biology: Integrative Modeling across Scales, from Gene Regulatory Networks to the Rhizosphere. Plant Physiol. 2013, 163, 1487–1503. [Google Scholar]
- Hodgman, T.C.; Ajmera, I. The successful application of systems approaches in plant biology. Prog. Biophys. Mol. Biol. 2015, 117, 59–68. [Google Scholar] [CrossRef]
- Tardieu, F.; Tardieu, F. Why work and discuss the basic principles of plant modelling 50 years after the first plant models? J. Exp. Bot. 2010, 61, 2039–2041. [Google Scholar] [CrossRef]
- Idso, S.B. Mathematical Models in Plant Physiology: A Quantitative Approach to Problems in Plant and Crop Physiology: J. H. M. Thornley. Experimental Botany, Vol. 8, Academic Press, London, 1976, 318 pp., £9.80; Academic Press: London, UK, 1978; Volume 19, pp. 59–60. [Google Scholar]
- Ajmera, I.; Shi, J.; Giri, J.; Wu, P.; Stekel, D.J.; Lu, C.; Hodgman, T.C. Regulatory feedback response mechanisms to phosphate starvation in rice. NPJ Syst. Boil. Appl. 2018, 4, 4. [Google Scholar] [CrossRef]
- Roose, T.; Fowler, A.C. A mathematical model for water and nutrient uptake by plant root systems. J. Theor. Boil. 2004, 228, 173–184. [Google Scholar] [CrossRef]
- Leitner, D.; Klepsch, S.; Ptashnyk, M.; Marchant, A.; Kirk, G.J.; Schnepf, A.; Roose, T. A dynamic model of nutrient uptake by root hairs. New Phytol. 2010, 185, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Ptashnyk, M.; Roose, T.; Jones, D.L.; Kirk, G.J. Enhanced zinc uptake by rice through phytosiderophore secretion: A modelling study. Plant Cell Environ. 2011, 34, 2038–2046. [Google Scholar] [CrossRef] [PubMed]
- Zygalakis, K.C.; Kirk, G.J.D.; Jones, D.L.; Wissuwa, M.; Roose, T. A dual porosity model of nutrient uptake by root hairs. New Phytol. 2011, 192, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Zygalakis, K.C.; Roose, T. A mathematical model for investigating the effect of cluster roots on plant nutrient uptake. Eur. Phys. J. Spec. Top. 2012, 204, 103–118. [Google Scholar] [CrossRef]
- Keyes, S.D.; Daly, K.R.; Gostling, N.J.; Jones, D.L.; Talboys, P.; Pinzer, B.R.; Boardman, R.; Sinclair, I.; Marchant, A.; Roose, T. High resolution synchrotron imaging of wheat root hairs growing in soil and image based modelling of phosphate uptake. New Phytol. 2013, 198, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Payvandi, S.; Daly, K.R.; Jones, D.L.; Talboys, P.; Zygalakis, K.C.; Roose, T. A Mathematical Model of Water and Nutrient Transport in Xylem Vessels of a Wheat Plant. Bull. Math. Boil. 2014, 76, 566–596. [Google Scholar] [CrossRef] [PubMed]
- Heppell, J.; Payvandi, S.; Talboys, P.; Zygalakis, K.C.; Fliege, J.; Langton, D.; Sylvester-Bradley, R.; Walker, R.; Jones, D.L.; Roose, T. Modelling the optimal phosphate fertiliser and soil management strategy for crops. Plant Soil 2016, 401, 135–149. [Google Scholar] [CrossRef]
- Andrea, S.; Tiina, R. Modelling the contribution of arbuscular mycorrhizal fungi to plant phosphate uptake. New Phytol. 2010, 171, 669–682. [Google Scholar]
- Schnepf, A.; Roose, T.; Schweiger, P. Impact of growth and uptake patterns of arbuscular mycorrhizal fungi on plant phosphorus uptake—A modelling study. Plant Soil 2008, 312, 85–99. [Google Scholar] [CrossRef]
- Schnepf, A.; Jones, D.; Roose, T. Modelling Nutrient Uptake by Individual Hyphae of Arbuscular Mycorrhizal Fungi: Temporal and Spatial Scales for an Experimental Design. Bull. Math. Biol. 2011, 73, 2175–2200. [Google Scholar] [CrossRef]
- Kirk, G.J.D. Modelling root-induced solubilization of nutrients. Plant Soil 2002, 245, 49–57. [Google Scholar] [CrossRef]
- Barrow, N.J. A mechanistic model for describing the sorption and desorption of phosphate by soil. J. Soil Sci. 1983, 34, 733–750. [Google Scholar] [CrossRef]
- Silberbush, M.; Barber, S.A. Sensitivity of simulated phosphorus uptake to parameters used by a mechanistic-mathematical model. Plant Soil 1983, 74, 93–100. [Google Scholar] [CrossRef]
- Grant, R.F.; Robertson, J.A. Phosphorus uptake by root systems: Mathematical modelling in ecosys. Plant Soil 1997, 188, 279–297. [Google Scholar] [CrossRef]
- Rowell, D.L. Solute Movement in the Rhizosphere. J. Soil Sci. 2010, 52, 521–522. [Google Scholar] [CrossRef]
- Amijee, F.; Barraclouch, P.B.; Tinker, P.B. Modeling phosphorus uptake and utilization by plants. In Phosphorus Nutrition of Grain Legumes in the Semi-Arid Tropics; ICRISAT: Patancheru, India, 1991; pp. 63–75. [Google Scholar]
- Matthias, W. How do plants achieve tolerance to phosphorus deficiency? Small causes with big effects. Plant Physiol. 2003, 133, 1947–1958. [Google Scholar]
- Heppell, J.; Talboys, P.; Payvandi, S.; Zygalakis, K.C.; Fliege, J.; Withers, P.J.; Jones, D.L.; Roose, T. How changing root system architecture can help tackle a reduction in soil phosphate (P) levels for better plant P acquisition. Plant Cell Environ. 2015, 38, 118–128. [Google Scholar] [CrossRef]
- Postma, J.A.; Lynch, J.P. Theoretical evidence for the functional benefit of root cortical aerenchyma in soils with low phosphorus availability. Ann. Bot. 2011, 107, 829–841. [Google Scholar] [CrossRef]
- Richard, H.; Claire, H.; Penfold, C.A.; Emily, B.; Laura, B.; Moore, J.D.; Peijun, Z.; Alison, J.; Emma, C.; Findlay, B.C. A local regulatory network around three NAC transcription factors in stress responses and senescence in Arabidopsis leaves. Plant J. Cell Mol. Biol. 2013, 75, 26–39. [Google Scholar]
- Brown, T.B.; Cheng, R.; Sirault, X.R.R.; Rungrat, T.; Murray, K.D.; Trtilek, M.; Furbank, R.T.; Badger, M.; Pogson, B.J.; Borevitz, J.O. TraitCapture: Genomic and environment modelling of plant phenomic data. Curr. Opin. Plant Biol. 2014, 18, 73–79. [Google Scholar] [CrossRef]
- Veneklaas, E.; Lambers, H.; Bragg, J.; Finnegan, P.; Lovelock, C.; Plaxton, W.; Price, C.; Scheible, W.; Shane, M.; White, P.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Christophe, P.; Samuel, D.; Frédéric, B.; Christian, F.; Christophe, G. OpenAlea: A visual programming and component-based software platform for plant modelling. Funct. Plant Biol. 2008, 35, 751–760. [Google Scholar]
- Merks, R.; Guravage, M.; Inzé, D.; Beemster, G. Virtualleaf: An open-source framework for cell-based modeling of plant tissue growth and development. Plant Physiol. 2011, 155, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Hoon, C.Y.; Bénédicte, W.; Anna, F.; Virginie, M.; Jasper, T.; Davey, C.L.; Christopher, T.; Howard, T.; Ougham, H.J.; Philippe, D.R. Multiscale digital Arabidopsis predicts individual organ and whole-organism growth. Proc. Natl. Acad. Sci. USA 2014, 111, E4127–E4136. [Google Scholar]
- Arnaud, C.; Clement, M.; Thibaud, M.; Javot, H.; Chiarenza, S.; Delannoy, E.; Revol, J.; Soreau, P.; Bo, C.; Block, M.; et al. Identification of Phosphatin, a drug alleviating phosphate starvation responses in Arabidopsis. Plant Physiol. 2014, 166, 1479–1491. [Google Scholar] [CrossRef] [PubMed]
- Ricarda, J.; Made, P.; Lapis-Gaza, H.R.; Claudia, R.; Oliver, B.; Hans, L.; Finnegan, P.M. Differentiating phosphate-dependent and phosphate-independent systemic phosphate-starvation response networks in Arabidopsis thaliana through the application of phosphite. J. Exp. Bot. 2015, 66, 2501–2514. [Google Scholar]
- Fang, Z.; Shao, C.; Meng, Y.; Wu, P.; Chen, M. Phosphate signaling in Arabidopsis and Oryza sativa. Plant Sci. 2009, 176, 170–180. [Google Scholar] [CrossRef]
- Carlos, C.V.; Sawers, R.J.H.; Luis, H.E. Phosphate deprivation in maize: Genetics and genomics. Plant Physiol. 2011, 156, 1067–1077. [Google Scholar]
- plant.ensembl.org. Available online: http://plants.ensembl.org/index.html (accessed on 11 September 2019).
- Calderon-Vazquez, C.; Sawers, J.H.R.; Herrera-Estrella, L. Phosphate Deprivation in Maize: Genetics and Genomics. Plant Physiology. 2011, 156, 1067–1077. [Google Scholar] [CrossRef]
- Mirbase.org. Available online: http://mirbase.org (accessed on 11 September 2019).
- Wilson, M.H.; Holman, T.J.; Sørensen, I.; Canchosanchez, E.; Wells, D.M.; Swarup, R.; Knox, J.P.; Willats, W.G.; Ubedatomás, S.; Holdsworth, M. Multi-omics analysis identifies genes mediating the extension of cell walls in the Arabidopsis thaliana root elongation zone. Front. Cell Dev. Biol. 2015, 3, 10. [Google Scholar] [CrossRef]
- Band, L.R.; Fozard, J.A.; Christophe, G.; Jensen, O.E.; Tony, P.; Bennett, M.J.; King, J.R. Multiscale systems analysis of root growth and development: Modeling beyond the network and cellular scales. Plant Cell 2012, 24, 3892–3906. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Huang, T.K.; Kuo, H.F.; Chiou, T.J. Role of vacuoles in phosphorus storage and remobilization. J. Exp. Bot. 2017, 68, 3045–3055. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Banerjee, S.; Wheeler, A.; Ratliff, L.A.; Irigoyen, S.; Garcia, L.R.; Lockless, S.W.; Versaw, W.K. Live imaging of inorganic phosphate in plants with cellular and subcellular resolution. Plant Physiol. 2015, 167, 628–638. [Google Scholar] [CrossRef] [PubMed]
- De Col, V.; Fuchs, P.; Nietzel, T.; Elsasser, M.; Voon, C.P.; Candeo, A.; Seeliger, I.; Fricker, M.D.; Grefen, C.; Moller, I.M.; et al. ATP sensing in living plant cells reveals tissue gradients and stress dynamics of energy physiology. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S., III. Effects of multiple environmental stresses on nutrient availability and use. In Response of Plants to Multiple Stresses; Academic Press: Cambridge, MA, USA, 1991; Chapter 3; pp. 67–88. [Google Scholar]
- Kenji, M.; Aiko, S.; Masaru, O.; Jun, F. Increased tolerance to salt stress in the phosphate-accumulating Arabidopsis mutants siz1 and pho2. Planta 2011, 234, 1191–1199. [Google Scholar]
- Plaxton, W.C.; Lambers, H. Interactions between Nitrogen and Phosphorus Metabolism. In Annual Plant Reviews; John Wiley & Sons, Inc.: Chichester, UK, 2015; Volume 48, Chapter 7; pp. 187–214. [Google Scholar]
- Giehl, R.F.H.; Gruber, B.D.; Von Wirén, N. It’s time to make changes: Modulation of root system architecture by nutrient signals. J. Exp. Bot. 2014, 65, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Vos, J.; Evers, J.B.; Bucksorlin, G.H.; Andrieu, B.; Chelle, M.; Visser, P.H.B.D.; Tardieu, F. Functional-structural plant modelling: A new versatile tool in crop science. J. Exp. Bot. 2010, 61, 2101–2115. [Google Scholar] [CrossRef] [PubMed]
- Kuijken, R.C.; van Eeuwijk, F.A.; Marcelis, L.F.; Bouwmeester, H.J. Root phenotyping: From component trait in the lab to breeding. J. Exp. Bot. 2015, 66, 5389–5401. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Schneider, H.; Burridge, J.; Ascanio, A.K.M.; Wojciechowski, T.; Topp, C.N.; Lynch, J.P.; Weitz, J.S.; Bucksch, A. Digital imaging of root traits (DIRT): A high-throughput computing and collaboration platform for field-based root phenomics. Plant Methods 2015, 11, 51. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Gene | MSU Id (LOC_Os) | P+ | P-(Whole Root) | ||
|---|---|---|---|---|---|
| EES | Cortex | EPS | |||
| ARP6 | 01g16414 | → | |||
| bHLH32 | 03g15440 | → | |||
| CAX1 | 11g05070 | ↘ | |||
| CAX3 ** | 02g21009 | ↗ | |||
| IPK1 | 04g56580 | → | |||
| IPS1 ** | 03g05334 | ↗ | |||
| LPR1 | 01g03530 | ↘ | |||
| MYB62 ** | 01g03720 | ↗ | |||
| PHF1 | 07g09000 | ↗ | |||
| PHO1 | 02g56510 | ↗ | |||
| PHO2 | 05g48390 | ↘ | |||
| PHR1 | 03g21240 | → | |||
| PHR2 | 07g25710 | → | |||
| PHT1;1 ** | 04g10800 | ↗ | |||
| PHT1;2 | 03g05620 | ↗ | |||
| PHT1;4 | 04g10750 | ↗ | |||
| PHT1;6 | 08g45000 | ↗ | |||
| PHT1;8 | 10g30790 | ↗ | |||
| PHT2;1 ** | 02g38020 | → | |||
| PLD1 | 05g29050 | → | |||
| PTF1 | 06g09370 | → | |||
| SIZ1 | 05g03430 | → | |||
| SPX1 | 06g40120 | ↗ | |||
| SPX3 | 10g25310 | ↗ | |||
| SQD1 | 07g01030 | ↗ | |||
| ZAT6 ** | 03g32230 | ↗ | |||
| Very high | High | Medium | Low | Very low | Nil |
| Gene | Arabidopsis | Rice | Maize | Sorghum | Brachy | Wheat | Barley | Soybean | Tomato | Potato | Brassica | Grape | Medicago | Populus |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prefix | At | LOC_Os | GRMZ2G | Sb | Bradi | Traes_ | MLOC_ | GLYMA | Solyc |
PGSC0003 DMG4000 | Bra0 | VIT_ | MTR_ | POPTR_0 |
| ARP6 | 3g33520 | 01g16414 | 088487 | 08g21780 | 2g10130 | 2BL_221EC0BB1 | 64804 | 04G07540 06G26590 | 05g018600 | 14966 | 30818 | 11s0016g05490 | ND | 018s12840 002s0340 |
| 1BL_51C489EEB | ||||||||||||||
| 2AL_4DCD06BE9 | ||||||||||||||
| 2DL_E17A1BB12 | ||||||||||||||
|
At4, IPS1 | 5g03545 | 03g05334 | 086179 | ND | ND | 4AL_D7C83DB52 | ND | ND | ND | ND | ND | ND | ND | ND |
| 3g09922 | 01g0838350 ** | 843352 | 4BL_00AAC9279 | |||||||||||
| bHLH32 | 3g25710 | 03g15440 01g06640 | 043854 088443 | 01g040450 03g005250 | 1g67500 2g03830 | 4DL_7BD3165FB | 66385 | 12G14400 | 07g053290 | 23577 | 26510 25158 | 10s0003g01160 | 5g066080 | 004s05480 |
| 4AS_DD16F89E7 | ||||||||||||||
| 4BL_A2B32FCEE | ||||||||||||||
| CAX1 | 1g08960 | 11g05070 | 004414 | 08g002860 | 4g42870 4g42880 | 5BL_6A7BE3F0C | 21103 16013 | 03G39920 | 01g098800 | 28607 | 31650 | 14s0128g00240 | 7g113730 | 013s02590 |
| 5AL_97C4C3E20 | ||||||||||||||
| 5DL_B669983C4 | ||||||||||||||
| 5AL_B925FC757 | ||||||||||||||
| CAX3 | 3g51860 | 01g37690 02g21009 | 011592 | 03g024820 04g010130 | 2g41090 | 2AL_26065F906 | 17450 13658 | 01G30610 | 09g005260 | 11787 | 12833 | 08s0007g02240 | 7g068380 4g016720 | 006s10080 016s12290 |
| 01G30643 | ||||||||||||||
| 2BL_445F5B3B7 | 03G07910 | |||||||||||||
| 2DL_2A2F4F62B | 03G07950 | |||||||||||||
| 18G43000 | ||||||||||||||
| 3AL_3CA77B366 | ||||||||||||||
| IPK1 | 5g42810 | 04g56580 | 150496 | 06g31650 | 5g24890 | 2AL_55460AE54 | 38910 | 14G07880 | 04g080670 | 03698 15950 | 27458 | 18s0001g12910 | ND | 005s25180 |
| 2BL_A465A5769 | ||||||||||||||
| 2DL_EFEF40224 | ||||||||||||||
| LPR1 | 1g23010 | 01g03530 01g03549 01g03620 | 086727 054050 | 03g007480 03g007470 03g007440 | 2g01850 | 4DS_DE0D4FCF6 | 79739 | 20G26270 10G41010 | 05g008290 | 30551 | 24558 | 01s0011g04720 | ND | 010s12440 |
| 4BS_ECDA94252 | ||||||||||||||
| 1BS_5144C54E4 | ||||||||||||||
| 4BS_06EE678FF | ||||||||||||||
| miR399 | 2g34208 | * osa MIR399- a-k | * zma MIR399- a-j | * sbi MIR399- a-k | * bdi-MIR399 a-b | * tae-MIR399 | * hvu-MIR399 | *gma-MIR399 a-h | * sly- MIR399 | *stu-MIR399 a-o | * bna-MiR399 a-c | * vvi-MIR399 a-i | *mtr- MIR399 a-t | * ptc-MI399 a-e |
| 1g63005 | ||||||||||||||
| 5g62162 | ||||||||||||||
| 2g34202 | ||||||||||||||
| 2g34204 | ||||||||||||||
| 2g34208 | ||||||||||||||
| MYB62 | 1g68320 | 01g03720 | 096358 162709 | 03g007360 | 2g01960 | 3AS_0A8F6B069 | 65555 | 10G41930 20G25110 | 05g009230 | 14550 | 04041 04297 | 01s0011g03730 | 1g110460 | 008s12130 010s13290 |
| 3AS_5E49D447AF | ||||||||||||||
| 3B_5033C9A8 | ||||||||||||||
| PHF1 | 3g52190 | 07g09000 | 158489 | 02g005080 | 1g55000 | 7BL_48A3E8A28 | 22417 | 10G34380 | 11g007780 | * G404027324 | 06909 | 13s0074g00010 | ND | 008s02920 |
| 5DL_42842D4B4 | 20G33180 | * G402027324 | 010s23870 | |||||||||||
| PHO1 | 3g23430 | 02g56510 | 466545 | 04g036730 | 3g54920 | 6DL_C4D2B23A | 12153 56639 | 10G00720 | 09g090360 | 17163 | 14948 23727 | 05s0049g01410 | 2g077690 1g075640 | 008s16930 010s07970 008s18760 |
| 6BL_6D7561359 | 10G32670 | |||||||||||||
| 20G34390 | ||||||||||||||
| 6BL_0325BEAA4 | 02G00640 | |||||||||||||
| 6BL_CB0A10DC9 | ||||||||||||||
| PHO2 | 2g33770 | 05g48390 05g48400 | 381709 | 09g028110 | 2g16960 | 1DL_F122253A3 1AL_E5224B6EA 5BL_196F297B6 1BL_241A3B9EF | 53410 | 07G31360 | 02g078210 | 29724 | 21874 | 00s1927g00020 00s0265g00070 | 2g013650 4g020620 | 004s04350 011s05240 |
| 13G24810 | ||||||||||||||
| 13G31290 | ||||||||||||||
| 15G08040 | ||||||||||||||
| PHR1 | 4g28610 | 03g21240 07g25710 | 006477 162409 | 01g036440 02g010520 | 1g63530 1g28920 | 3DS_0240F189D 4AS_7220D33B3 3AS_6EEB8D2B2 3B_6780F56DD | 5585 60198 | 19G35080 10G04540 03G32350 | 06g008200 09g072830 | 23467 | 10355 11042 24188 | 07s0005g04120 | 7g098250 | 002s25960 |
| PHT1;1 | 5g43350 | 04g10800 | 170208 | 06g002560 | 5g02750 | 4AL_81AC819EF | 02G0084010G00950 | 03g00530 03g00560 | 13403 | 27491 | 05s0049g00920 05s0049g00930 | 1g043200 1g043290 1g043220 | 0010s08270 | |
| 10g33030 | 35977 | |||||||||||||
| 20G34616 | 33675 | |||||||||||||
| PHT1;4 | 2g38940 | 08g45000 | 045473 154090 | 06g002800 01g046900 01g020570 | 5g02730 3g27680 | 4AL_C56125840 | 03G31950 | 06g034200 | 03798 03839 10288 10289 17503 | 0107 5069 5070 5071 17093 17275 | 07s0005g03290 07s0005g03300 | 5g068140 | 005s24500 005s24510 | |
| 04g10690 | 10G04230 | |||||||||||||
| 03g05610 | 13G18421 | 09g066410 | ||||||||||||
| 10g30790 | 19G34710 | 09g073010 | ||||||||||||
| PHT2;1 | 3g26570 | 02g38020 | 092780 | 04g024630 | 3g47550 | 6AL_294A9853 2 | 6818 | 08G38840 18G20870 | 05g013510 | 18604 | 25212 34242 | 00s0291g00060 | 8g069390 | 008s19070 |
| 6BL_6BC705098 | ||||||||||||||
| 6BL_5C168B1DD | ||||||||||||||
| 7DL_53DB0C6B3 | ||||||||||||||
| PLD1 | 3g16785 | 05g29050 01g20860 | 066485 | 01g017850 03g012720 | 2g27950 | 1AL_11AB1B224 | 70374 | 09G04620 15G16270 | 01g065720 | 09598 | 22178 | 05s0077g01830 | 1g083620 | 008s23240 010s00850 |
| 1BL_884FA4347 | ||||||||||||||
| 1DL_AA198212F | ||||||||||||||
| PTF1 | 5g58010 | 06g09370 | 024530 | 10g006250 | 1g46700 | 7AS_D56F0F9E5 | 59503 | 06G38935 | 12g100140 | 22058 | 20398 06788 | 11s0016g00380 | 3g027650 | 006s13790 |
| 7BS_13A10F94A | ||||||||||||||
| 7DS_68E3ACF8E | ||||||||||||||
| SPX1 | 5g20150 | 06g40120 | 171423 | 04g006990 10g023590 | 1g36610 3g07470 | 7AL_2DSB863A7 | 54859 | 06G07260 | 12g009480 | 02890 | 06543 20088 | 11s0016g05330 | 1g012440 | 006s06880 018s13140 |
| 7BL_DD88849AE | ||||||||||||||
| 7DL_B994066FA | ||||||||||||||
| SPX3 | 2g45130 | 10g25310 03g29250 | 370780 | 01g032880 01g023270 | 1g60250 | 2BS_E9B276FE4 | 62152 | 01G33170 03G03820 | 01g090890 | 2601 | 00373 40324 | 05s0077g00270 | ND | 002s14470 014s06020 017s00240 |
| 7AL_40678A3B5 | ||||||||||||||
| 2AS_96D993EDD | ||||||||||||||
| 7DL_AC271F4EE | ||||||||||||||
| SQD2 | 5g01220 | 07g01030 01g04920 03g15840 | 100652 049190 | 02g000240 03g006480 01g040150 | 1g59860 2g02800 1g67200 | 3B_4DEB64E2E | 63045 12521 | 07G17680 01G27840 03G14200 | 09g014300 10g085100 | 11091 20317 | 05694 09633 | 08s0007g01940 | 4g015260 7g067340 | 006s09880 016s12010 |
| 3DS_CBE46365A | ||||||||||||||
| 2BL_4CF076AAB | ||||||||||||||
| 2AL_218CD1AAD | ||||||||||||||
| 3B_4DEB64E2E | ||||||||||||||
| ZAT6 | 5g04340 | 03g32230 | 106026 | 01g031900 | 1g16010 | 4BS_FE945DFC | 54674 | 17G35430 | 01g107170 | 34322 | 09464 | 03s0091g00690 | 1g106730 | 002s12010 |
| 2g49250 | 4DS_98E9655C8 | |||||||||||||
| 4g03340 | 4DS_676CACEAC | 51405 | ||||||||||||
| 70662 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ajmera, I.; Hodgman, T.C.; Lu, C. An Integrative Systems Perspective on Plant Phosphate Research. Genes 2019, 10, 139. https://doi.org/10.3390/genes10020139
Ajmera I, Hodgman TC, Lu C. An Integrative Systems Perspective on Plant Phosphate Research. Genes. 2019; 10(2):139. https://doi.org/10.3390/genes10020139
Chicago/Turabian StyleAjmera, Ishan, T. Charlie Hodgman, and Chungui Lu. 2019. "An Integrative Systems Perspective on Plant Phosphate Research" Genes 10, no. 2: 139. https://doi.org/10.3390/genes10020139
APA StyleAjmera, I., Hodgman, T. C., & Lu, C. (2019). An Integrative Systems Perspective on Plant Phosphate Research. Genes, 10(2), 139. https://doi.org/10.3390/genes10020139