Metagenome Profiling Identifies Potential Biocontrol Agents for Selaginella kraussiana in New Zealand

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction
2.3. Sequencing
2.4. Data Processing
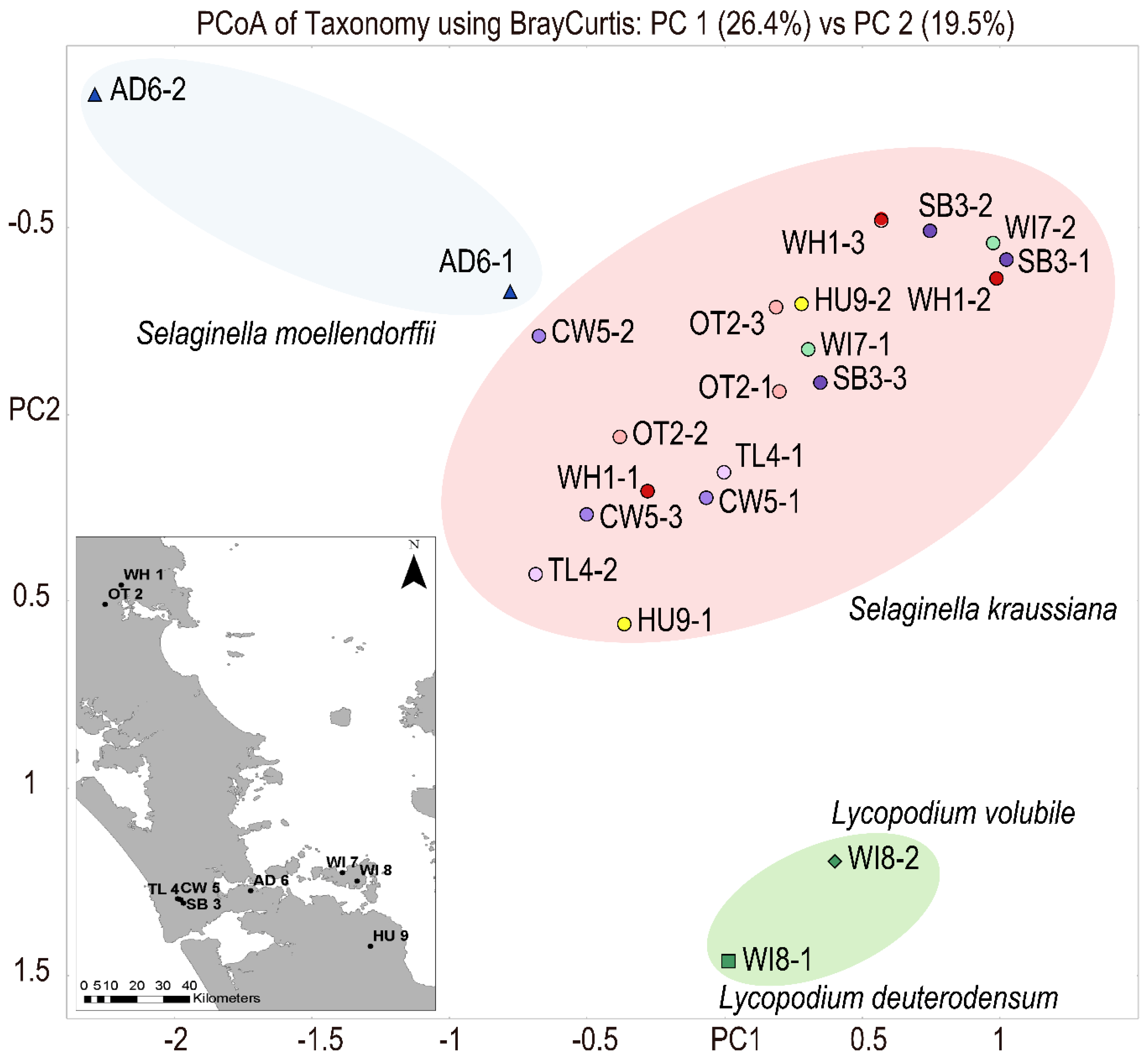
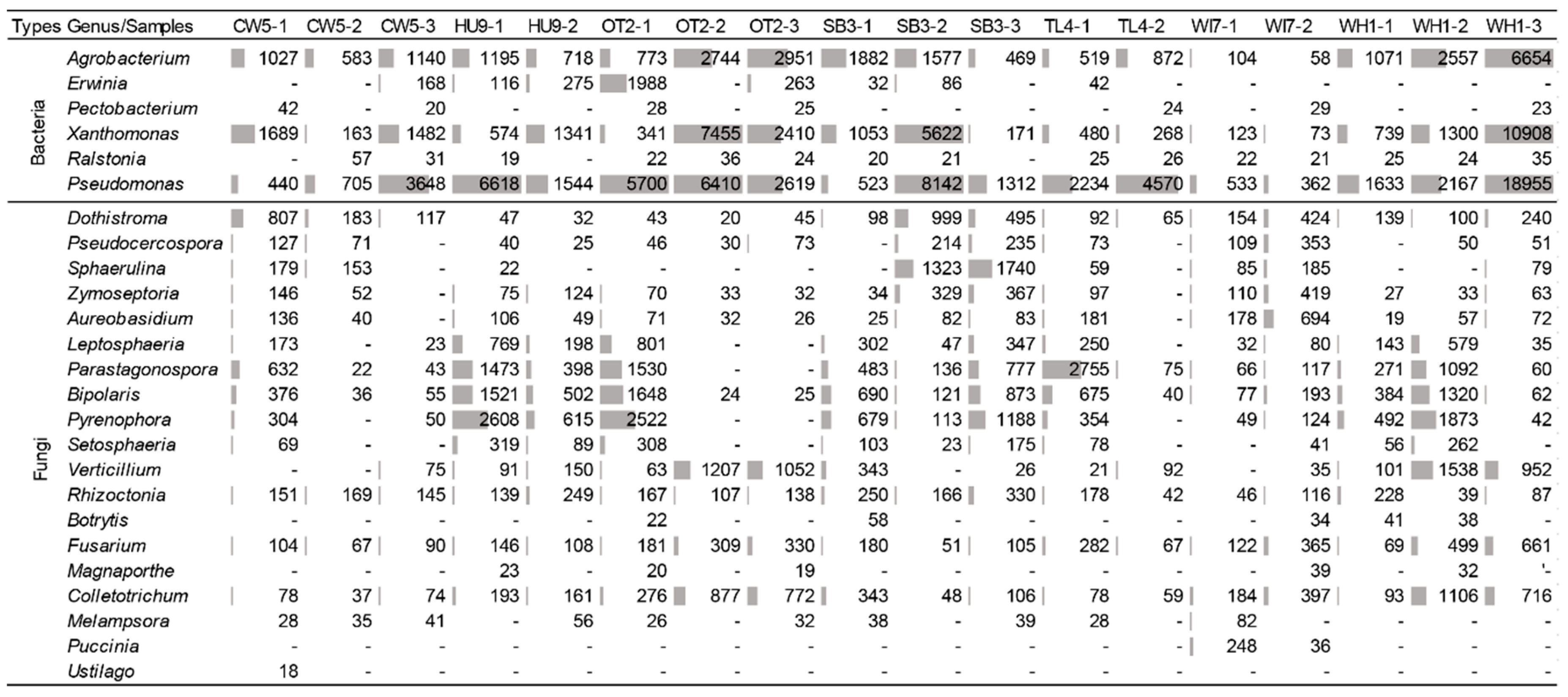
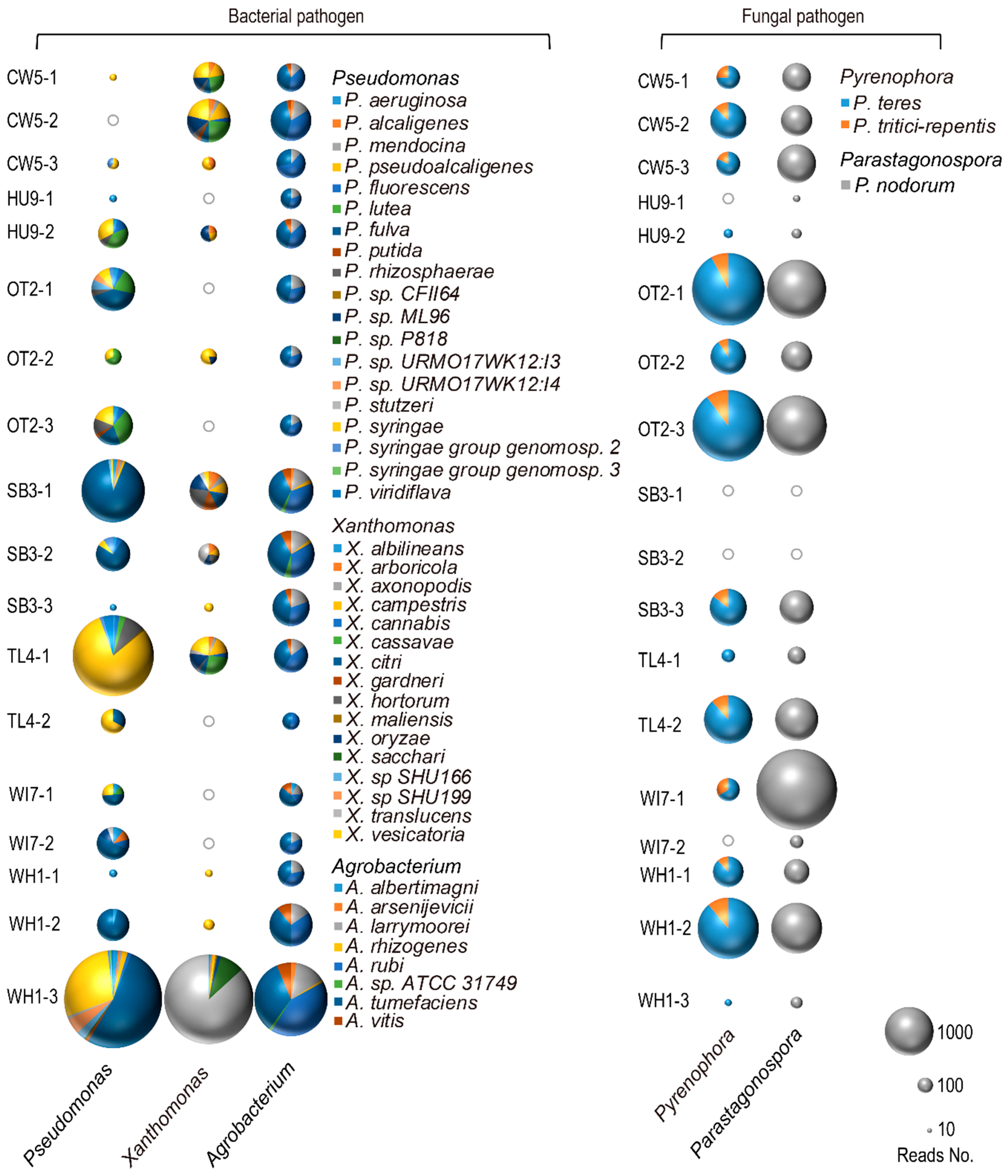
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Keane, M.J.; Crawley, R.M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Burkhart, K.; Nentwig, W. Control of Impatiens glandulifera (Balsaminaceae) by antagonists in its invaded range. Invasive Plant Sci. Manag. 2008, 1, 352–358. [Google Scholar] [CrossRef]
- Flory, S.L.; Clay, K. Pathogen accumulation and long-term dynamics of plant invasions. J. Ecol. 2013, 101, 607–613. [Google Scholar] [CrossRef]
- Najberek, K.; Nentwig, W.; Olejniczak, P.; Król, W.; Baś, C.; Solarz, W. Factors limiting and promoting invasion of alien Impatiens balfourii in Alpine foothills. Flora 2017, 234, 224–232. [Google Scholar] [CrossRef]
- Diez, J.M.; Dickie, I.; Edwards, G.; Hulme, P.E.; Sullivan, J.J.; Duncan, R.P. Negative soil feedbacks accumulate over time for non-native plant species. Ecol. Lett. 2010, 13, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.E.; Blumenthal, D.; Jarošík, V.; Puckett, E.E.; Pyšek, P. Controls on pathogen species richness in plants’ introduced and native ranges: Roles of residence time, range size and host traits. Ecol. Lett. 2010, 13, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.M.; Li, S.; Liao, H.X.; Peng, S.L. Do forest soil microbes have the potential to resist plant invasion? A case study in Dinghushan Biosphere Reserve (South China). Acta Oecol. 2017, 81, 1–9. [Google Scholar] [CrossRef]
- Stricker, K.; Harmon, P.; Goss, E.; Clay, K.; Flory, S.L. Emergence and accumulation of novel pathogens suppress an invasive species. Ecol. Lett. 2016, 19, 469–477. [Google Scholar] [CrossRef]
- Crous, C.J.; Burgess, T.I.; Le Roux, J.J.; Richardson, D.M.; Slippers, B.; Wingfield, M.J. Ecological disequilibrium drives insect pest and pathogen accumulation in non-native trees. AoB Plants 2017, 9, plw081. [Google Scholar] [CrossRef]
- Dostál, P.; Müllerová, J.; Klinerová, P.; Pyšek, T.; Pergl, J. The impact of an invasive plant changes over time. Ecol. Lett. 2013, 16, 1277–1284. [Google Scholar] [CrossRef]
- Mitchell, C.E.; Power, A.G. Release of invasive plants from fungal nature, and viral pathogens. Nature 2003, 421, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Barton, J. How good are we at predicting the field host-range of fungal pathogens used for classical biological control of weeds? Biol. Control 2004, 31, 99–122. [Google Scholar] [CrossRef]
- Ellison, C.A.; Barreto, R.W. Prospects for the management of invasive alien weeds using co-evolved fungal pathogens: A Latin American perspective. Biol. Invasions 2004, 6, 23–45. [Google Scholar] [CrossRef]
- Barton, J.; Fowler, S.V.; Gianotti, A.F.; Winks, C.J.; de Beurs, M.A.; Arnold, G.C.; Forrester, G. Successful biological control of mist flower (Ageratina riparia) in New Zealand: Agent establishment, impact and benefits to the native flora. Biol. Control 2007, 40, 370–385. [Google Scholar] [CrossRef]
- Scott, J.K.; Yeoh, P.B.; Michael, P.J. Methods to select areas to survey for biological control agents: An example based on growth in relation to temperature and distribution of the weed Conyza bonariensis. Biol. Control 2016, 97, 21–30. [Google Scholar] [CrossRef]
- Harding, D.P.; Raizada, M.N. Controlling weeds with fungi, bacteria and viruses: A review. Front. Plant Sci. 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Van Leeuwen, J.F.N.; Schäfer, H.; van der Knaap, W.O.; Rittenour, T.; Björck, S.; Ammann, B. Native or introduced? Fossil pollen and spores may say. An example from the Azores Islands. Neobiota 2005, 6, 27–34. [Google Scholar]
- Nessia, H.; Dale, A.; Perrott, J.; Waipara, N.; Aguilar, G.; Blanchon, D. Comparison of species richness and frequency cover of forest floor plants and lichens in sites invaded and uninvaded by the invasive club moss Selaginella kraussiana (Kunze) A. Braun. Plant Prot. Q. 2014, 29, 66–70. [Google Scholar]
- Cheeseman, T.F. Contributions to a fuller knowledge of the flora of New Zealand, No 6. Trans. N. Z. Inst. 1919, 51, 85–92. [Google Scholar]
- Barton, J. Prospects for Biological Control of African Club Moss (Selaginella kraussiana: Selaginellaceae); Landcare Reserch Contract Report: LC0405/066; Auckland Regional Council: Auckland, New Zealand, 2005; pp. 1–24. [Google Scholar]
- McClymont, M.; Waipara, N.; Nessia, H.; Blanchon, D.J. First record of Phoma selaginellicola on Selaginella kraussiana (African clubmoss): An invasive plant species in New Zealand. Plant Pathol. Quar. 2013, 3, 140–144. [Google Scholar] [CrossRef]
- McClymont, M.; Nessia, H.; Waipara, N.; Blanchon, D.J. First report of Pestalotiopsis clavispora from Selaginella kraussiana (African club moss): An invasive plant species in New Zealand. Australas. Plant Dis. Notes 2013, 8, 79–80. [Google Scholar] [CrossRef]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, e485. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-hadidi, M. MEGAN Community Edition-Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [PubMed]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, e11257. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar]
- Langlois, P.A.; Snelling, J.; Hamilton, J.P.; Bragard, C.; Koebnik, R.; Verdier, V.; Triplett, L.R.; Blom, J.; Tisserat, N.A.; Leach, J.E. Characterization of the Xanthomonas translucens Complex Using Draft Genomes, Comparative Genomics, Phylogenetic Analysis, and Diagnostic LAMP Assays. Phytopathology 2017, 107, 519–527. [Google Scholar] [CrossRef]
- Vancheva, T.; Lefeuvre, P.; Bogatzevska, N.; Moncheva, P.; Koebnik, R. Draft Genome Sequences of Two Xanthomonas euvesicatoria strains from the Balkan Peninsula. Genome Announc. 2015, 3, 3–4. [Google Scholar] [CrossRef]
- Gelvin, S.B. Agrobacterium in the Genomics Age. Plant Physiol. 2009, 150, 1665–1676. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ellwood, S.R.; Oliver, R.P.; Friesen, T.L. Pyrenophora teres: Profile of an increasingly damaging barley pathogen. Mol. Plant Pathol. 2010, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, S.; Sehgal, S.K.; Ali, S.; Liatukas, Z.; Ittu, M.; Kaur, N. Characterization of Pyrenophora tritici-repentis (Tan Spot of Wheat) Races in Baltic States and Romania. Plant Pathol. J. 2017, 33, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.K.; Wyatt, N.A.; Liu, Z.; Faris, J.D.; Friesen, T.L. Reference Quality Genome Assemblies of Three Parastagonospora nodorum Isolates Differing in Virulence on Wheat. G3 Genes Genom. Genet. 2018, 8, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Yarza, P.; Yilmaz, P.; Panzer, K.; Glöckner, F.O.; Reich, M. A phylogenetic framework for the kingdom Fungi based on 18S rRNA gene sequences. Mar. Genom. 2017, 36, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A. Reply to Kiss: Internal transcribed spacer (ITS) remains the best candidate as a universal DNA barcode marker for Fungi despite imperfections. Proc. Natl. Acad. Sci. USA 2012, 109, e1812. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, Z.; McLenachan, P.A.; Lockhart, P.J.; Waipara, N.; Er, O.; Reynolds, C.; Blanchon, D. Metagenome Profiling Identifies Potential Biocontrol Agents for Selaginella kraussiana in New Zealand. Genes 2019, 10, 106. https://doi.org/10.3390/genes10020106
Dang Z, McLenachan PA, Lockhart PJ, Waipara N, Er O, Reynolds C, Blanchon D. Metagenome Profiling Identifies Potential Biocontrol Agents for Selaginella kraussiana in New Zealand. Genes. 2019; 10(2):106. https://doi.org/10.3390/genes10020106
Chicago/Turabian StyleDang, Zhenhua, Patricia A. McLenachan, Peter J. Lockhart, Nick Waipara, Orhan Er, Christy Reynolds, and Dan Blanchon. 2019. "Metagenome Profiling Identifies Potential Biocontrol Agents for Selaginella kraussiana in New Zealand" Genes 10, no. 2: 106. https://doi.org/10.3390/genes10020106
APA StyleDang, Z., McLenachan, P. A., Lockhart, P. J., Waipara, N., Er, O., Reynolds, C., & Blanchon, D. (2019). Metagenome Profiling Identifies Potential Biocontrol Agents for Selaginella kraussiana in New Zealand. Genes, 10(2), 106. https://doi.org/10.3390/genes10020106