The Molecular Characteristics of the FAM13A Gene and the Role of Transcription Factors ACSL1 and ASCL2 in Its Core Promoter Region

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Construction of the Phylogenetic Tree of the FAM13A Gene
2.3. Tissue Expression Analysis of mRNA
2.4. FAM13A Gene Promoter Region Cloning
2.5. Bovine Preadipocyte Culture and Cell Transfection
2.6. Site-Specific Mutation of TFs and Mutation Vector Construction
2.7. EMSA Validation of Transcription Factor Binding
2.8. Statistical Analysis
3. Results
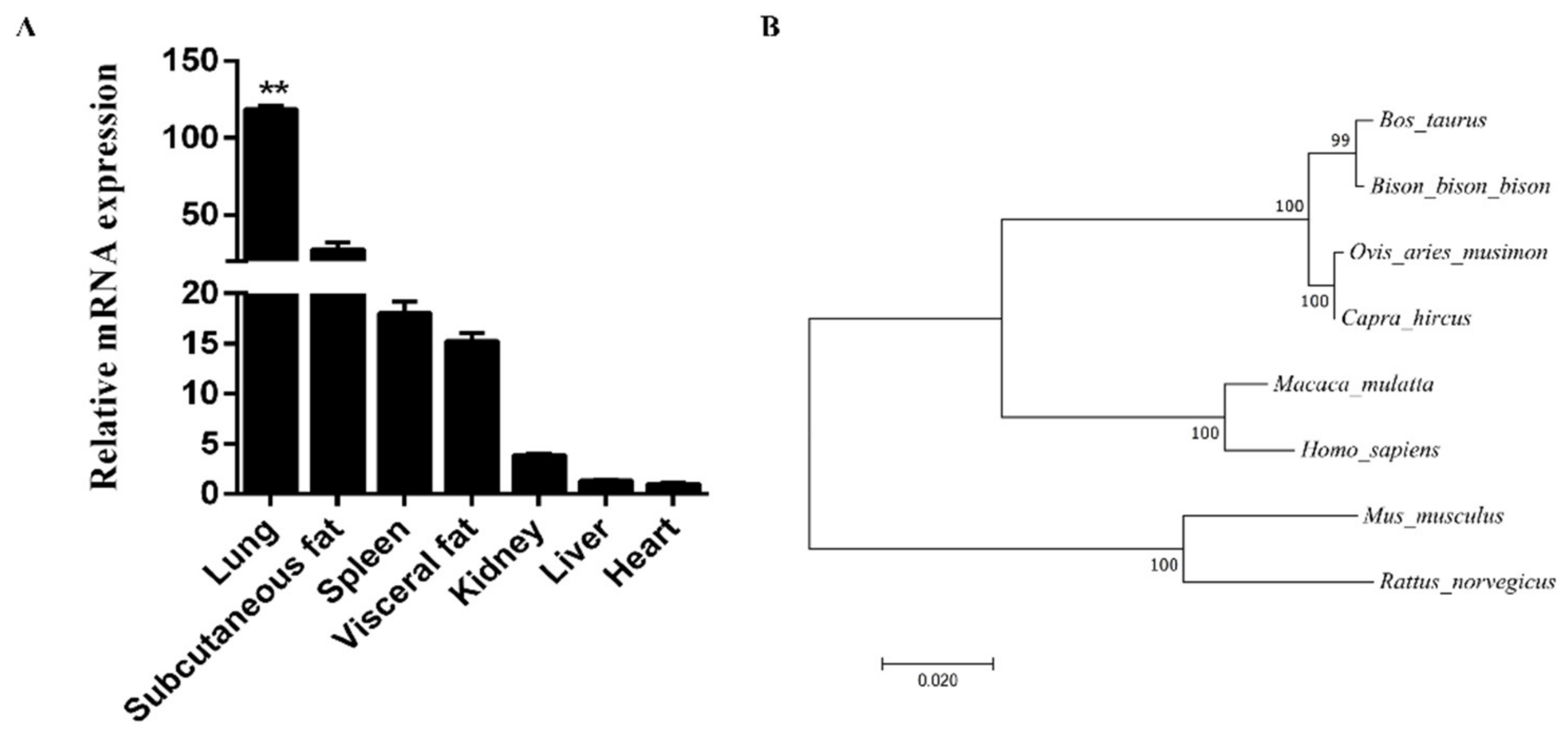
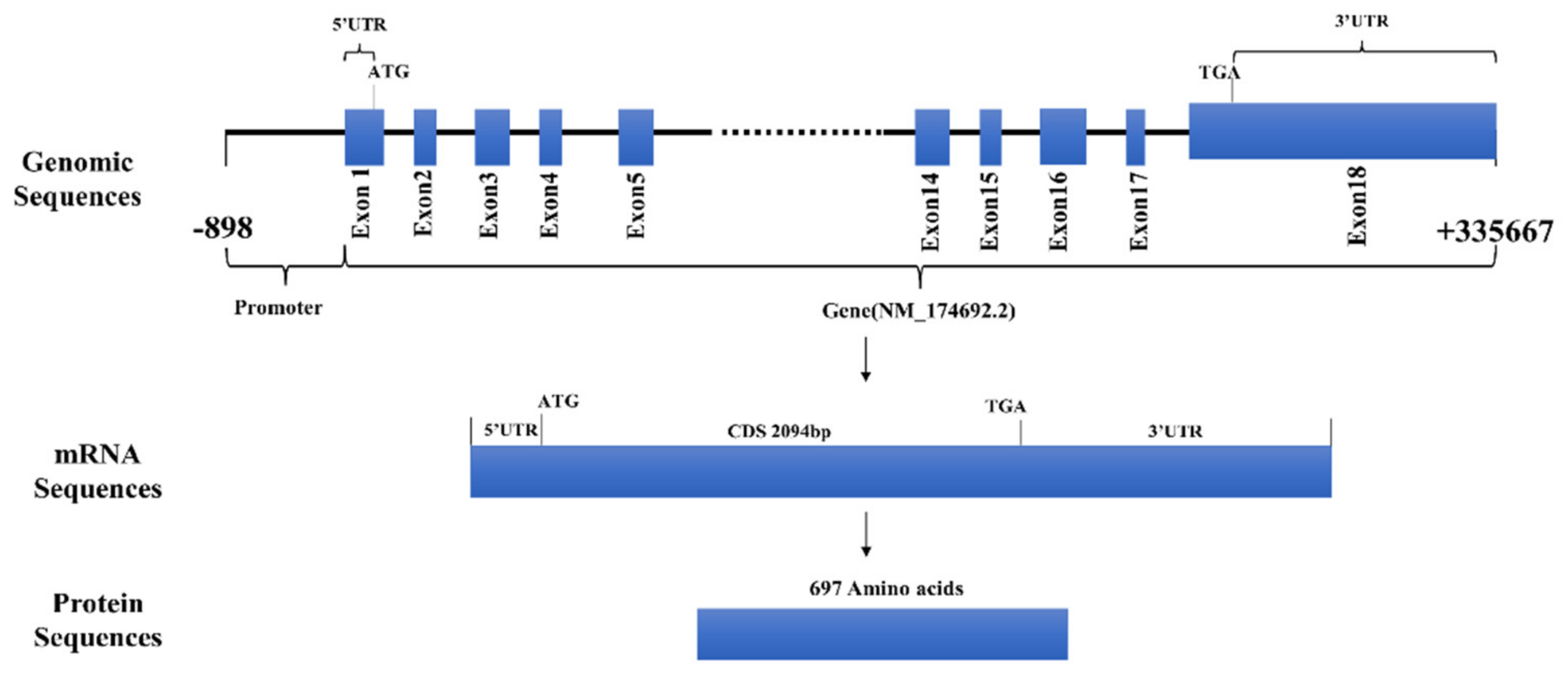
3.1. Expression Patterns and Bioinformatics Analysis of FAM13A Genes
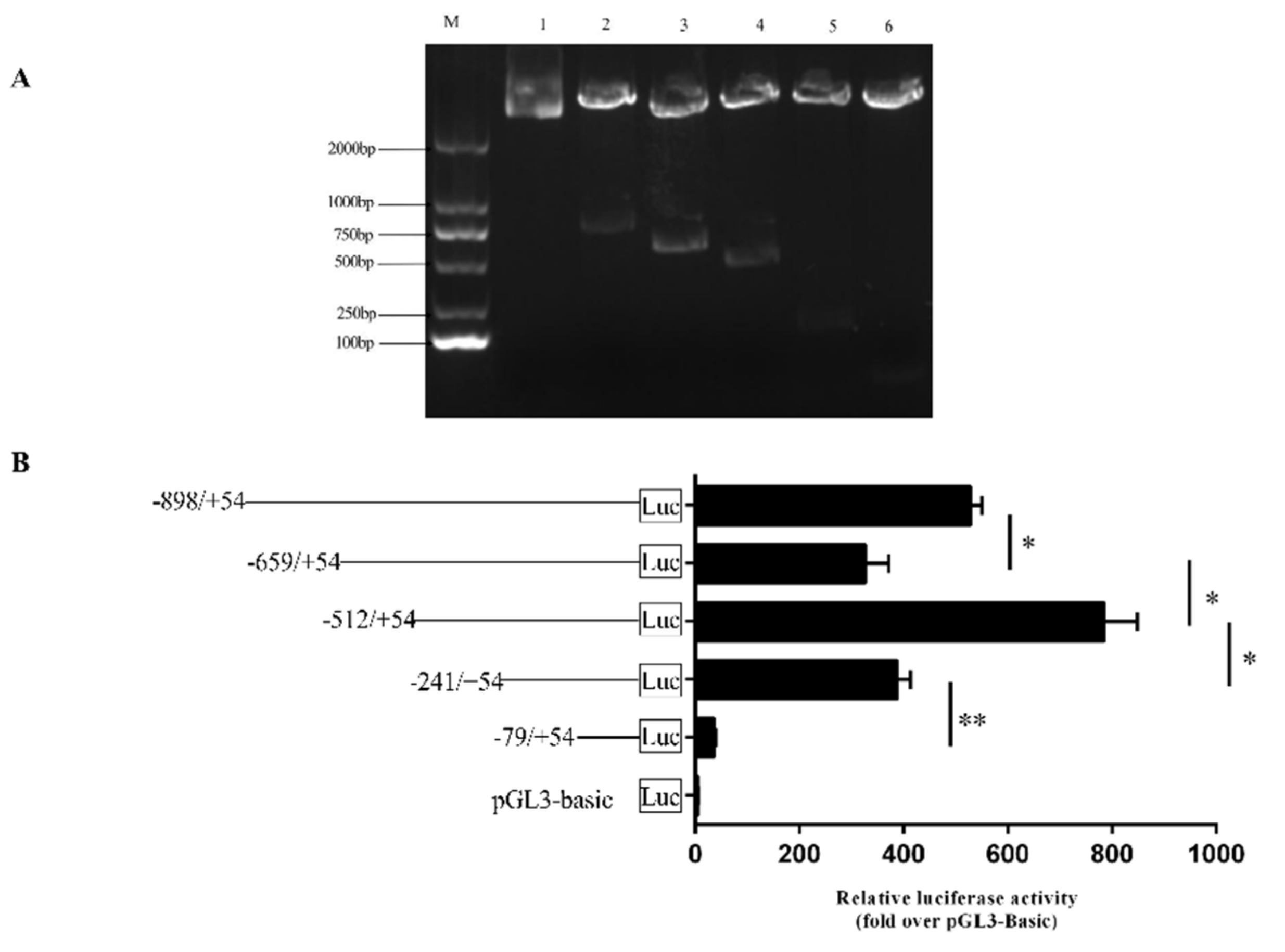
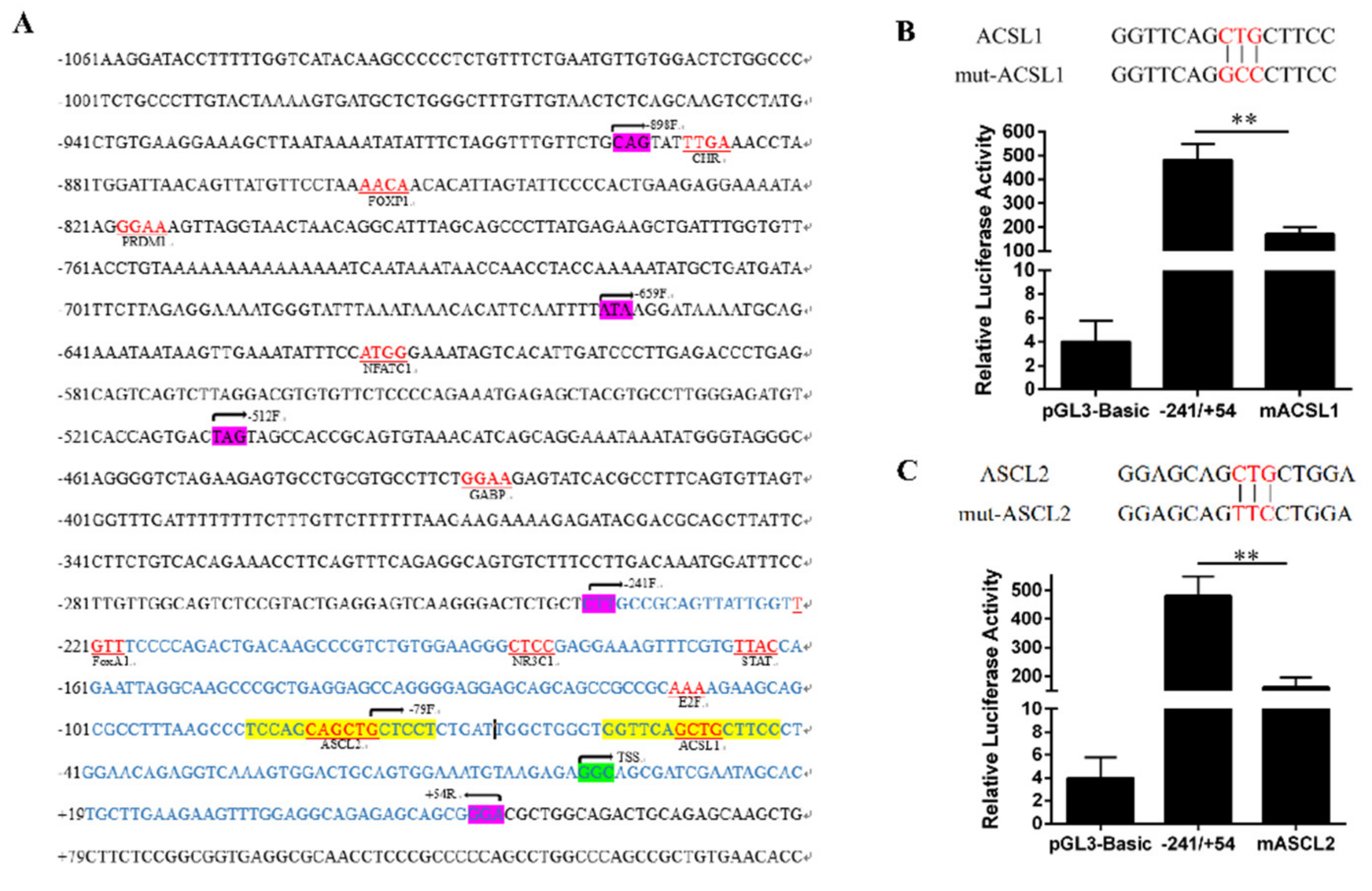
3.2. Transcription Factor Prediction and Enzyme Activity Determination of FAM13A Promoter Region.
3.3. EMSA Experimental Verification of ACSL1 and ASCL2 Transcription Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Albrecht, E.; Gotoh, T.; Ebara, F.; Xu, J.X.; Viergutz, T.; Nurnberg, G.; Maak, S.; Wegner, J. Cellular conditions for intramuscular fat deposition in Japanese Black and Holstein steers. Meat Sci. 2011, 89, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, S.P.; Nathanielsz, P.W. Fetal programming of skeletal muscle development in ruminant animals. J. Anim. Sci. 2010, 88, E51–E60. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Khan, R.; Abdelnour, S.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Taha, A.; Ohran, H.; Mei, C.; Schreurs, N.M.; Zan, L. Advances of Molecular Markers and Their Application for Body Variables and Carcass Traits in Qinchuan Cattle. Genes 2019, 10, 717. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Khan, R.; Schreurs, N.M.; Guo, H.; Gui, L.; Mei, C.; Zan, L. Expression of the bovine KLF6 gene polymorphisms and their association with carcass and body measures in Qinchuan cattle (Bos Taurus). Genomics 2019, in press. [Google Scholar] [CrossRef]
- Raza, S.H.A.; Gui, L.; Khan, R.; Schreurs, N.M.; Xiaoyu, W.; Wu, S.; Mei, C.; Wang, L.; Ma, X.; Wei, D.; et al. Association between FASN gene polymorphisms ultrasound carcass traits and intramuscular fat in Qinchuan cattle. Gene 2018, 645, 55–59. [Google Scholar] [CrossRef]
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef]
- Wei, D.; Li, A.; Zhao, C.; Wang, H.; Mei, C.; Khan, R.; Zan, L. Transcriptional regulation by CpG sites methylation in the core promoter region of the bovine SIX1 gene: Roles of histone H4 and E2F2. Int. J. Mol. Sci. 2018, 19, 213. [Google Scholar] [CrossRef]
- Guo, H.; Raza, S.H.A.; Schreurs, N.M.; Khan, R.; Wei, D.; Wang, L.; Zhang, S.; Zhang, L.; Wu, S.; Ullah, I.; et al. Genetic variants in the promoter region of the KLF3 gene associated with fat deposition in Qinchuan cattle. Gene 2018, 672, 50–55. [Google Scholar] [CrossRef]
- Wei, D.; Raza, S.H.A.; Zhang, J.; Gui, L.; Rahman, S.U.; Khan, R.; Hosseini, S.M.; Kaleri, H.A.; Zan, L. Polymorphism in promoter of SIX4 gene shows association with its transcription and body measurement traits in Qinchuan cattle. Gene 2018, 656, 9–16. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell Proteomics 2014, 13, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liang, B.; Yang, J.; Xiao, J.; Ma, C.; Xu, S.; Lei, J.; Xu, X.; Liao, Z.; Liu, H.; et al. Association of FAM13A polymorphisms with COPD and COPD-related phenotypes in Han Chinese. Clin. Biochem. 2013, 46, 1683–1688. [Google Scholar] [CrossRef] [PubMed]
- Lundbäck, V.; Kulyte, A.; Strawbridge, R.J.; Ryden, M.; Arner, P.; Marcus, C.; Dahlman, I. FAM13A and POM121C are candidate genes for fasting insulin: Functional follow-up analysis of a genome-wide association study. Diabetologia 2018, 61, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Young, R.P.; Hopkins, R.J.; Hay, B.A.; Whittington, C.F.; Epton, M.J.; Gamble, G.D. FAM13A locus in COPD is independently associated with lung cancer–evidence of a molecular genetic link between COPD and lung cancer. Appl Clin Genet. 2011, 4, 1. [Google Scholar] [CrossRef]
- Zhang, Y.; Qiu, J.; Zhang, P.; Zhang, J.; Jiang, M.; Ma, Z. Genetic variants in FAM13A and IREB2 are associated with the susceptibility to COPD in a Chinese rural population: A case-control study. Int. J. Chronic Obstr. Pulm. Dis. 2018, 13, 1735. [Google Scholar] [CrossRef]
- Wardhana, D.A.; Ikeda, K.; Barinda, A.J.; Nugroho, D.B.; Qurania, K.R.; Yagi, K.; Miyata, K.; Oike, Y.; Hirata, K.; Emoto, N. Family with sequence similarity 13, member A modulates adipocyte insulin signaling and preserves systemic metabolic homeostasis. Proc. Natl. Acad. Sci. USA 2018, 115, 1529–1534. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Li, A.; Zhang, Y.; Zhao, Z.; Wang, M.; Zan, L. Molecular Characterization and Transcriptional Regulation Analysis of the Bovine PDHB Gene. PLoS ONE 2016, 11, e0157445. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Khan, R.; Raza, S.H.A.; Li, A.; Zhang, Y.; Liang, C.; Yang, W.; Wu, S.; Zan, L. Molecular characterization of ABHD5 gene promoter in intramuscular preadipocytes of Qinchuan cattle: Roles of Evi1 and C/EBPα. Gene 2019, 690, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Chen, Y.; Zhao, X.; Niu, Y.; Cong, P.; Zhang, Z.; Chen, W.; Jiang, W.; Mo, D. Characterization and transcriptional regulation analysis of the porcine TNFAIP8L2 gene. Mol. Genet. Genomic 2010, 284, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Raza, S.H.A.; Schreurs, N.; Yu, W.; Hongbao, W.; Ullah, I.; Rahman, A.; Suhail, S.M.; Khan, S.; Linsen, Z. Bioinformatics analysis and transcription regulation of TORC1 gene through transcription factors NRF1 and Smad3 in bovine preadipocytes. Genomics 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Raza, S.H.A.; Junjvlieke, Z.; Xiaoyu, W.; Garcia, M.; Elnour, I.E.; Hongbao, W.; Linsen, Z. Function and Transcriptional Regulation of Bovine TORC2 Gene in Adipocytes: Roles of C/EBP, XBP1, INSM1 and ZNF263. Int. J. Mol. Sci. 2019, 20, 4338. [Google Scholar] [CrossRef]
- Wei, D.-W.; Gui, L.-S.; Raza, S.H.A.; Zhang, S.; Khan, R.; Wang, L.; Guo, H.-F.; Zan, L.-S. NRF1 and ZSCAN10 bind to the promoter region of the SIX1 gene and their effects body measurements in Qinchuan cattle. Sci. Rep. 2017, 7, 7867. [Google Scholar] [CrossRef]
- Guo, H.; Khan, R.; Raza, S.H.A.; Ning, Y.; Wei, D.; Wu, S.; Hosseini, S.M.; Ullah, I.; Garcia, M.D.; Zan, L. KLF15 promotes transcription of KLF3 gene in bovine adipocytes. Gene 2018, 659, 77–83. [Google Scholar] [CrossRef]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of lipolysis in adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef]
- Brown, A.E.; Walker, M. Genetics of insulin resistance and the metabolic syndrome. Curr. Cardiol. Rep. 2016, 18, 75. [Google Scholar] [CrossRef]
- Copps, K.D.; White, M.F. Regulation of insulin sensitivity by serine/threonine phosphorylation of insulin receptor substrate proteins IRS1 and IRS2. Diabetologia 2012, 55, 2565–2582. [Google Scholar] [CrossRef]
- Morley, T.S.; Xia, J.Y.; Scherer, P.E. Selective enhancement of insulin sensitivity in the mature adipocyte is sufficient for systemic metabolic improvements. Nat. Commun. 2015, 6, 7906. [Google Scholar] [CrossRef] [PubMed]
- Castaldi, P.J.; Guo, F.; Qiao, D.; Du, F.; Naing, Z.Z.C.; Li, Y.; Pham, B.; Mikkelsen, T.S.; Cho, M.H.; Silverman, E.K.; et al. Identification of functional variants in the FAM13A chronic obstructive pulmonary disease genome-wide association study locus by massively parallel reporter assays. Am. J. Respir. Crit. Care Med. 2019, 199, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Junjvlieke, Z.; Mei, C.-G.; Khan, R.; Zhang, W.-Z.; Hong, J.-Y.; Wang, L.; Li, S.-J.; Zan, L.-S. Transcriptional regulation of bovine elongation of very long chain fatty acids protein 6 in lipid metabolism and adipocyte proliferation. J. Cell Biochem. 2019, 120, 13932–13943. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.; Raza, S.H.A.; Sun, Y.; Khan, R.; Ullah, I.; Han, Y. Detection of polymorphisms in the promoter of bovine SIRT1 gene and their effects on intramuscular fat content in Chinese indigenous cattle. Gene 2019, 700, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Ning, Y.; Raza, S.H.A.; Zhang, C.; Zhang, L.; Cheng, G.; Wang, H.; Schreurs, N.; Zan, L. Genetic variants and haplotype combination in the bovine CRTC3 affected conformation traits in two Chinese native cattle breeds (Bos Taurus). Genomics 2019, 111, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Ohkuni, A.; Ohno, Y.; Kihara, A. Identification of acyl-CoA synthetases involved in the mammalian sphingosine 1-phosphate metabolic pathway. Biochem. Biophys. Res Commun. 2013, 442, 195–201. [Google Scholar] [CrossRef]
- Nakahara, K.; Ohkuni, A.; Kitamura, T.; Abe, K.; Naganuma, T.; Ohno, Y.; Zoeller, R.A.; Kihara, A. The Sjögren-Larsson syndrome gene encodes a hexadecenal dehydrogenase of the sphingosine 1-phosphate degradation pathway. Mol. Cell 2012, 46, 461–471. [Google Scholar] [CrossRef]
- Gui, L.; Wu, H.; Raza, S.H.A.; Schreurs, N.M.; Shah, M.A. The effect of haplotypes in the promoter region of SIRT4 gene on the ultrasound traits in Qinchuan cattle. Trop. Anim. Health Prod. 2019, 51, 1877–1882. [Google Scholar] [CrossRef]
- Gui, L.; Raza, S.H.A.; Garcia, M.; Sun, Y.; Ullah, I.; Han, Y. Genetic variants in the SIRT6 transcriptional regulatory region affect gene activity and carcass quality traits in indigenous Chinese beef cattle (Bos taurus). BMC Genomics 2018, 19, 785. [Google Scholar] [CrossRef]
- Gui, L.; Raza, S.H.A.; Jia, J. Analysis of the oxidized low density lipoprotein receptor 1 gene as a potential marker for carcass quality traits in Qinchuan cattle. Asian Australas. J. Anim. Sci. 2019, 32, 58. [Google Scholar] [CrossRef]
- Bougarne, N.; Viacheslav, M.; Ratman, D.; Beck, I.M.; Thommis, J.; De Cauwer, L.; Staels, B.; Tavernier, J.; Libert, C.; De Bosscher, K. Mechanisms underlying the functional cooperation between PPARalpha and GRalpha to attenuate inflammatory responses. Front. Immunol. 2019, 10, 1769. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′–3′) | Annealing Temperature (°C) |
|---|---|---|
| 18S-F | CCTGCGGCTTAATTTGACTC | 57 |
| 18S-R | AACTAAGAACGGCCATGCAC | 58 |
| FAM13A-F | GTACCGCCTGGTCAAACAGATCCTA | 64 |
| FAM13A-R | TAGTTATCGTCTTCTGAACCCTC | 57 |
| Primer | Primer Sequence (5′–3′) | Annealing Temperature (°C) |
|---|---|---|
| FAM13A(−898)-F | CGGGGTACCCAGTATTTGAAACCTATGGATTAAC | 55 |
| FAM13A(−659)-F | CGGGGTACCATAAGGATAAAATGCAGAAATAATA | 51 |
| FAM13A(−512)-F | CGGGGTACCTAGTAGCCACCGCAGTGTAAACATC | 62 |
| FAM13A(−241)-F | CGGGGTACCCTTGCCGCAGTTATTGGTTGTTTCC | 63 |
| FAM13A(−79)-F | CGGGGTACCGCTCCTCTGATTGGCTGGGTGGTTC | 67 |
| FAM13A(+54)-R | TCCCCCGGGTCCCGCTGCTCTCTGCCTCCAAACT | 69 |
| mACSL1-F | GCTGGGTGGTTCAGACCCTTCCCTGGAA | 71 |
| mACSL1-R | GGTCTGAACCACCCAGCCAATCAGAGGA | 70 |
| mASCL2-F | TAAGCCCTCCAGCAGGCC CTCCTCTGAT | 71 |
| mASCL2-R | GGCCTGCTGGAGGGCTTA AAGGCGCTGC | 74 |
| Primer | Primer Sequence (5′–3′) | Annealing Temperature (°C) |
|---|---|---|
| ACSL1-bio-F | Biotin-TGGCTGGGTGGTTCAGCTGCTTCCCTGGAACAGA | 75 |
| ACSL1-bio-R | Biotin-TCTGTTCCAGGGAAGCAGCTGAACCACCCAGCCA | 75 |
| ACSL1-jz-F | TGGCTGGGTGGTTCAGCTGCTTCCCTGGAACAGA | 75 |
| ACSL1-jz-R | TCTGTTCCAGGGAAGCAGCTGAACCACCCAGCCA | 75 |
| ACSL1-mut-F | TGGCTGGGTGGTTCAGACCCTTCCCTGGAACAGA | 75 |
| ASCL2-mut-R | TCTGTTCCAGGGAAGGGTCTGAACCACCCAGCCA | 75 |
| ASCL2-bio-F | Biotin-TAAGCCCTCCAGCAGCTGCTCCTCTGATTGGCT | 74 |
| ASCL2-bio-R | Biotin-AGCCAATCAGAGGAGCAGCTGCTGGAGGGCTTA | 74 |
| ASCL2-jz-F | TAAGCCCTCCAGCAGCTGCTCCTCTGATTGGCT | 74 |
| ASCL2-jz-R | AGCCAATCAGAGGAGCAGCTGCTGGAGGGCTTA | 74 |
| ASCL2-mut-F | TAAGCCCTCCAGCAGGCCCTCCTCTGATTGGCT | 75 |
| ASCL2-mut-R | AGCCAATCAGAGGAGGGCCTGCTGGAGGGCTTA | 75 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, C.; Li, A.; Raza, S.H.A.; Khan, R.; Wang, X.; Wang, S.; Wang, G.; Zhang, Y.; Zan, L. The Molecular Characteristics of the FAM13A Gene and the Role of Transcription Factors ACSL1 and ASCL2 in Its Core Promoter Region. Genes 2019, 10, 981. https://doi.org/10.3390/genes10120981
Liang C, Li A, Raza SHA, Khan R, Wang X, Wang S, Wang G, Zhang Y, Zan L. The Molecular Characteristics of the FAM13A Gene and the Role of Transcription Factors ACSL1 and ASCL2 in Its Core Promoter Region. Genes. 2019; 10(12):981. https://doi.org/10.3390/genes10120981
Chicago/Turabian StyleLiang, Chengcheng, Anning Li, Sayed Haidar Abbas Raza, Rajwali Khan, Xiaoyu Wang, Sihu Wang, Guohua Wang, Yu Zhang, and Linsen Zan. 2019. "The Molecular Characteristics of the FAM13A Gene and the Role of Transcription Factors ACSL1 and ASCL2 in Its Core Promoter Region" Genes 10, no. 12: 981. https://doi.org/10.3390/genes10120981
APA StyleLiang, C., Li, A., Raza, S. H. A., Khan, R., Wang, X., Wang, S., Wang, G., Zhang, Y., & Zan, L. (2019). The Molecular Characteristics of the FAM13A Gene and the Role of Transcription Factors ACSL1 and ASCL2 in Its Core Promoter Region. Genes, 10(12), 981. https://doi.org/10.3390/genes10120981