High Prevalence of Staphylococcus aureus Enterotoxin Gene Cluster Superantigens in Cystic Fibrosis Clinical Isolates

Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. S. aureus Specimens From Patients With CF Are Heterogeneous in Phenotypic Appearance
3.1.1. Colony Morphology
3.1.2. Antibiotic Resistance
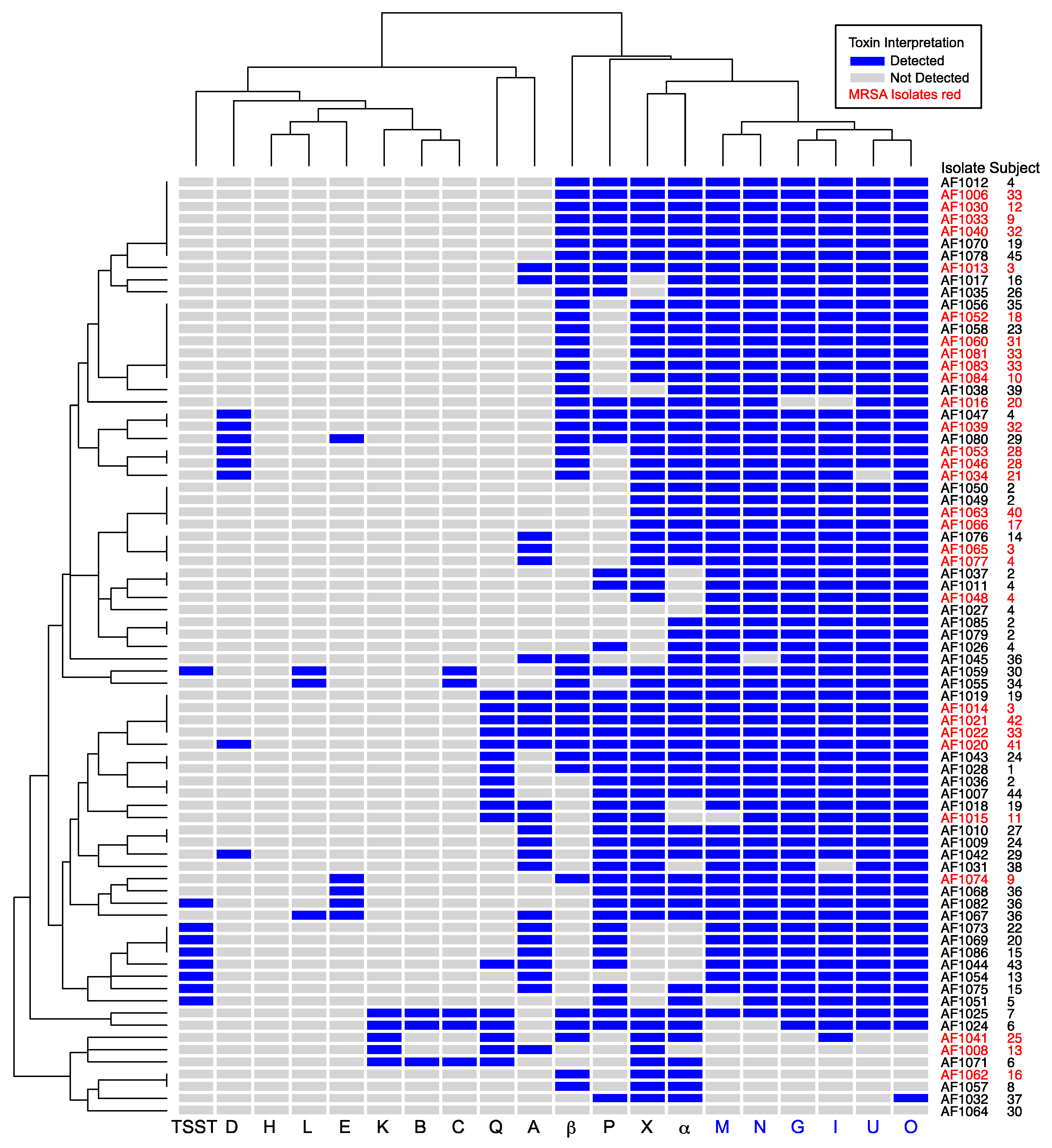
3.2. High Prevalence of Enterotoxin Gene Cluster Genes in S. aureus Isolated From Patients With Cystic Fibrosis
3.2.1. EGC Prevalence in MRSA and MSSA
3.2.2. EGC Prevalence in S. aureus From Other U.S. CF Centers
4. Discussion
4.1. Advantages
4.2. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Salsgiver, E.L.; Fink, A.K.; Knapp, E.A.; LiPuma, J.J.; Olivier, K.N.; Marshall, B.C.; Saiman, L. Changing epidemiology of the respiratory bacteriology of patients with cystic fibrosis. Chest 2016, 149, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Cystic Fibrosis Foundation 2016 Patient Registry Annual Data Report. 2017. Available online: https://www.cff.org/research/researcher-resources/patient-registry/2016-patient-registry-annual-data-report.pdf (accessed on 4 November 2019).
- Ramsey, B.W.; Pepe, M.S.; Quan, J.M.; Otto, K.L.; Montgomery, A.B.; Williams-Warren, J.; Vasiljev, K.M.; Borowitz, D.; Bowman, C.M.; Marshall, B.C.; et al. Intermittent administration of inhaled tobramycin in patients with cystic fibrosis. Cystic fibrosis inhaled tobramycin study group. N. Engl. J. Med. 1999, 340, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.L.; Emerson, J.; McNamara, S.; Burns, J.L.; Rosenfeld, M.; Yunker, A.; Hamblett, N.; Accurso, F.; Dovey, M.; Hiatt, P.; et al. Significant microbiological effect of inhaled tobramycin in young children with cystic fibrosis. Am. J. Respir. Crit. Care Med. 2003, 167, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Tiddens, H.A.; De Boeck, K.; Clancy, J.P.; Fayon, M.; Arets, H.G.M.; Bresnik, M.; Derchak, A.; Lewis, S.A.; Oermann, C.M. Open label study of inhaled aztreonam for Pseudomonas eradication in children with cystic fibrosis: The alpine study. J. Cyst. Fibros. 2015, 14, 111–119. [Google Scholar] [CrossRef]
- Dezube, R.; Jennings, M.T.; Rykiel, M.; Diener-West, M.; Boyle, M.P.; Chmiel, J.F.; Dasenbrook, E.C. Eradication of persistent methicillin-resistant Staphylococcus aureus infection in cystic fibrosis. J. Cyst. Fibros. 2018. [Google Scholar] [CrossRef]
- Heltshe, S.L.; Mayer-Hamblett, N.; Burns, J.L.; Khan, U.; Baines, A.; Ramsey, B.W.; Rowe, S.M. Pseudomonas aeruginosa in cystic fibrosis patients with G551D-CFTR treated with ivacaftor. Clin. Infect. Dis. 2015, 60, 703–712. [Google Scholar] [CrossRef]
- Singh, S.B.; McLearn-Montz, A.J.; Milavetz, F.; Gates, L.K.; Fox, C.; Murry, L.T.; Sabus, A.; Porterfield, H.S.; Fischer, A.J. Pathogen acquisition in patients with cystic fibrosis receiving ivacaftor or lumacaftor/ivacaftor. Pediatr. Pulmonol. 2019. [Google Scholar] [CrossRef]
- Caudri, D.; Turkovic, L.; Ng, J.; de Klerk, N.H.; Rosenow, T.; Hall, G.L.; Ranganathan, S.C.; Sly, P.D.; Stick, S.M. The association between Staphylococcus aureus and subsequent bronchiectasis in children with cystic fibrosis. J. Cyst. Fibros. 2018, 17, 462–469. [Google Scholar] [CrossRef]
- Dasenbrook, E.C.; Merlo, C.A.; Diener-West, M.; Lechtzin, N.; Boyle, M.P. Persistent methicillin-resistant Staphylococcus aureus and rate of FEV1 decline in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2008, 178, 814–821. [Google Scholar] [CrossRef]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef]
- Spaulding, A.R.; Salgado-Pabon, W.; Kohler, P.L.; Horswill, A.R.; Leung, D.Y.; Schlievert, P.M. Staphylococcal and streptococcal superantigen exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef] [PubMed]
- McCormick, J.K.; Yarwood, J.M.; Schlievert, P.M. Toxic shock syndrome and bacterial superantigens: An update. Annu. Rev. Microbiol. 2001, 55, 77–104. [Google Scholar] [CrossRef] [PubMed]
- Jarraud, S.; Peyrat, M.A.; Lim, A.; Tristan, A.; Bes, M.; Mougel, C.; Etienne, J.; Vandenesch, F.; Bonneville, M.; Lina, G. egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J. Immunol. 2001, 166, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Friedrich, A.W.; Peters, G.; von Eiff, C. Systematic survey on the prevalence of genes coding for staphylococcal enterotoxins SELM, SELO, and SELN. Mol. Nutr. Food Res. 2004, 48, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Grumann, D.; Scharf, S.S.; Holtfreter, S.; Kohler, C.; Steil, L.; Engelmann, S.; Hecker, M.; Volker, U.; Broker, B.M. Immune cell activation by enterotoxin gene cluster (egc)-encoded and non-egc superantigens from Staphylococcus aureus. J. Immunol. 2008, 181, 5054–5061. [Google Scholar] [CrossRef]
- Holtfreter, S.; Bauer, K.; Thomas, D.; Feig, C.; Lorenz, V.; Roschack, K.; Friebe, E.; Selleng, K.; Lovenich, S.; Greve, T.; et al. egc-Encoded superantigens from Staphylococcus aureus are neutralized by human sera much less efficiently than are classical staphylococcal enterotoxins or toxic shock syndrome toxin. Infect. Immun. 2004, 72, 4061–4071. [Google Scholar] [CrossRef]
- Stach, C.S.; Vu, B.G.; Merriman, J.A.; Herrera, A.; Cahill, M.P.; Schlievert, P.M.; Salgado-Pabon, W. Novel tissue level effects of the Staphylococcus aureus enterotoxin gene cluster are essential for infective endocarditis. PLoS ONE 2016, 11, e0154762. [Google Scholar] [CrossRef]
- Garbacz, K.; Piechowicz, L.; Podkowik, M.; Mroczkowska, A.; Empel, J.; Bania, J. Emergence and spread of worldwide Staphylococcus aureus clones among cystic fibrosis patients. Infect. Drug Res. 2018, 11, 247–255. [Google Scholar] [CrossRef]
- Mempel, M.; Lina, G.; Hojka, M.; Schnopp, C.; Seidl, H.P.; Schafer, T.; Ring, J.; Vandenesch, F.; Abeck, D. High prevalence of superantigens associated with the egc locus in Staphylococcus aureus isolates from patients with atopic eczema. Eur. J. Clin. Microbiol. Infect. Dis. 2003, 22, 306–309. [Google Scholar] [CrossRef]
- Merriman, J.A.; Mueller, E.A.; Cahill, M.P.; Beck, L.A.; Paller, A.S.; Hanifin, J.M.; Ong, P.Y.; Schneider, L.; Babineau, D.C.; David, G.; et al. Temporal and racial differences associated with atopic dermatitis Staphylococcus aureus and encoded virulence factors. mSphere 2016, 1. [Google Scholar] [CrossRef]
- Dasenbrook, E.C.; Checkley, W.; Merlo, C.A.; Konstan, M.W.; Lechtzin, N.; Boyle, M.P. Association between respiratory tract methicillin-resistant Staphylococcus aureus and survival in cystic fibrosis. JAMA J. Am. Med. Assoc. 2010, 303, 2386–2392. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Pabon, W.; Case-Cook, L.C.; Schlievert, P.M. Molecular analysis of staphylococcal superantigens. Methods Mol. Biol. 2014, 1085, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Buckley, D.G.; Wu, Z.; Saenphimmachak, C.; Hoffman, L.R.; D’Argenio, D.A.; Miller, S.I.; Ramsey, B.W.; Speert, D.P.; Moskowitz, S.M.; et al. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2006, 103, 8487–8492. [Google Scholar] [CrossRef] [PubMed]
- Clauditz, A.; Resch, A.; Wieland, K.P.; Peschel, A.; Gotz, F. Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its ability to cope with oxidative stress. Infect. Immun. 2006, 74, 4950–4953. [Google Scholar] [CrossRef]
- Kolar, S.L.; Ibarra, J.A.; Rivera, F.E.; Mootz, J.M.; Davenport, J.E.; Stevens, S.M.; Horswill, A.R.; Shaw, L.N. Extracellular proteases are key mediators of Staphylococcus aureus virulence via the global modulation of virulence-determinant stability. MicrobiologyOpen 2013, 2, 18–34. [Google Scholar] [CrossRef]
- Jarry, T.M.; Memmi, G.; Cheung, A.L. The expression of alpha-haemolysin is required for Staphylococcus aureus phagosomal escape after internalization in CFT-1 cells. Cell. Microbiol. 2008, 10, 1801–1814. [Google Scholar] [CrossRef]
- Keitsch, S.; Riethmuller, J.; Soddemann, M.; Sehl, C.; Wilker, B.; Edwards, M.J.; Caldwell, C.C.; Fraunholz, M.; Gulbins, E.; Becker, K.A. Pulmonary infection of cystic fibrosis mice with Staphylococcus aureus requires expression of alpha-toxin. Biol. Chem. 2018, 399, 1203–1213. [Google Scholar] [CrossRef]
- Schlievert, P.M.; Strandberg, K.L.; Lin, Y.C.; Peterson, M.L.; Leung, D.Y. Secreted virulence factor comparison between methicillin-resistant and methicillin-sensitive Staphylococcus aureus, and its relevance to atopic dermatitis. J. Allergy Clin. Immunol. 2010, 125, 39–49. [Google Scholar] [CrossRef]
- Vu, B.G.; Stach, C.S.; Salgado-Pabon, W.; Diekema, D.J.; Gardner, S.E.; Schlievert, P.M. Superantigens of Staphylococcus aureus from patients with diabetic foot ulcers. J. Infect. Dis. 2014, 210, 1920–1927. [Google Scholar] [CrossRef]
- Roetzer, A.; Gruener, C.S.; Haller, G.; Beyerly, J.; Model, N.; Eibl, M.M. Enterotoxin gene cluster-encoded sei and seln from Staphylococcus aureus isolates are crucial for the induction of human blood cell proliferation and pathogenicity in rabbits. Toxins 2016, 8, 314. [Google Scholar] [CrossRef]
- Hollsing, A.E.; Granstrom, M.; Strandvik, B. Prospective study of serum staphylococcal antibodies in cystic fibrosis. Arch. Dis. Child. 1987, 62, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Moss, R.B.; Hsu, Y.P.; Lewiston, N.J. 125I-Clq-binding and specific antibodies as indicators of pulmonary disease activity in cystic fibrosis. J. Pediatr. 1981, 99, 215–222. [Google Scholar] [CrossRef]
- Spaulding, A.R.; Salgado-Pabon, W.; Merriman, J.A.; Stach, C.S.; Ji, Y.; Gillman, A.N.; Peterson, M.L.; Schlievert, P.M. Vaccination against Staphylococcus aureus pneumonia. J. Infect. Dis. 2014, 209, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Jilma, B.; Eibl, M.M. Vaccine against toxic shock syndrome in a first-in-man clinical trial. Expert Rev. Vaccines 2017, 16, 81–83. [Google Scholar] [CrossRef]


{kind=link}
| Characteristic | Number of Isolates (Total = 77) | % |
|---|---|---|
| Clinical Source | ||
| Sputum | 44 | 57.1% |
| Throat culture | 33 | 42.9% |
| Antibiotic Resistance | ||
| Oxacillin | 28 | 36.4% |
| Chloramphenicol | 0 | 0.0% |
| Tetracycline | 12 | 15.6% |
| Erythromycin | 75 | 97.4% |
| Hemolysis | ||
| Complete, rabbit blood agar ( α-toxin) | 63 | 81.8% |
| Partial, sheep blood agar (β-toxin) | 41 | 53.2% |
| Color | ||
| White | 13 | 16.9% |
| Yellow | 37 | 48.1% |
| Gold | 27 | 35.1% |
| Secreted Protease | ||
| Not detected | 41 | 53.2% |
| Faint | 13 | 16.9% |
| Present | 23 | 29.9% |
| Toxin | Iowa Subjects with CF Total = 45 | Single Culture Total = 28 | Multiple Cultures Total = 17 | P† | |||
|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | ||
| sea | 19 | 42.2% | 8 | 28.6% | 11 | 64.7% | 0.03 |
| seb | 2 | 4.4% | 1 | 3.6% | 1 | 5.9% | 1.00 |
| sec | 4 | 8.9% | 2 | 7.1% | 2 | 11.8% | 0.63 |
| sed | 6 | 13.3% | 2 | 7.1% | 4 | 23.5% | 0.18 |
| see | 3 | 6.7% | 0 | 0.0% | 3 | 17.6% | 0.05 |
| seg | 42 | 93.3% | 25 | 89.3% | 17 | 100.0% | 0.28 |
| sel-h | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | - |
| sel-i | 42 | 93.3% | 25 | 89.3% | 17 | 100.0% | 0.28 |
| sel-k | 4 | 8.9% | 2 | 7.1% | 2 | 11.8% | 0.63 |
| sel-l | 3 | 6.7% | 1 | 3.6% | 2 | 11.8% | 0.55 |
| sel-m | 39 | 86.7% | 23 | 82.1% | 16 | 94.1% | 0.38 |
| sel-n | 41 | 91.1% | 25 | 89.3% | 16 | 94.1% | 1.00 |
| sel-o | 43 | 95.6% | 26 | 92.9% | 17 | 100.0% | 0.52 |
| sel-p | 30 | 66.7% | 15 | 53.6% | 15 | 88.2% | 0.02 |
| sel-q | 15 | 33.3% | 8 | 28.6% | 7 | 41.2% | 0.52 |
| sel-u | 41 | 91.1% | 24 | 85.7% | 17 | 100.0% | 0.28 |
| sel-x | 39 | 86.7% | 23 | 82.1% | 16 | 94.1% | 0.38 |
| tstH | 8 | 17.8% | 3 | 10.7% | 5 | 29.4% | 0.23 |
| ≥1 of egc | 44 | 97.8% | 27 | 96.4% | 17 | 100.0% | 1.00 |
| 6 of egc | 37 | 82.2% | 21 | 75.0% | 16 | 94.1% | 0.13 |
| S. aureus Total = 77 | MSSA Total = 49 | MRSA Total = 28 | OR * | P† | ||||
|---|---|---|---|---|---|---|---|---|
| Toxin | N | % | N | % | N | % | ||
| sea | 25 | 32.5% | 17 | 34.7% | 8 | 28.6% | 0.76 | 0.62 |
| seb | 3 | 3.9% | 3 | 6.1% | 0 | 0.0% | - | 0.30 |
| sec | 5 | 6.5% | 5 | 10.2% | 0 | 0.0% | - | 0.15 |
| sed | 8 | 10.4% | 3 | 6.1% | 5 | 17.9% | 3.28 | 0.13 |
| see | 5 | 6.5% | 4 | 8.2% | 1 | 3.6% | 0.42 | 0.65 |
| seg | 69 | 89.6% | 45 | 91.8% | 24 | 85.7% | 0.54 | 0.45 |
| sel-h | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | - | - |
| sel-i | 69 | 89.6% | 44 | 89.8% | 25 | 89.3% | 0.95 | 1.00 |
| sel-k | 5 | 6.5% | 3 | 6.1% | 2 | 7.1% | 1.18 | 1.00 |
| sel-l | 3 | 3.9% | 3 | 6.1% | 0 | 0.0% | - | 0.30 |
| sel-m | 67 | 87.0% | 42 | 85.7% | 25 | 89.3% | 1.38 | 0.74 |
| sel-n | 68 | 88.3% | 43 | 87.8% | 25 | 89.3% | 1.16 | 1.00 |
| sel-o | 71 | 92.2% | 46 | 93.9% | 25 | 89.3% | 0.55 | 0.66 |
| sel-p | 46 | 59.7% | 34 | 69.4% | 12 | 42.9% | 0.34 | 0.03 |
| sel-q | 17 | 22.1% | 11 | 22.4% | 6 | 21.4% | 0.94 | 1.00 |
| sel-u | 69 | 89.6% | 45 | 91.8% | 24 | 85.7% | 0.54 | 0.45 |
| sel-x | 61 | 79.2% | 33 | 67.3% | 28 | 100.0% | - | 0.0003 |
| tstH | 9 | 11.7% | 9 | 18.4% | 0 | 0.0% | - | 0.02 |
| hla | 63 | 81.8% | 37 | 75.5% | 26 | 92.9% | 4.15 | 0.07 |
| hlb | 41 | 53.2% | 19 | 38.8% | 22 | 78.6% | 5.65 | 0.001 |
| Toxin | Iowa Total = 77 | Emory Total = 20 | Dartmouth Total = 12 | |||
|---|---|---|---|---|---|---|
| N | % | N | % | N | % | |
| sea | 25 | 32.5% | 0 | 0.0% | 2 | 16.7% |
| seb | 3 | 3.9% | 0 | 0.0% | 0 | 0.0% |
| sec | 5 | 6.5% | 1 | 5.0% | 0 | 0.0% |
| sed | 8 | 10.4% | 3 | 15.0% | 0 | 0.0% |
| see | 5 | 6.5% | 4 | 20.0% | 0 | 0.0% |
| seg | 69 | 89.6% | 16 | 80.0% | 10 | 83.3% |
| sel-h | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% |
| sel-i | 69 | 89.6% | 16 | 80.0% | 10 | 83.3% |
| sel-k | 5 | 6.5% | 1 | 5.0% | 2 | 16.7% |
| sel-l | 3 | 3.9% | 1 | 5.0% | 0 | 0.0% |
| sel-m | 67 | 87.0% | 15 | 75.0% | 9 | 75.0% |
| sel-n | 68 | 88.3% | 16 | 80.0% | 10 | 83.3% |
| sel-o | 71 | 92.2% | 15 | 75.0% | 9 | 75.0% |
| sel-p | 46 | 59.7% | 5 | 25.0% | 0 | 0.0% |
| sel-q | 17 | 22.1% | 2 | 10.0% | 2 | 16.7% |
| sel-u | 69 | 89.6% | 16 | 80.0% | 10 | 83.3% |
| sel-x | 61 | 79.2% | 19 | 95.0% | 11 | 91.7% |
| tstH | 9 | 11.7% | 1 | 5.0% | 2 | 16.7% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fischer, A.J.; Kilgore, S.H.; Singh, S.B.; Allen, P.D.; Hansen, A.R.; Limoli, D.H.; Schlievert, P.M. High Prevalence of Staphylococcus aureus Enterotoxin Gene Cluster Superantigens in Cystic Fibrosis Clinical Isolates. Genes 2019, 10, 1036. https://doi.org/10.3390/genes10121036
Fischer AJ, Kilgore SH, Singh SB, Allen PD, Hansen AR, Limoli DH, Schlievert PM. High Prevalence of Staphylococcus aureus Enterotoxin Gene Cluster Superantigens in Cystic Fibrosis Clinical Isolates. Genes. 2019; 10(12):1036. https://doi.org/10.3390/genes10121036
Chicago/Turabian StyleFischer, Anthony J., Samuel H. Kilgore, Sachinkumar B. Singh, Patrick D. Allen, Alexis R. Hansen, Dominique H. Limoli, and Patrick M. Schlievert. 2019. "High Prevalence of Staphylococcus aureus Enterotoxin Gene Cluster Superantigens in Cystic Fibrosis Clinical Isolates" Genes 10, no. 12: 1036. https://doi.org/10.3390/genes10121036
APA StyleFischer, A. J., Kilgore, S. H., Singh, S. B., Allen, P. D., Hansen, A. R., Limoli, D. H., & Schlievert, P. M. (2019). High Prevalence of Staphylococcus aureus Enterotoxin Gene Cluster Superantigens in Cystic Fibrosis Clinical Isolates. Genes, 10(12), 1036. https://doi.org/10.3390/genes10121036