AMH: Could It Be Used as A Biomarker for Fertility and Superovulation in Domestic Animals?

 ,
, 
 ,
,  , and
, and 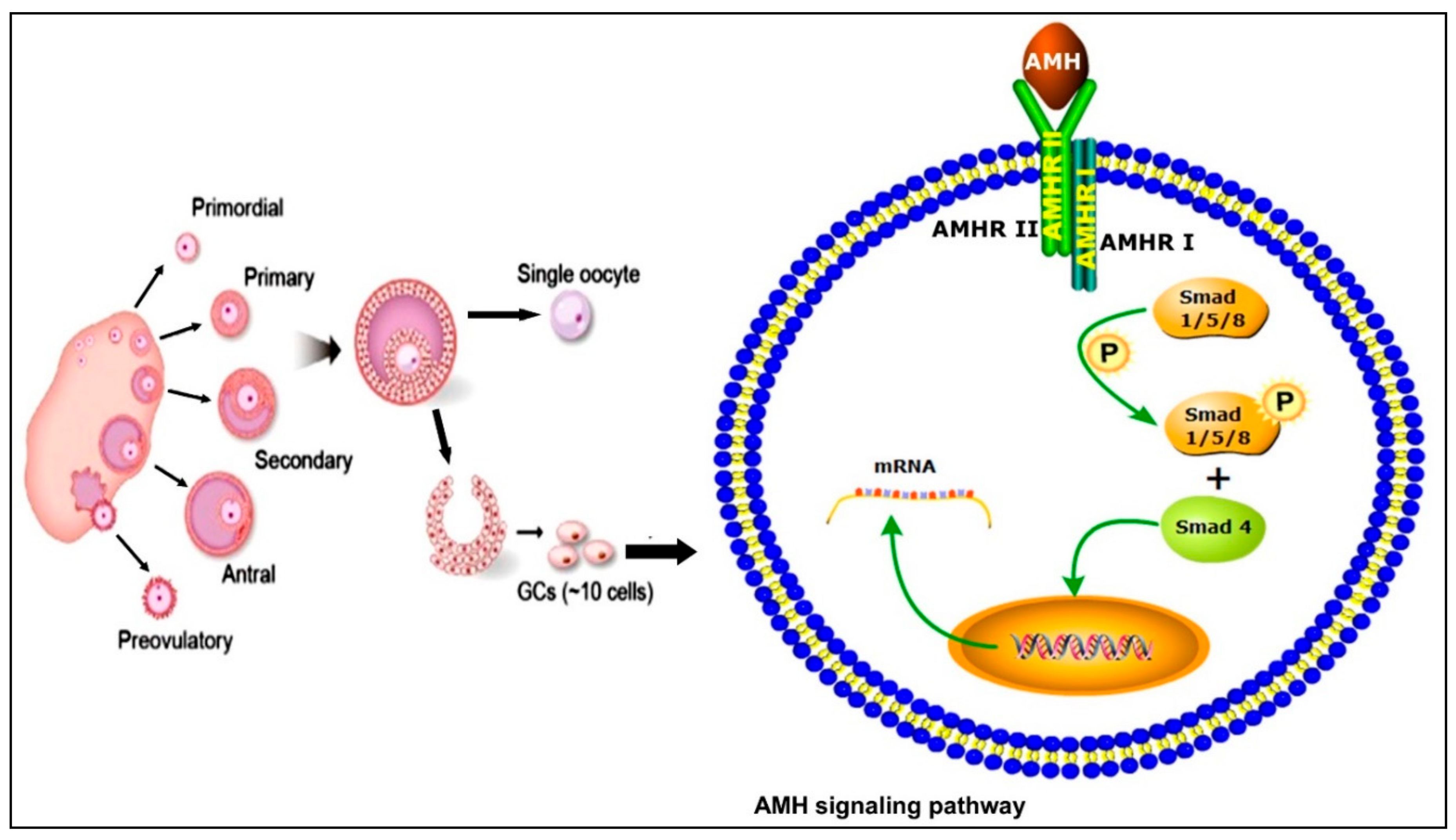{kind=link}
{kind=link}
Abstract
1. Introduction
2. Anti-Müllerian Hormone Signaling Pathways
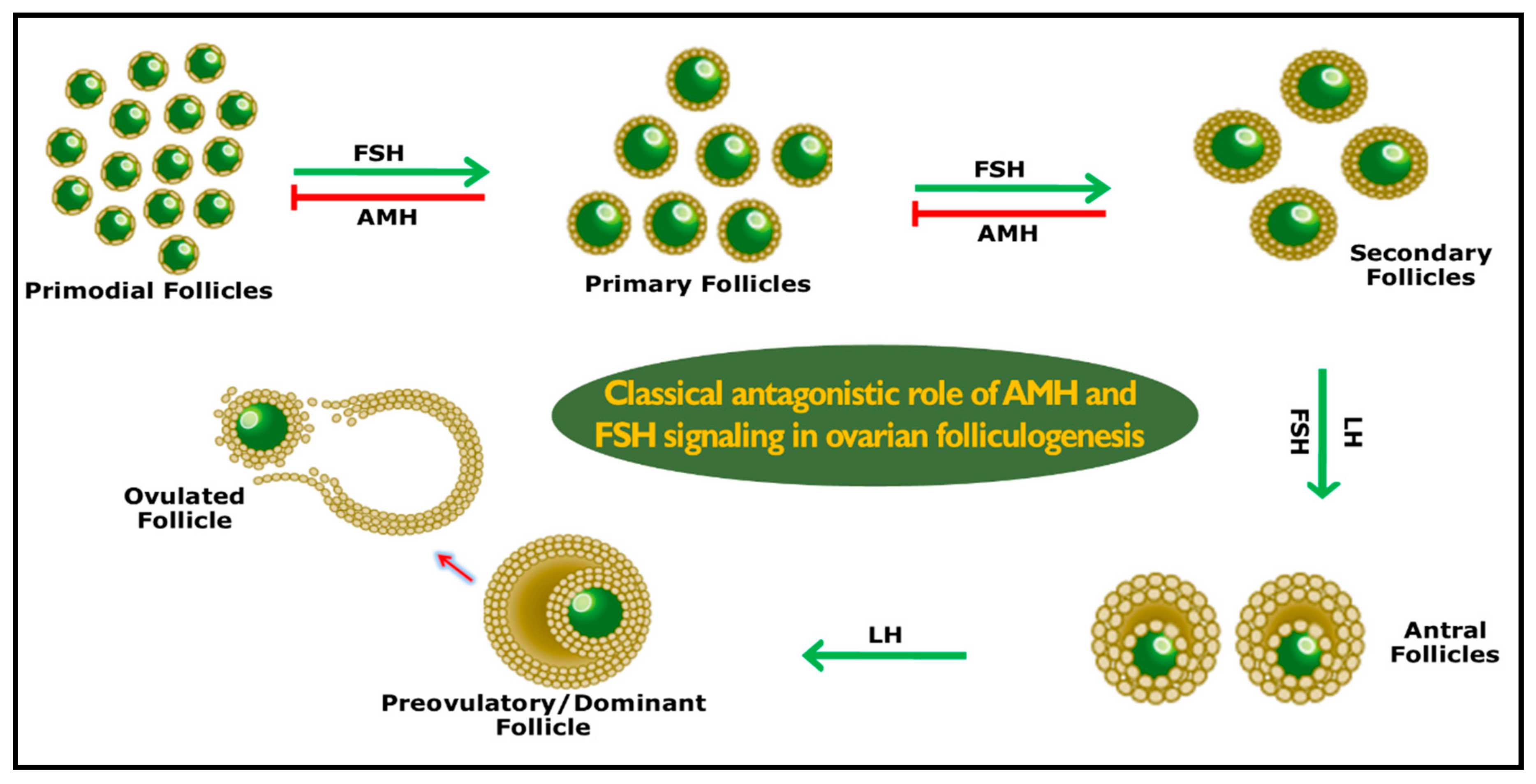
3. Role of AMH
4. How Antral Follicle Count Became a Fertility Biomarker
5. AMH Repeatability and Relationship with AFC
6. Fertility and Dynamics of AMH in Different Age Groups
7. AMH and Assisted Reproductive Technologies
8. Heritability of AMH
9. Factors Affecting AMH
9.1. Nutrition
9.2. Hormones
9.3. Disease
9.4. Granulosa Cells
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jost, A. Problems of fetal endocrinology: The gonadal and hypophyseal hormones. Recent Prog. Horm. Res. 1953, 8, 379–413. [Google Scholar]
- Picon, R. Action du testicule foetal sur le développement in vitro des canaux de Müller chez le rat. Arch. Anat. Microsc. Morphol. Exp. 1969, 58, 1–19. [Google Scholar] [PubMed]
- Picard, J.-Y.; Josso, N. Purification of testicular anti-Müllerian hormone allowing direct visualization of the pure glycoprotein and determination of yield and purification factor. Mol. Cell. Endocrinol. 1984, 34, 23–29. [Google Scholar] [CrossRef]
- Vigier, B.; Picard, J.-Y.; Tran, D.; Legeai, L.; Josso, N. Production of anti-Müllerian hormone: Another homology between Sertoli and granulosa cells. Endocrinology 1984, 114, 1315–1320. [Google Scholar] [CrossRef]
- Takahashi, M.; Hayashi, M.; Manganaro, T.F.; Donahoe, P.K. The ontogeny of mullerian inhibiting substance in granulosa-cells of the bovine ovarian follicle. Biol. Reprod. 1986, 35, 447–453. [Google Scholar] [CrossRef]
- Bézard, J.; Vigier, B.; Tran, D.; Mauleon, P.; Josso, N. Immunocytochemical study of anti-Müllerian hormone in sheep ovarian follicles during fetal and post-natal development. J. Reprod. Fertil. 1987, 80, 509–516. [Google Scholar] [CrossRef]
- Josso, N. Remembrance of Dr. Alfred Jost. Endocrinology 1991, 129, 2274–2276. [Google Scholar]
- Teixeira, J.; Maheswaran, S.; Donahoe, P.K. Mullerian inhibiting substance: An instructive developmental hormone with diagnostic and possible therapeutic applications. Endocr. Rev. 2001, 22, 657–674. [Google Scholar] [CrossRef]
- Cate, R.L.; Mattaliano, R.J.; Hession, C.; Tizard, R.; Farber, N.M.; Cheung, A.; Ninfa, E.G.; Frey, A.Z.; Gash, D.J.; Chow, E.P. Isolation of the bovine and human genes for Müllerian inhibiting substance and expression of the human gene in animal cells. Cell 1986, 45, 685–698. [Google Scholar] [CrossRef]
- Josso, N.; Di Clemente, N. Transduction pathway of anti-Müllerian hormone, a sex-specific member of the TGF-β family. Trends Endocrinol. Metab. 2003, 14, 91–97. [Google Scholar] [CrossRef]
- Claes, A.; Ball, B.A.; Almeida, J.; Corbin, C.J.; Conley, A.J. Serum anti-Müllerian hormone concentrations in stallions: Developmental changes, seasonal variation, and differences between intact stallions, cryptorchid stallions, and geldings. Theriogenology 2013, 79, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Womack, J.E. A genetic map of bovine chromosome 7 with an interspecific hybrid backcross panel. Mamm. Genome 1997, 8, 258–261. [Google Scholar] [CrossRef] [PubMed]
- NCBI. Available online: https://www.ncbi.nlm.nih.gov/pubmed (accessed on 3 April 2019).
- Monniaux, D.; Barbey, S.; Rico, C.; Fabre, S.; Gallard, Y.; Larroque, H. Anti-Müllerian hormone: A predictive marker of embryo production in cattle? Reprod. Fertil. Dev. 2010, 22, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J. TGF-β signal transduction. Annu. Rev. Biochem. 1998, 67, 753–791. [Google Scholar]
- Massagué, J.; Wotton, D. Transcriptional control by the TGF-β/Smad signaling system. EMBO J. 2000, 19, 1745–1754. [Google Scholar] [CrossRef]
- Visser, J.A. AMH signaling: From receptor to target gene. Mol. Cell. Endocrinol. 2003, 211, 65–73. [Google Scholar] [CrossRef]
- Moustakas, A.; Heldin, C.H. Non-Smad TGF-beta signals. J. Cell Sci. 2005, 118, 3573–3584. [Google Scholar] [CrossRef]
- Yi, J.Y.; Shin, I.; Arteaga, C.L. Type I transforming growth factor β receptor binds to and activates phosphatidylinositol 3-kinase. J. Biol. Chem. 2005, 280, 10870–10876. [Google Scholar] [CrossRef]
- Baarends, W.M.; Van Helmond, M.J.; Post, M.; van Der Schoot, P.J.; Hoogerbrugge, J.W.; de Winter, J.P.; Uilenbroek, J.T.; Karels, B.; Wilming, L.G.; Meijers, J.H. A novel member of the transmembrane serine/threonine kinase receptor family is specifically expressed in the gonads and in mesenchymal cells adjacent to the mullerian duct. Development 1994, 120, 189–197. [Google Scholar]
- Miyazono, K.; Kamiya, Y.; Morikawa, M. Bone morphogenetic protein receptors and signal transduction. J. Biochem. 2010, 147, 35–51. [Google Scholar] [CrossRef]
- Erickson, G.F.; Shimasaki, S. The spatiotemporal expression pattern of the bone morphogenetic protein family in rat ovary cell types during the estrous cycle. Reprod. Biol. Endocrinol. 2003, 1, 9. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, S.; Zachow, R.J.; Li, D.; Kim, H.; Iemura, S.; Ueno, N.; Sampath, K.; Chang, R.J.; Erickson, G.F. A functional bone morphogenetic protein system in the ovary. Proc. Natl. Acad. Sci. USA 1999, 96, 7282–7287. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, S.; Moore, R.K.; Otsuka, F.; Erickson, G.F. The bone morphogenetic protein system in mammalian reproduction. Endocr. Rev. 2004, 25, 72–101. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Guo, F.; Sun, X.; He, Q.; Dai, Z.; Chen, X.; Zhao, Y.; Wang, J. BMP15 regulates AMH expression via the p38 MAPK pathway in granulosa cells from goat. Theriogenology 2018, 118, 72–79. [Google Scholar] [CrossRef] [PubMed]
- McGee, E.A.; Hsueh, A.J.W. Initial and cyclic recruitment of ovarian follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar] [CrossRef] [PubMed]
- UENO, S.; Kuroda, T.; Maclaughlin, D.T.; Ragin, R.C.; Manganaro, T.F.; Donahoe, P.K. Mullerian inhibiting substance in the adult rat ovary during various stages of the estrous cycle. Endocrinology 1989, 125, 1060–1066. [Google Scholar] [CrossRef]
- Weenen, C.; Laven, J.S.E.; von Bergh, A.R.M.; Cranfield, M.; Groome, N.P.; Visser, J.A.; Kramer, P.; Fauser, B.C.J.M.; Themmen, A.P.N. Anti-Müllerian hormone expression pattern in the human ovary: Potential implications for initial and cyclic follicle recruitment. MHR Basic Sci. Reprod. Med. 2004, 10, 77–83. [Google Scholar] [CrossRef]
- Monniaux, D.; Clemente, N.D.; Touzé, J.-L.; Belville, C.; Rico, C.; Bontoux, M.; Picard, J.-Y.; Fabre, S. Intrafollicular steroids and anti-Müllerian hormone during normal and cystic ovarian follicular development in the cow. Biol. Reprod. 2008, 79, 387–396. [Google Scholar] [CrossRef]
- Veiga-Lopez, A.; Ye, W.; Padmanabhan, V. Developmental programming: Prenatal testosterone excess disrupts anti-Müllerian hormone expression in preantral and antral follicles. Fertil. Steril. 2012, 97, 748–756. [Google Scholar] [CrossRef]
- Liang, A.; Salzano, A.; D’Esposito, M.; Comin, A.; Montillo, M.; Yang, L.; Campanile, G.; Gasparrini, B. Anti-Mullerian hormone (AMH) concentration in follicular fluid and mRNA expression of AMH receptor type II and LH receptor in granulosa cells as predictive markers of good buffalo (Bubalus bubalis) donors. Theriogenology 2016, 86, 963–970. [Google Scholar] [CrossRef]
- Rocha, R.M.P.; Lima, L.F.; Carvalho, A.A.; Chaves, R.N.; Bernuci, M.P.; Rosa-e-Silva, A.C.J.S.; Rodrigues, A.P.R.; Campello, C.C.; Figueiredo, J.R. Immunolocalization of the Anti-Müllerian Hormone (AMH) in caprine follicles and the effects of AMH on in vitro culture of caprine pre-antral follicles enclosed in ovarian tissue. Reprod. Domest. Anim. 2016, 51, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Claes, A.; Ball, B.A.; Troedsson, M.H.T.; Curry, T.E.; Squires, E.L.; Scoggin, K.E. Molecular changes in the equine follicle in relation to variations in antral follicle count and anti-Müllerian hormone concentrations. Equine Vet. J. 2016, 48, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.R.C.L.; Costermans, N.G.J.; Soede, N.M.; Bunschoten, A.; Keijer, J.; Kemp, B.; Teerds, K.J. Presence of anti-Müllerian hormone (AMH) during follicular development in the porcine ovary. PLoS ONE 2018, 13, e0197894. [Google Scholar] [CrossRef] [PubMed]
- Durlinger, A.L.L.; Kramer, P.; Karels, B.; de Jong, F.H.; Uilenbroek, J.T.J.; Grootegoed, J.A.; Themmen, A.P.N. Control of primordial follicle recruitment by anti-Mullerian hormone in the mouse ovary. Endocrinology 1999, 140, 5789–5796. [Google Scholar] [CrossRef] [PubMed]
- Durlinger, A.L.L.; Gruijters, M.J.G.; Kramer, P.; Karels, B.; Kumar, T.R.; Matzuk, M.M.; Rose, U.M.; de Jong, F.H.; Uilenbroek, J.T.J.; Grootegoed, J.A. Anti-Mullerian hormone attenuates the effects of FSH on follicle development in the mouse ovary. Endocrinology 2001, 142, 4891–4899. [Google Scholar] [CrossRef]
- Pellatt, L.; Rice, S.; Dilaver, N.; Heshri, A.; Galea, R.; Brincat, M.; Brown, K.; Simpson, E.R.; Mason, H.D. Anti-Müllerian hormone reduces follicle sensitivity to follicle-stimulating hormone in human granulosa cells. Fertil. Steril. 2011, 96, 1246–1251. [Google Scholar] [CrossRef]
- Gordon, I. The bovine oestrous cycle and associated events. In Laboratory Production of Cattle Embryos; CABI: Wallingford, UK, 2009. [Google Scholar]
- Di Clemente, N.; Goxe, B.; Remy, J.J.; Cate, R.L.; Josso, N.; Vigier, B.; Salesse, R. Inhibitory effect of AMH upon aromatase activity and LH receptors of granulosa cells of rat and porcine immature ovaries. Endocrine 1994, 2, 553–558. [Google Scholar]
- Ayalon, D.; Tsafriri, A.; Lindner, H.; Cordova, T.; Harell, A. Serum gonadotropin levels in pro-oestrous rats in relation to the resumption of meiosis by the oocytes. J. Reprod. Fertil. 1972, 31, 51–58. [Google Scholar] [CrossRef][Green Version]
- Takahashi, M.; Koide, S.; Donahoe, P. Müllerian inhibiting substance as oocyte meiosis inhibitor. Mol. Cell. Endocrinol. 1986, 47, 225–234. [Google Scholar] [CrossRef]
- Dewailly, D.; Andersen, C.Y.; Balen, A.; Broekmans, F.; Dilaver, N.; Fanchin, R.; Griesinger, G.; Kelsey, T.W.; La Marca, A.; Lambalk, C. The physiology and clinical utility of anti-Müllerian hormone in women. Hum. Reprod. Update 2014, 20, 370–385. [Google Scholar] [CrossRef]
- Adams, G.P.; Matteri, R.L.; Kastelic, J.P.; Ko, J.C.H.; Ginther, O.J. Association between surges of follicle-stimulating hormone and the emergence of follicular waves in heifers. Reproduction 1992, 94, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.S.; Jimenez-Krassel, F.; Ireland, J.L.H.; Knight, P.G.; Ireland, J.J. Numbers of antral follicles during follicular waves in cattle: Evidence for high variation among animals, very high repeatability in individuals, and an inverse association with serum follicle-stimulating hormone concentrations. Biol. Reprod. 2005, 73, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Ireland, J.J.; Ward, F.; Jimenez-Krassel, F.; Ireland, J.L.H.; Smith, G.W.; Lonergan, P.; Evans, A.C.O. Follicle numbers are highly repeatable within individual animals but are inversely correlated with FSH concentrations and the proportion of good-quality embryos after ovarian stimulation in cattle. Hum. Reprod. 2007, 22, 1687–1695. [Google Scholar] [CrossRef] [PubMed]
- Ireland, J.L.H.; Scheetz, D.; Jimenez-Krassel, F.; Themmen, A.P.N.; Ward, F.; Lonergan, P.; Smith, G.W.; Perez, G.I.; Evans, A.C.O.; Ireland, J.J. Antral follicle count reliably predicts number of morphologically healthy oocytes and follicles in ovaries of young adult cattle. Biol. Reprod. 2008, 79, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Haughian, J.M.; Ginther, O.J.; Kot, K.; Wiltbank, M.C. Relationships between FSH patterns and follicular dynamics and the temporal associations among hormones in natural and GnRH-induced gonadotropin surges in heifers. Reproduction 2004, 127, 23–33. [Google Scholar] [CrossRef]
- Mossa, F.; Jimenez-Krassel, F.; Folger, J.K.; Ireland, J.L.H.; Smith, G.W.; Lonergan, P.; Evans, A.C.O.; Ireland, J.J. Evidence that high variation in antral follicle count during follicular waves is linked to alterations in ovarian androgen production in cattle. Reproduction 2010, 140, 713. [Google Scholar] [CrossRef]
- Singh, J.; Domínguez, M.; Jaiswal, R.; Adams, G.P. A simple ultrasound test to predict the superstimulatory response in cattle. Theriogenology 2004, 62, 227–243. [Google Scholar] [CrossRef]
- Jimenez-Krassel, F.; Folger, J.K.; Ireland, J.L.H.; Smith, G.W.; Hou, X.; Davis, J.S.; Lonergan, P.; Evans, A.C.O.; Ireland, J.J. Evidence that high variation in ovarian reserves of healthy young adults has a negative impact on the corpus luteum and endometrium during estrous cycles in cattle. Biol. Reprod. 2009, 80, 1272–1281. [Google Scholar] [CrossRef]
- Diskin, M.G.; Morris, D.G. Embryonic and early foetal losses in cattle and other ruminants. Reprod. Domest. Anim. 2008, 43, 260–267. [Google Scholar] [CrossRef]
- Basir, G.S.; So, W.W.K.; Ng, E.H.Y.; Ho, P.C. Evaluation of cycle-to-cycle variation of endometrial responsiveness using transvaginal sonography in women undergoing assisted reproduction. Ultrasound Obstet. Gynecol. 2002, 19, 484–489. [Google Scholar] [CrossRef]
- Scheetz, D.; Folger, J.K.; Smith, G.W.; Ireland, J.J. Granulosa cells are refractory to FSH action in individuals with a low antral follicle count. Reprod. Fertil. Dev. 2012, 24, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Ireland, J.J.; Zielak-Steciwko, A.E.; Jimenez-Krassel, F.; Folger, J.; Bettegowda, A.; Scheetz, D.; Walsh, S.; Mossa, F.; Knight, P.G.; Smith, G.W. Variation in the ovarian reserve is linked to alterations in intrafollicular estradiol production and ovarian biomarkers of follicular differentiation and oocyte quality in cattle. Biol. Reprod. 2009, 80, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Mossa, F.; Walsh, S.W.; Butler, S.T.; Berry, D.P.; Carter, F.; Lonergan, P.; Smith, G.W.; Ireland, J.J.; Evans, A.C.O. Low numbers of ovarian follicles ≥ 3 mm in diameter are associated with low fertility in dairy cows. J. Dairy Sci. 2012, 95, 2355–2361. [Google Scholar] [CrossRef] [PubMed]
- El-Sheikh Ali, H.; Kitahara, G.; Nibe, K.; Yamaguchi, R.; Horii, Y.; Zaabel, S.; Osawa, T. Plasma anti-Müllerian hormone as a biomarker for bovine granulosa-theca cell tumors: Comparison with immunoreactive inhibin and ovarian steroid concentrations. Theriogenology 2013, 80, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Ireland, J.J.; Smith, G.W.; Scheetz, D.; Jimenez-Krassel, F.; Folger, J.K.; Ireland, J.L.H.; Mossa, F.; Lonergan, P.; Evans, A.C.O. Does size matter in females? An overview of the impact of the high variation in the ovarian reserve on ovarian function and fertility, utility of anti-Müllerian hormone as a diagnostic marker for fertility and causes of variation in the ovarian reserve in. Reprod. Fertil. Dev. 2010, 23, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.; Fabre, S.; Médigue, C.; Clemente, N.D.; Clément, F.; Bontoux, M.; Touzé, J.-L.; Dupont, M.; Briant, E.; Rémy, B. Anti-Müllerian hormone is an endocrine marker of ovarian gonadotropin-responsive follicles and can help to predict superovulatory responses in the cow. Biol. Reprod. 2009, 80, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.H.; Carvalho, P.D.; Rozner, A.E.; Vieira, L.M.; Hackbart, K.S.; Bender, R.W.; Dresch, A.R.; Verstegen, J.P.; Shaver, R.D.; Wiltbank, M.C. Relationship between circulating anti-Müllerian hormone (AMH) and superovulatory response of high-producing dairy cows. J. Dairy Sci. 2015, 98, 169–178. [Google Scholar] [CrossRef]
- Pfeiffer, K.E.; Jury, L.J.; Larson, J.E. Determination of anti-Müllerian hormone at estrus during a synchronized and a natural bovine estrous cycle. Domest. Anim. Endocrinol. 2014, 46, 58–64. [Google Scholar] [CrossRef]
- Nabenishi, H.; Kitahara, G.; Takagi, S.; Yamazaki, A.; Osawa, T. Relationship between plasma anti-Müllerian hormone concentrations during the rearing period and subsequent embryo productivity in Japanese black cattle. Domest. Anim. Endocrinol. 2017, 60, 19–24. [Google Scholar] [CrossRef]
- Hirayama, H.; Naito, A.; Fujii, T.; Sugimoto, M.; Takedomi, T.; Moriyasu, S.; Sakai, H.; Kageyama, S. Effects of genetic background on responses to superovulation in Japanese Black cattle. J. Vet. Med. Sci. 2019, 81, 373–378. [Google Scholar] [CrossRef]
- Claes, A.; Ball, B.A.; Scoggin, K.E.; Esteller-Vico, A.; Kalmar, J.J.; Conley, A.J.; Squires, E.L.; Troedsson, M.H.T. The interrelationship between anti-Müllerian hormone, ovarian follicular populations and age in mares. Equine Vet. J. 2015, 47, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Waheeb, R.S. Changes in serum levels of anti-mullerian hormone and ovarian steroids in Barki sheep during follicular and early luteal phase of estrous cycle. Pak. Vet. J. 2017, 37, 421–426. [Google Scholar]
- Santa Cruz, R.; Cushman, R.A.; Viñoles, C. Antral follicular count is a tool that may allow the selection of more precocious Bradford heifers at weaning. Theriogenology 2018, 119, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Baldrighi, J.M.; Sá Filho, M.F.D.; Batista, E.D.O.S.; Lopes, R.N.V.R.; Visintin, J.A.; Baruselli, P.S.; Assumpção, M.E.O.A. Anti-Mullerian Hormone Concentration and Antral Ovarian Follicle Population in M urrah H eifers Compared to Holstein and G yr Kept Under the Same Management. Reprod. Domest. Anim. 2014, 49, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.S.; Mahboubi, K.; Costa, N.N.; Gillis, D.J.; Carter, T.F.; Neal, M.S.; Miranda, M.S.; Ohashi, O.M.; Favetta, L.A.; King, W.A. Systemic and local anti-Mullerian hormone reflects differences in the reproduction potential of Zebu and European type cattle. Anim. Reprod. Sci. 2016, 167, 51–58. [Google Scholar] [CrossRef]
- Lahoz, B.; Alabart, J.L.; Cocero, M.J.; Monniaux, D.; Echegoyen, E.; Sánchez, P.; Folch, J. Anti-Müllerian hormone concentration in sheep and its dependence of age and independence of BMP15 genotype: An endocrine predictor to select the best donors for embryo biotechnologies. Theriogenology 2014, 81, 347–357. [Google Scholar] [CrossRef]
- Torres-Rovira, L.; Succu, S.; Pasciu, V.; Manca, M.E.; Gonzalez-Bulnes, A.; Leoni, G.G.; Pennino, M.G.; Spezzigu, A.; Gallus, M.; Dattena, M.; et al. Postnatal pituitary and follicular activation: A revisited hypothesis in a sheep model. Reproduction 2016, 151, 215–225. [Google Scholar] [CrossRef]
- Monniaux, D.; Drouilhet, L.; Rico, C.; Estienne, A.; Jarrier, P.; Touzé, J.-L.; Sapa, J.; Phocas, F.; Dupont, J.; Dalbiès-Tran, R. Regulation of anti-Müllerian hormone production in domestic animals. Reprod. Fertil. Dev. 2012, 25, 1–16. [Google Scholar] [CrossRef]
- Ball, B.A.; El-Sheikh Ali, H.; Scoggin, K.E.; Riddle, W.T.; Schnobrich, M.; Bradekamp, E.; Agnew, M.; Squires, E.L.; Troedsson, M.H.T. Relationship between anti-Müllerian hormone and fertility in the mare. Theriogenology 2019, 125, 335–341. [Google Scholar] [CrossRef]
- Uliani, R.C.; Conley, A.J.; Jo Corbin, C.; Friso, A.M.; Maciel, L.F.S.; Alvarenga, M.A. Anti-Müllerian hormone and ovarian aging in mares. J. Endocrinol. 2019, 240, 147–156. [Google Scholar] [CrossRef]
- Evans, A.C.O.; Adams, G.P.; Rawling, N.C. Follicular and hormonal development in prepubertal heifers from 2 to 36 weeks of age. Reproduction 1994, 102, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.E.S.; Kitahara, G.; Takahashi, T.; Mido, S.; Sadawy, M.; Kobayashi, I.; Hemmi, K.; Osawa, T. Plasma anti-Müllerian hormone profile in heifers from birth through puberty and relationship with puberty onset. Biol. Reprod. 2017, 97, 153–161. [Google Scholar]
- Wehrman, M.E.; Kojima, F.N.; Sanchez, T.; Mariscal, D.V.; Kinder, J.E. Incidence of precocious puberty in developing beef heifers. J. Anim. Sci. 1996, 74, 2462–2467. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Buskirk, D.D.; Faulkner, D.B.; Ireland, F.A. Increased postweaning gain of beef heifers enhances fertility and milk production. J. Anim. Sci. 1995, 73, 937–946. [Google Scholar] [CrossRef]
- Bagley, C.P. Nutritional management of replacement beef heifers: A review. J. Anim. Sci. 1993, 71, 3155–3163. [Google Scholar] [CrossRef]
- Lesmeister, J.L.; Burfening, P.J.; Blackwell, R.L. Date of first calving in beef cows and subsequent calf production. J. Anim. Sci. 1973, 36, 1–6. [Google Scholar] [CrossRef]
- Jimenez-Krassel, F.; Scheetz, D.M.; Neuder, L.M.; Ireland, J.L.H.; Pursley, J.R.; Smith, G.W.; Tempelman, R.J.; Ferris, T.; Roudebush, W.E.; Mossa, F. Concentration of anti-Müllerian hormone in dairy heifers is positively associated with productive herd life. J. Dairy Sci. 2015, 98, 3036–3045. [Google Scholar] [CrossRef]
- Mossa, F.; Jimenez-Krassel, F.; Walsh, S.; Berry, D.P.; Butler, S.T.; Folger, J.; Smith, G.W.; Ireland, J.L.H.; Lonergan, P.; Ireland, J.J. Inherent capacity of the pituitary gland to produce gonadotropins is not influenced by the number of ovarian follicles ≥ 3 mm in diameter in cattle. Reprod. Fertil. Dev. 2010, 22, 550–557. [Google Scholar] [CrossRef]
- Ribeiro, E.S.; Bisinotto, R.S.; Lima, F.S.; Greco, L.F.; Morrison, A.; Kumar, A.; Thatcher, W.W.; Santos, J.E.P. Plasma anti-Müllerian hormone in adult dairy cows and associations with fertility. J. Dairy Sci. 2014, 97, 6888–6900. [Google Scholar] [CrossRef]
- Lei, G.; Wu, C.; Dai, J.; Li, Y.; Lu, Y.; Chen, X.; Zhang, S.; Zhang, D. Effects of inhibin immunization on superovulation in Chongming white goats. Acta Agric. Shanghai 2018, 34, 96–100. [Google Scholar]
- KAWAMATA, M. Relationships between the number of small follicles prior to superovulatory treatment and superovulatory response in Holstein cows. J. Vet. Med. Sci. 1994, 56, 965–967. [Google Scholar] [CrossRef] [PubMed]
- Cushman, R.A.; Allan, M.F.; Kuehn, L.A.; Snelling, W.M.; Cupp, A.S.; Freetly, H.C. Evaluation of antral follicle count and ovarian morphology in crossbred beef cows: Investigation of influence of stage of the estrous cycle, age, and birth weight. J. Anim. Sci. 2009, 87, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Taneja, M.; Bols, P.E.J.; de Velde, A.V.; Ju, J.-C.; Schreiber, D.; Tripp, M.W.; Levine, H.; Echelard, Y.; Riesen, J.; Yang, X. Developmental competence of juvenile calf oocytes in vitro and in vivo: Influence of donor animal variation and repeated gonadotropin stimulation. Biol. Reprod. 2000, 62, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.; Drouilhet, L.; Salvetti, P.; Dalbiès-Tran, R.; Jarrier, P.; Touzé, J.-L.; Pillet, E.; Ponsart, C.; Fabre, S.; Monniaux, D. Determination of anti-Müllerian hormone concentrations in blood as a tool to select Holstein donor cows for embryo production: From the laboratory to the farm. Reprod. Fertil. Dev. 2012, 24, 932–944. [Google Scholar] [CrossRef]
- Hirayama, H.; Kageyama, S.; Naito, A.; Fukuda, S.; Fujii, T.; Minamihashi, A. Prediction of superovulatory response in Japanese Black cattle using ultrasound, plasma anti-Müllerian hormone concentrations and polymorphism in the ionotropic glutamate receptor AMPA1/GRIA. J. Reprod. Dev. 2012, 58, 380–383. [Google Scholar] [CrossRef]
- Guerreiro, B.M.; Batista, E.O.S.; Vieira, L.M.; Sá Filho, M.F.; Rodrigues, C.A.; Netto, A.C.; Silveira, C.R.A.; Bayeux, B.M.; Dias, E.A.R.; Monteiro, F.M. Plasma anti-mullerian hormone: An endocrine marker for in vitro embryo production from Bos taurus and Bos indicus donors. Domest. Anim. Endocrinol. 2014, 49, 96–104. [Google Scholar] [CrossRef]
- Ghanem, N.; Jin, J.I.; Kim, S.S.; Choi, B.H.; Lee, K.L.; Ha, A.N.; Song, S.H.; Kong, I.K. The anti-Müllerian hormone profile is linked with the in vitro embryo production capacity and embryo viability after transfer but cannot predict pregnancy outcome. Reprod. Domest. Anim. 2016, 51, 301–310. [Google Scholar] [CrossRef]
- Vernunft, A.; Schwerhoff, M.; Viergutz, T.; Diederich, M.; Kuwer, A. Anti-Muellerian hormone levels in plasma of Holstein-Friesian heifers as a predictive parameter for ovum pick-up and embryo production outcomes. J. Reprod. Dev. 2015, 61, 74–79. [Google Scholar] [CrossRef]
- Monniaux, D.; Baril, G.; Laine, A.L.; Jarrier, P.; Poulin, N.; Cognieánd, J.; Fabre, S. Anti-Müllerian hormone as a predictive endocrine marker for embryo production in the goat. Reproduction 2011, 142, 845–854. [Google Scholar] [CrossRef]
- Pinto, P.H.N.; Balaro, M.F.A.; Souza-Fabjan, J.M.G.; dos Santos Ribeiro, L.; Bragança, G.M.; Leite, C.R.; Arashiro, E.K.N.; de Moraes Silva, K.; Da Fonseca, J.F.; Brandão, F.Z. Anti-Müllerian hormone and antral follicle count are more effective for selecting ewes with good potential for in vivo embryo production than the presence of FecGE mutation or eCG pre-selection tests. Theriogenology 2018, 113, 146–152. [Google Scholar]
- Berry, D.P.; Wall, E.; Pryce, J.E. Genetics and genomics of reproductive performance in dairy and beef cattle. Animal 2014, 8, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.Y.; Jimenez-Krassel, F.; Steibel, J.P.; Lu, Y.; Baktula, A.; Vukasinovic, N.; Neuder, L.; Ireland, J.L.H.; Ireland, J.J.; Tempelman, R.J. Genomic heritability and genome-wide association analysis of anti-Müllerian hormone in Holstein dairy heifers. J. Dairy Sci. 2018, 101, 8063–8075. [Google Scholar] [CrossRef] [PubMed]
- Gobikrushanth, M.; Purfield, D.C.; Colazo, M.G.; Butler, S.T.; Wang, Z.; Ambrose, D.J. The relationship between serum anti-Müllerian hormone concentrations and fertility, and genome-wide associations for anti-Müllerian hormone in Holstein cows. J. Dairy Sci. 2018, 101, 7563–7574. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.W.; Mossa, F.; Butler, S.T.; Berry, D.P.; Scheetz, D.; Jimenez-Krassel, F.; Tempelman, R.J.; Carter, F.; Lonergan, P.; Evans, A.C.O. Heritability and impact of environmental effects during pregnancy on antral follicle count in cattle. J. Dairy Sci. 2014, 97, 4503–4511. [Google Scholar] [CrossRef] [PubMed]
- Jaton, C.; Schenkel, F.S.; Sargolzaei, M.; Cánova, A.; Malchiodi, F.; Price, C.A.; Baes, C.; Miglior, F. Genome-wide association study and in silico functional analysis of the number of embryos produced by Holstein donors. J. Dairy Sci. 2018, 101, 7248–7257. [Google Scholar] [CrossRef] [PubMed]
- Erickson, B.H. Development and senescence of the postnatal bovine ovary. J. Anim. Sci. 1966, 25, 800–805. [Google Scholar] [CrossRef] [PubMed]
- Mossa, F.; Carter, F.; Walsh, S.W.; Kenny, D.A.; Smith, G.W.; Ireland, J.L.H.; Hildebrandt, T.B.; Lonergan, P.; Ireland, J.J.; Evans, A.C.O. Maternal undernutrition in cows impairs ovarian and cardiovascular systems in their offspring. Biol. Reprod. 2013, 88, 1–9, 92. [Google Scholar] [CrossRef]
- Sullivan, T.M.; Micke, G.C.; Greer, R.M.; Perry, V.E.A. Dietary manipulation of Bos indicus × heifers during gestation affects the prepubertal reproductive development of their bull calves. Anim. Reprod. Sci. 2010, 118, 131–139. [Google Scholar] [CrossRef]
- Padmanabhan, V.; Veiga-Lopez, A. Reproduction symposium: Developmental programming of reproductive and metabolic health. J. Anim. Sci. 2014, 92, 3199–3210. [Google Scholar] [CrossRef][Green Version]
- El-Sheikh Ali, H.; Kitahara, G.; Nibe, K.; Osawa, T. Endocrinological characterization of an ovarian sex cord–stromal tumor with a Sertoli cell pattern in a Japanese Black cow. Reprod. Domest. Anim. 2019, 54, 1501–1504. [Google Scholar] [CrossRef]
- Kitahara, G.; Nambo, Y.; El-Sheikh Ali, H.; Kajisa, M.; Tani, M.; Nibe, K.; Kamimura, S. Anti-Müllerian hormone profiles as a novel biomarker to diagnose granulosa-theca cell tumors in cattle. J. Reprod. Dev. 2012, 58, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Ball, B.A.; Almeida, J.; Conley, A.J. Determination of serum anti-Müllerian hormone concentrations for the diagnosis of granulosa-cell tumours in mares. Equine Vet. J. 2013, 45, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.; Ball, B.A.; Conley, A.J.; Place, N.J.; Liu, I.K.M.; Scholtz, E.L.; Mathewson, L.; Stanley, S.D.; Moeller, B.C. Biological and clinical significance of anti-Müllerian hormone determination in blood serum of the mare. Theriogenology 2011, 76, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Walter, B.; Coelfen, A.; Jäger, K.; Reese, S.; Meyer-Lindenberg, A.; Aupperle-Lellbach, H. Anti-Muellerian hormone concentration in bitches with histopathologically diagnosed ovarian tumours and cysts. Reprod. Domest. Anim. 2018, 53, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Heaps, L.A.; Scudder, C.J.; Lipscomb, V.J.; Steinbach, S.M.; Priestnall, S.L.; Martineau, H.; Szladovits, B.; Fowkes, R.C.; Garden, O.A. Serum anti-Müllerian hormone concentrations before and after treatment of an ovarian granulosa cell tumour in a cat. J. Feline Med. Surg. Open Rep. 2017, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- El-Sheikh Ali, H.; Kitahara, G.; Torisu, S.; Nibe, K.; Kaneko, Y.; Hidaka, Y.; Osawa, T. Evidence of spontaneous recovery of granulosa-theca cell tumour in a Heifer: A retrospective report. Reprod. Domest. Anim. 2015, 50, 696–704. [Google Scholar] [CrossRef]
- Bungum, L.; Franssohn, F.; Bungum, M.; Humaidan, P.; Giwercman, A. The circadian variation in anti-müllerian hormone in patients with polycystic ovary syndrome differs significantly from normally ovulating women. PLoS ONE 2013, 8, e68223. [Google Scholar] [CrossRef]
- Díaz, P.U.; Rey, F.; Gareis, N.C.; Notaro, U.S.; Matiller, V.; Belotti, E.M.; Stassi, A.F.; Salvetti, N.R.; Ortega, H.H. Altered expression of anti-Müllerian hormone during the early stage of bovine persistent ovarian follicles. J. Comp. Pathol. 2018, 158, 22–31. [Google Scholar] [CrossRef]
- Caraviello, D.Z.; Weigel, K.A.; Shook, G.E.; Ruegg, P.L. Assessment of the impact of somatic cell count on functional longevity in Holstein and Jersey cattle using survival analysis methodology. J. Dairy Sci. 2005, 88, 804–811. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umer, S.; Zhao, S.J.; Sammad, A.; Weldegebriall Sahlu, B.; Pang, Y.; Zhu, H. AMH: Could It Be Used as A Biomarker for Fertility and Superovulation in Domestic Animals? Genes 2019, 10, 1009. https://doi.org/10.3390/genes10121009
Umer S, Zhao SJ, Sammad A, Weldegebriall Sahlu B, Pang Y, Zhu H. AMH: Could It Be Used as A Biomarker for Fertility and Superovulation in Domestic Animals? Genes. 2019; 10(12):1009. https://doi.org/10.3390/genes10121009
Chicago/Turabian StyleUmer, Saqib, Shan Jiang Zhao, Abdul Sammad, Bahlibi Weldegebriall Sahlu, YunWei Pang, and Huabin Zhu. 2019. "AMH: Could It Be Used as A Biomarker for Fertility and Superovulation in Domestic Animals?" Genes 10, no. 12: 1009. https://doi.org/10.3390/genes10121009
APA StyleUmer, S., Zhao, S. J., Sammad, A., Weldegebriall Sahlu, B., Pang, Y., & Zhu, H. (2019). AMH: Could It Be Used as A Biomarker for Fertility and Superovulation in Domestic Animals? Genes, 10(12), 1009. https://doi.org/10.3390/genes10121009