ZW Sex Chromosomes in Australian Dragon Lizards (Agamidae) Originated from a Combination of Duplication and Translocation in the Nucleolar Organising Region

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Sexing, Cell Culture and Chromosome Preparations
2.2. Probe Preparation and Fluorescence in Situ Hybridisation (FISH)
2.3. BLASTn Analysis of PviZW BAC (Pv151P16)
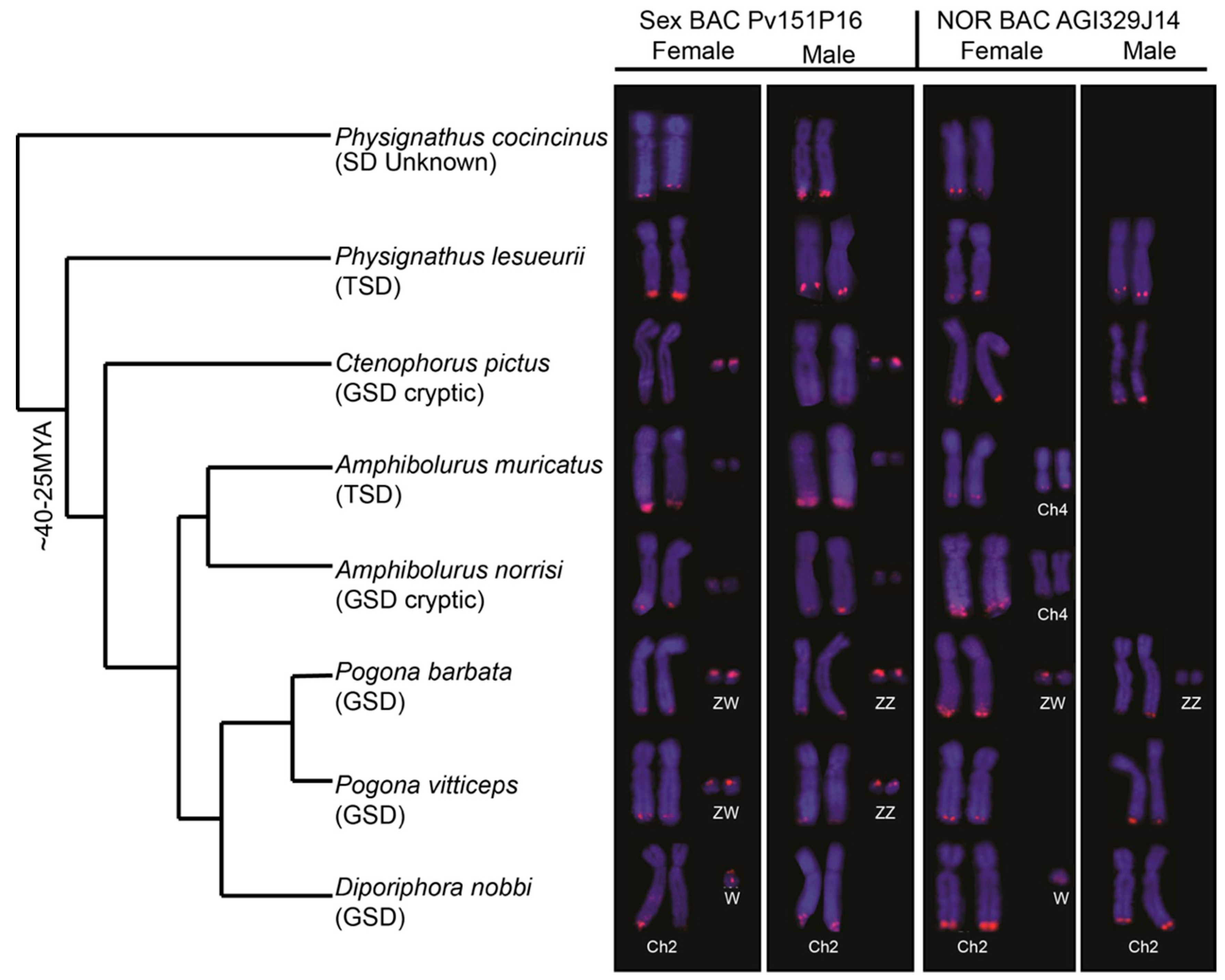
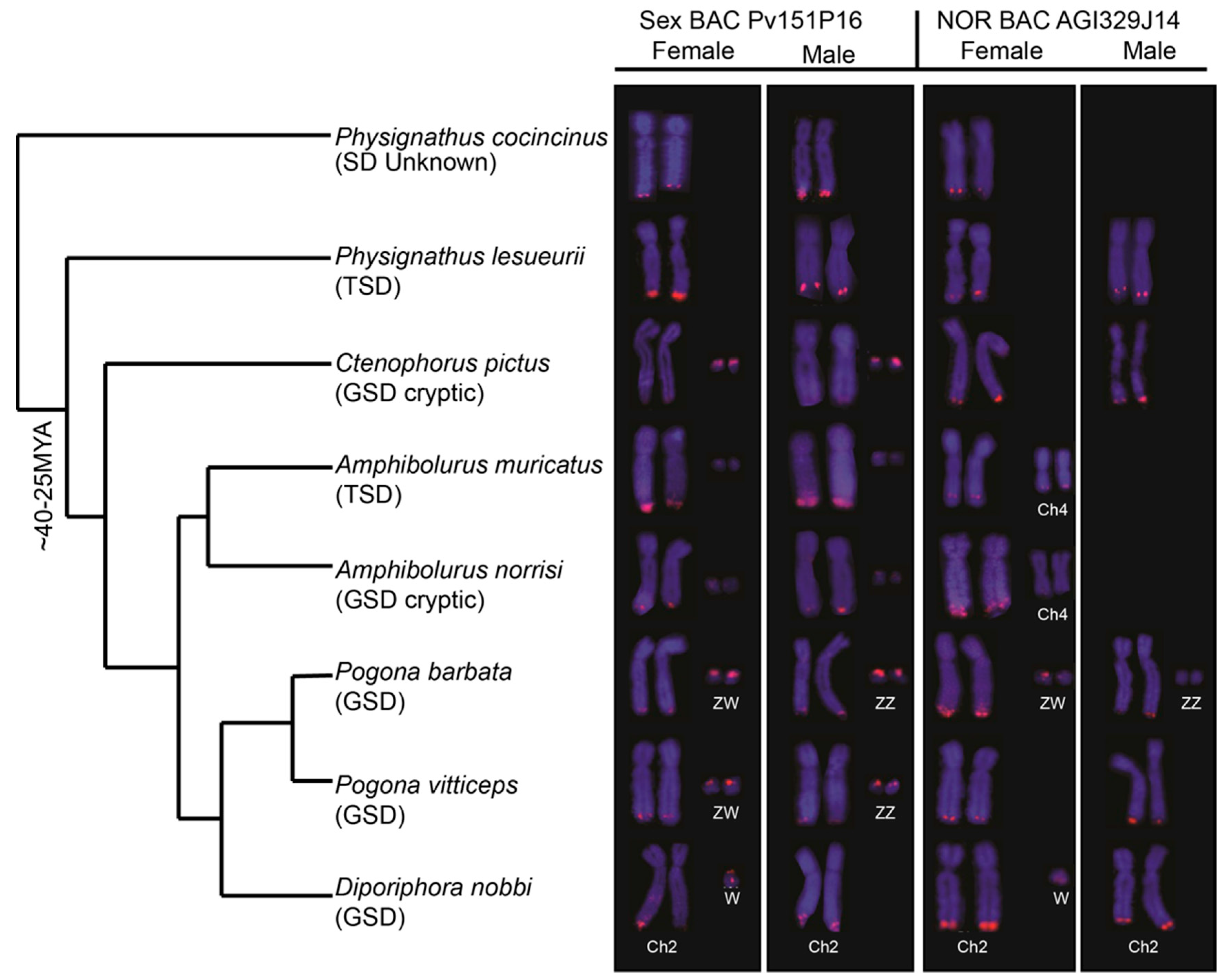
3. Results
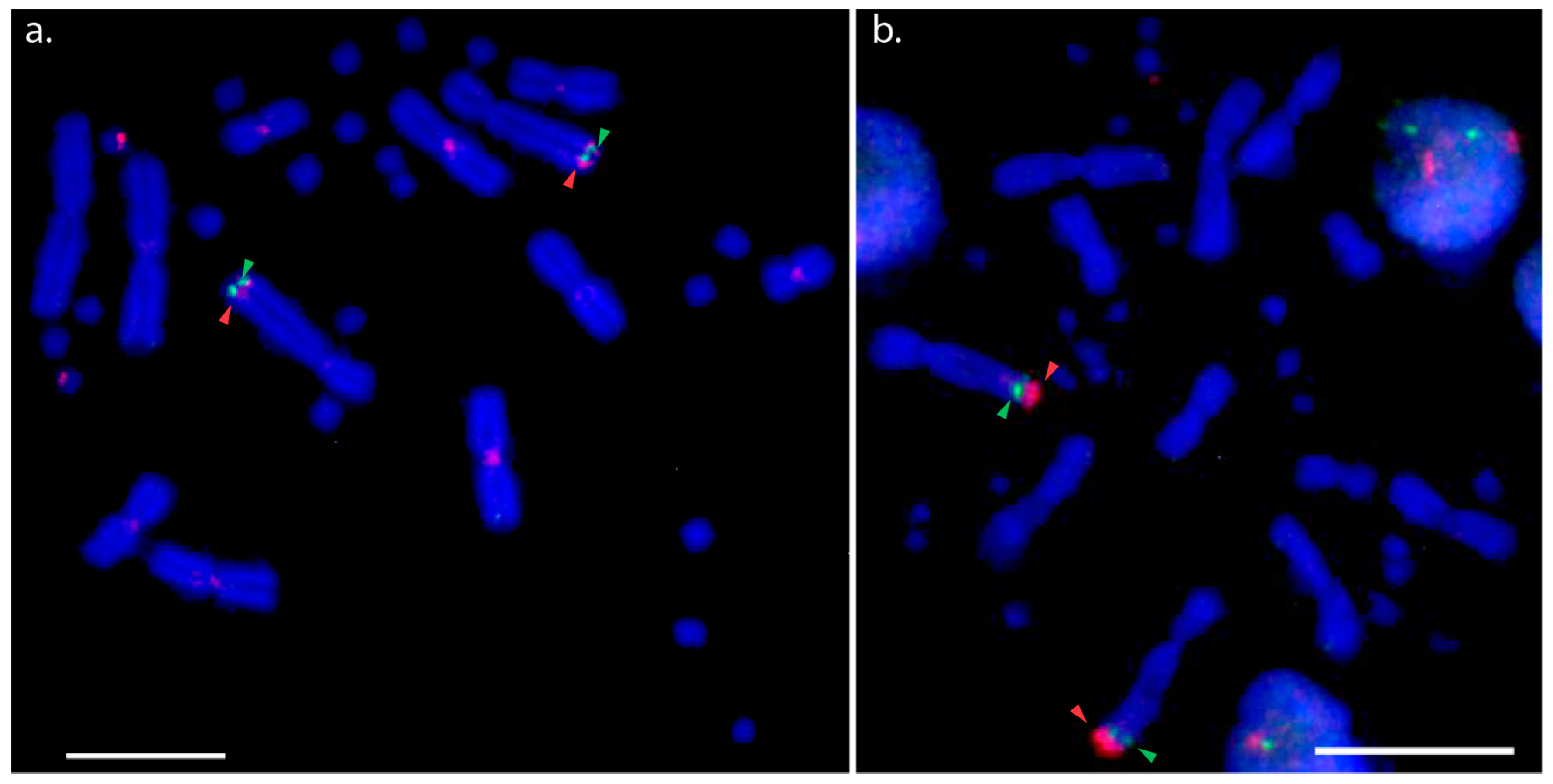
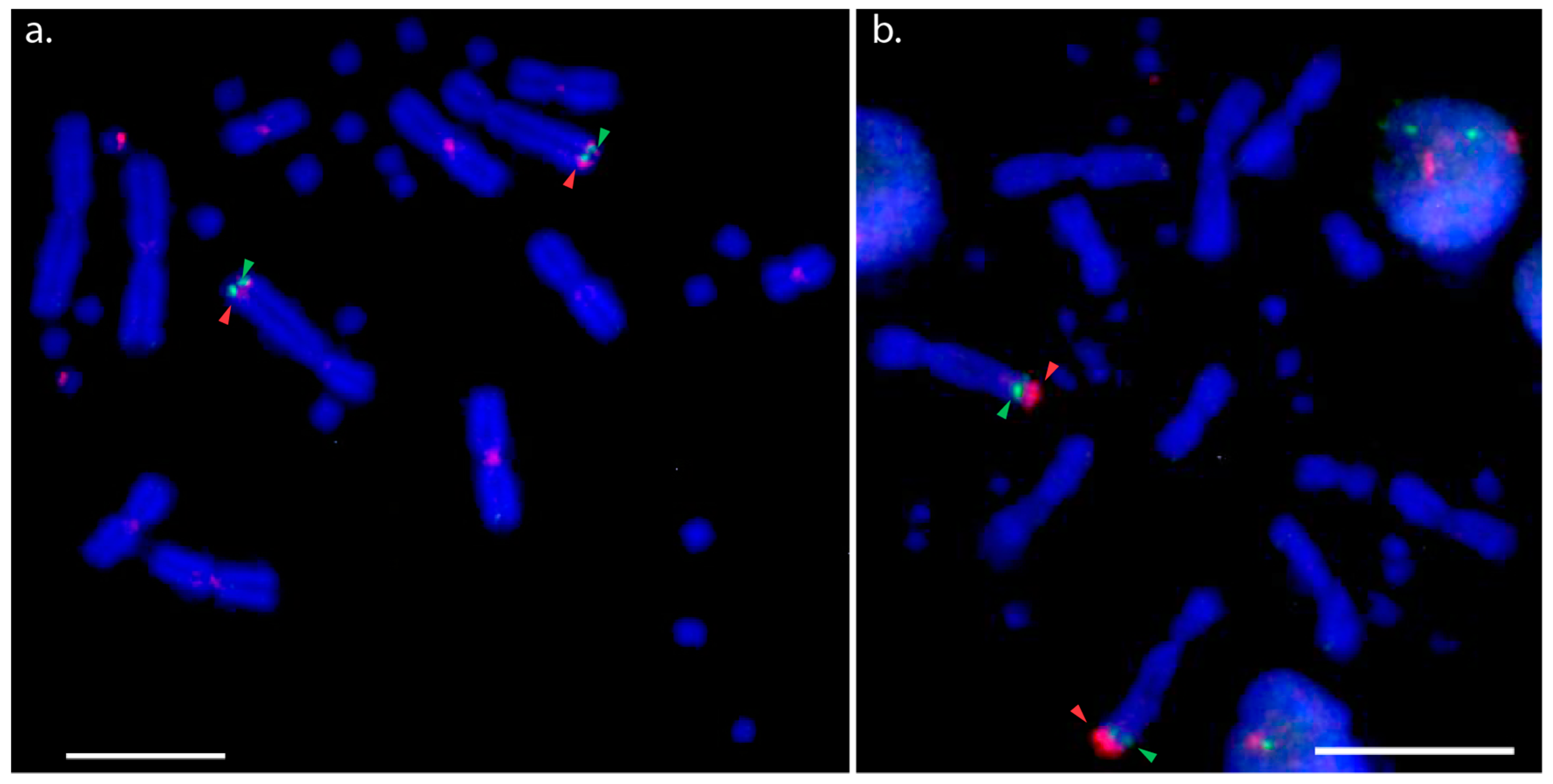
3.1. Physical Mapping of PviZW BAC
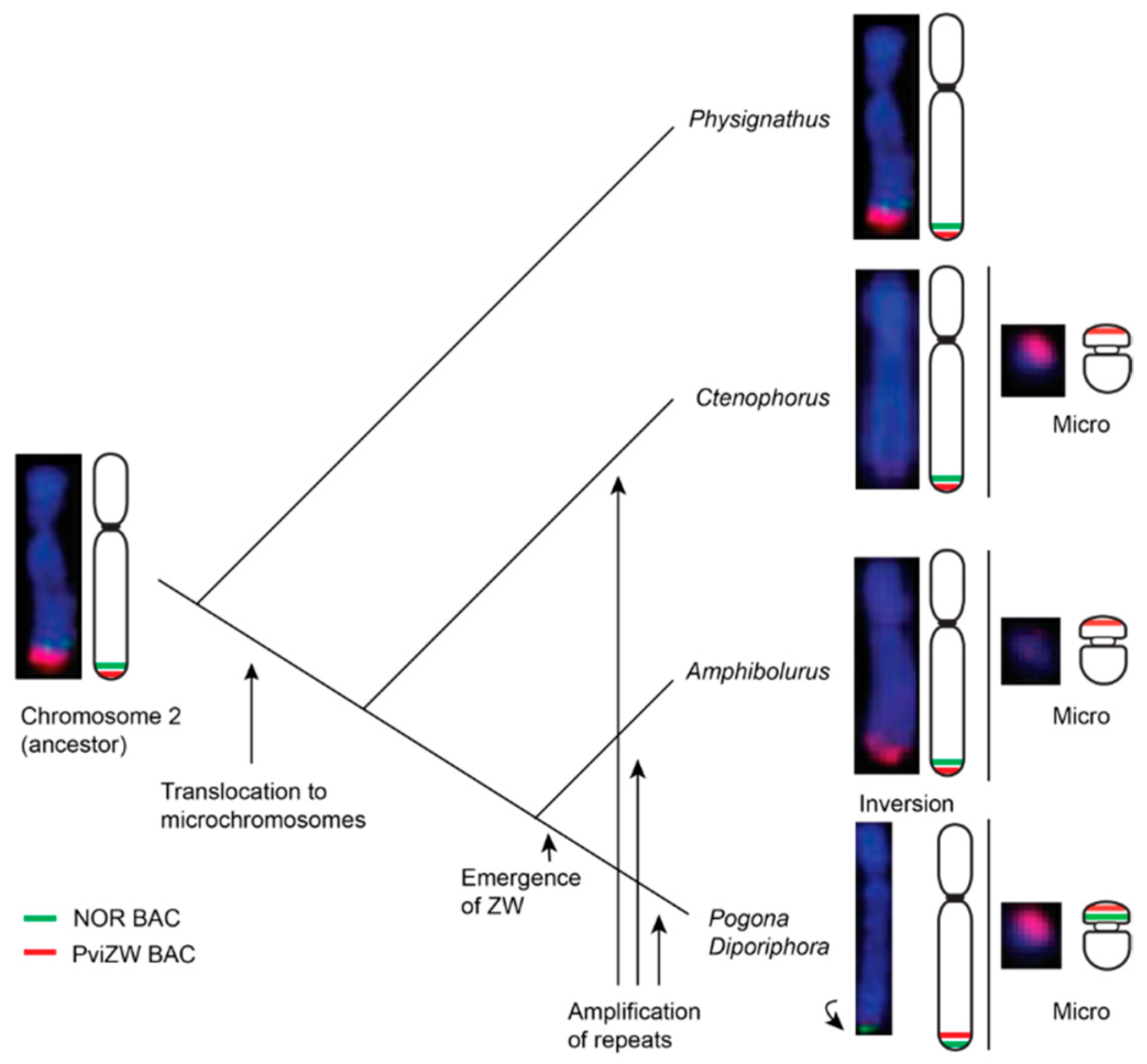
3.2. Physical Mapping of 18S–28S rDNA (NOR) Genes
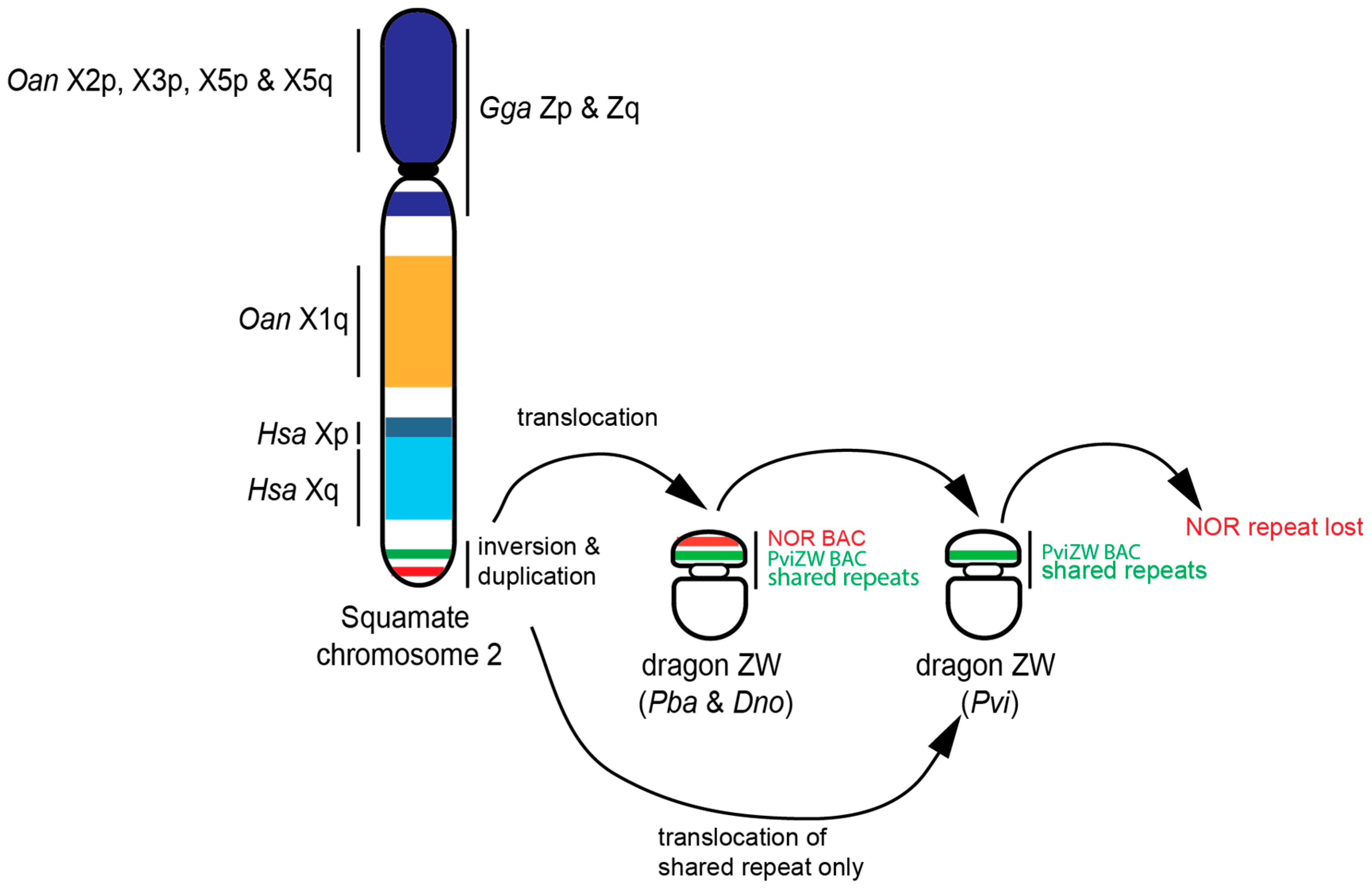
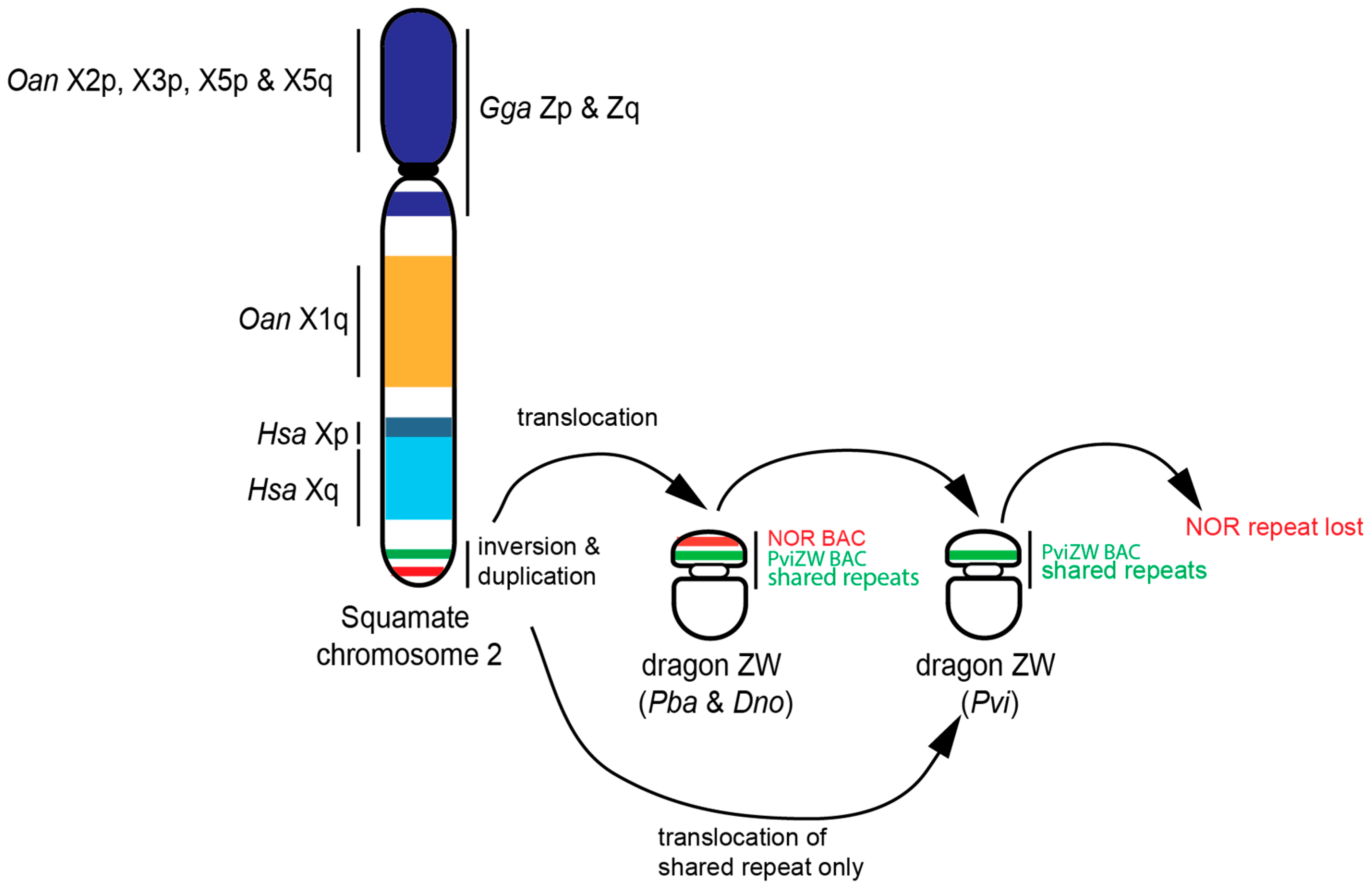
3.3. Blast Analysis of PviZW BAC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Meally, D.; Ezaz, T.; Georges, A.; Sarre, S.D.; Graves, J.A.M. Are some chromosomes particularly good at sex? Insights from amniotes. Chromosome Res. 2012, 20, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Haldane, J.B.S. Sex ratio and unisexual sterility in animal hybrids. J. Genet. 1922, 12, 101–109. [Google Scholar] [CrossRef]
- Dobzhansky, T.H. Studies in hybrid sterility. II. Localization of sterility factors in Drosophila pseudoobscura hybrids. Genetics 2017, 21, 113–135. [Google Scholar]
- Coyne, J.A. Genetic basis of male sterility in hybrids between two closely related species of Drosophila. Proc. Natl. Acad. Sci. USA 1984, 51, 4444–4447. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.A. “Two Rules of Speciation” revisited. Mol. Ecol. 2018, 27, 3749–3752. [Google Scholar] [CrossRef]
- Filatov, D.A. The two ‘rules of speciation’ in species with young sex chromosomes. Mol. Ecol. 2018, 27, 3799–3810. [Google Scholar] [CrossRef]
- Irwin, D.E. Sex chromosomes and speciation in birds and other ZW systems. Mol. Ecol. 2018, 27, 3831–3851. [Google Scholar] [CrossRef]
- O’Neill, M.J.; O’Neill, R.J. Sex chromosome repeats tip the balance towards speciation. Mol. Ecol. 2018, 27, 3745–3963. [Google Scholar] [CrossRef]
- Ezaz, T.; Moritz, B.; Waters, P.; Graves, J.A.M.; Georges, A.; Sarre, S.D. The ZW sex microchromosomes of an Australian dragon lizard share no homology with those of other reptiles or birds. Chromosome Res. 2009, 17, 965–973. [Google Scholar] [CrossRef]
- Ezaz, T.; Quinn, A.E.; Sarre, S.D.; Georges, A.; Graves, J.A.M. Molecular marker suggests rapid changes of sex-determining mechanisms in Australian dragon lizards. Chromosome Res. 2009, 17, 91–98. [Google Scholar] [CrossRef]
- Ezaz, T.; Sarre, S.D.; O’Meally, D.; Graves, J.A.M.; Georges, A. Sex chromosome evolution in lizards: Independent origins and rapid transitions. Cytogenet. Genome Res. 2009, 127, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Srikulnath, K.; Graves, J.A.M. Origin of amniote sex chromosomes: An ancestral super-sex chromosome, or common requirements? J. Hered. 2017, 108, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Quinn, A.E.; Miura, I.; Sarre, S.D.; Georges, A.; Graves, J.A.M. The dragon lizard Pogona vitticeps has ZZ/ZW micro-sex chromosomes. Chromosome Res. 2005, 13, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Quinn, A.E.; Georges, A.; Sarre, S.D.; Fiorenzo, G.; Ezaz, T.; Graves, J.A.M. Temperature sex reversal implies sex gene dosage in a reptile. Science 2007, 316, 411. [Google Scholar] [CrossRef] [PubMed]
- Quinn, A.E.; Ezaz, T.; Sarre, S.D.; Graves, J.A.M.; Georges, A. Extension, single-locus conversion and physical mapping of sex chromosome sequences identify the Z microchromosome and pseudo-autosomal region in a dragon lizard, Pogona vitticeps. Heredity 2010, 104, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Young, M.J.; O’Meally, D.; Sarre, S.D.; Georges, A.; Ezaz, T. Molecular cytogenetic map of the central bearded dragon, Pogona vitticeps (squamata: Agamidae). Chromosome Res. 2013, 21, 361–374. [Google Scholar] [CrossRef]
- Holleley, C.E.; O’Meally, D.; Sarre, S.D.; Graves, J.A.M.; Ezaz, T.; Matsubara, K.; Azad, B.; Zhang, X.; Georges, A. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 2015, 523, 79. [Google Scholar] [CrossRef]
- Quinn, A.E.; Sarre, S.D.; Ezaz, T.; Graves, J.A.M.; Georges, A. Evolutionary transitions between mechanisms of sex determination in vertebrates. Biol. Lett. 2011, 7, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Deveson, I.W.; Holleley, C.E.; Blackburn, J.; Graves, J.A.M.; Mattick, J.S.; Waters, P.D.; Georges, A. Differential intron retention in Jumonji chromatin modifier genes is implicated in reptile temperature-dependent sex determination. Sci. Adv. 2017, 3, e1700731. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Azad, B.; O’Meally, D.; Young, M.J.; Matsubara, K.; Edwards, M.J.; Zhang, X.; Holleley, C.E.; Deakin, J.E.; Graves, J.A.M.; et al. Sequence and gene content of a large fragment of a lizard sex chromosome and evaluation of candidate sex differentiating gene r-spondin 1. BMC Genom. 2013, 14, 899. [Google Scholar] [CrossRef]
- Deakin, J.E.; Edwards, M.J.; Patel, H.; O’Meally, D.; Lian, J.; Stenhouse, R.; Ryan, S.; Livernois, A.M.; Azad, B.; Holleley, C.E.; et al. Anchoring genome sequence to chromosomes of the central bearded dragon (Pogona vitticeps) enables reconstruction of ancestral squamate macrochromosomes and identifies sequence content of the Z chromosome. BMC Genom. 2016, 17, 447. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Azad, B.; Singchat, W.; Ezaz, T. Distribution and amplification of interstitial telomeric sequences (ITSs) in Australian dragon lizards support frequent chromosome fusions in Iguania. PLoS ONE 2019, 14, e0212683. [Google Scholar] [CrossRef] [PubMed]
- Abramyan, J.; Ezaz, T.; Graves, J.A.M.; Koopman, P. Z and W sex chromosomes in the cane toad (Bufo marinus). Chromosome Res. 2009, 17, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- De Smet, W.H.O. Description of the orcein stained karyotypes of 27 lizard species (Lacertilia Reptilia) belonging to the families Iguanidae, Agamidae, Chamaeleontidae and Gekkonidae (Ascalabota). Acta Zool. Pathol. Antverp. 1981, 76, 35–72. [Google Scholar]
- Witten, J.G. Some karyotypes of Australian agamids (Reptilia: Lacertilia). Aust. J. Zool. 1983, 3, 533–540. [Google Scholar] [CrossRef]
- Harlow, P.S. A harmless technique for sexing hatchling lizards. Herpetol. Rev. 1996, 27, 71–72. [Google Scholar]
- O’Meally, D.; Miller, H.; Patel, H.R.; Graves, J.A.M.; Ezaz, T. The first cytogenetic map of the tuatara, Sphenodon punctatus. Cytogenet. Genome Res. 2009, 127, 213–223. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hugall, A.F.; Foster, R.; Hutchinson, M.; Lee, M.S.Y. Phylogeny of Australasian agamid lizards based on nuclear and mitochondrial genes: Implications for morphological evolution and biogeography. Biol. J. Linn. Soc. 2008, 93, 343–358. [Google Scholar] [CrossRef]
- Singchat, W.; O’Connor, B.; Tawichasri, T.; Suntropong, A.; Silaparayoon, S.; Sutrarachun, S.; Maangmai, N.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; et al. Chromosome map of the Siamese cobra: Did partial synteny of sex chromosomes in the amniote represent “a hypothetical ancestral super-sex chromosome” or random distribution? BMC Genom. 2018, 19, 939. [Google Scholar] [CrossRef]
- O’Meally, D.; Patel, H.R.; Stiglec, R.; Sarre, S.D.; Georges, A.; Graves, J.A.M.; Ezaz, T. Non-homologous sex chromosomes of birds and snakes share repetitive sequences. Chromosome Res. 2010, 18, 787–800. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.; Hamilton, M.J.; Sites, J.W.; Baker, R.J. Location of ribosomal DNA in chromosomes of Squamate reptiles: Systematic and evolutionary implications. Herpetologica 1991, 47, 271–280. [Google Scholar]
- Kasahara, S.; Yonenaga-Yassuda, Y.; Rodrigues, M.T. Karyotype and evolution of the Tropidurus nanuzae species group (Sauria, Iguanidae). Rev. Bras. Genet. 1987, 2, 185–197. [Google Scholar]
- Srikulnath, K.; Matsubara, K.; Uno, Y.; Hongpan, A.; Suputtitada, S.; Apisitwanich, S.; Matsuda, Y.; Nishida, C. Karyological characterization of the butterfly lizard (Leiolepis reevesii rubritaeniata, Agamidae, Squamata) by molecular cytogenetic approach. Cytogenet. Genome Res. 2009, 125, 213–223. [Google Scholar] [CrossRef]
- Srikulnath, K.; Uno, Y.; Matsubara, K.; Thongpan, A.; Suputtitada, S.; Apisitwanich, S.; Nishida, C.; Matsuda, Y. Chromosomal localization of the 18S–28S and 5S rRNA genes and (TTAGGG)n sequences of butterfly lizards (Leiolepis belliana belliana and Leiolepis boehmei, Agamidae, Squamata). Genet. Mol. Biol. 2011, 34, 582–586. [Google Scholar] [CrossRef]
- Charlesworth, B.; Langley, C.H.; Stephan, W. The evolution of restricted recombination and the accumulation of repeated DNA sequences. Genetics 1986, 112, 947–962. [Google Scholar]
- Watson, J.M.; Meyne, J.; Graves, J.A.M. Ordered tandem arrangement of chromosomes in the sperm heads of monotreme mammals. Proc. Natl. Acad. Sci. USA 1996, 93, 10200–10205. [Google Scholar] [CrossRef]
- Kawagoshi, T.; Uno, Y.; Matsubara, K.; Matsuda, Y.; Nishida, C. The ZW micro-sex chromosomes of the Chinese soft-shelled turtle (Pelodiscus sinensis, Trionychidae, Testudines) have the same origin as chicken chromosome 15. Cytogenet. Genome Res. 2009, 125, 125–131. [Google Scholar] [CrossRef]
- Graves, J.; Peichel, C. Are homologies in vertebrate sex determination due to shared ancestry or to limited options? Genome Biol. 2010, 11, 205. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Diploidy (2n), SD and SC | Locality and Origin | Number of Animals Used (F + M) | |
|---|---|---|---|---|
| Pv151_P16 | AGI 329_J14 | |||
| Diporiphora nobbi | 32, GSD (ZW) | Vic, Australia | 1 + 1 | 1 + 1 |
| Pogona vitticeps | 32, GSD (ZW) | NSW, Australia | 1 + 1 | 1 + 1 |
| Pogona barbata | 32, GSD (ZW) | ACT, Australia | 1 + 1 | 1 + 1 |
| Amphibolurus norrisi | 32, GSD | Vic, Australia | 1 + 1 | 1 + 0 |
| Amphibolurus muricatus | 32, TSD | ACT, Australia | 1 + 1 | 1 + 0 |
| Ctenophorus pictus | 32, GSD | NSW, Australia | 1 + 1 | 1 + 1 |
| Physignathus lesueurii | 36, TSD | ACT, Australia | 1 + 1 | 1 + 1 |
| Physignathus cocincinus | 36, NK | Pet trade, Asia | 1 + 1 | 1 + 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsubara, K.; O’Meally, D.; Sarre, S.D.; Georges, A.; Srikulnath, K.; Ezaz, T. ZW Sex Chromosomes in Australian Dragon Lizards (Agamidae) Originated from a Combination of Duplication and Translocation in the Nucleolar Organising Region. Genes 2019, 10, 861. https://doi.org/10.3390/genes10110861
Matsubara K, O’Meally D, Sarre SD, Georges A, Srikulnath K, Ezaz T. ZW Sex Chromosomes in Australian Dragon Lizards (Agamidae) Originated from a Combination of Duplication and Translocation in the Nucleolar Organising Region. Genes. 2019; 10(11):861. https://doi.org/10.3390/genes10110861
Chicago/Turabian StyleMatsubara, Kazumi, Denis O’Meally, Stephen D. Sarre, Arthur Georges, Kornsorn Srikulnath, and Tariq Ezaz. 2019. "ZW Sex Chromosomes in Australian Dragon Lizards (Agamidae) Originated from a Combination of Duplication and Translocation in the Nucleolar Organising Region" Genes 10, no. 11: 861. https://doi.org/10.3390/genes10110861
APA StyleMatsubara, K., O’Meally, D., Sarre, S. D., Georges, A., Srikulnath, K., & Ezaz, T. (2019). ZW Sex Chromosomes in Australian Dragon Lizards (Agamidae) Originated from a Combination of Duplication and Translocation in the Nucleolar Organising Region. Genes, 10(11), 861. https://doi.org/10.3390/genes10110861