FAM64A: A Novel Oncogenic Target of Lung Adenocarcinoma Regulated by Both Strands of miR-99a (miR-99a-5p and miR-99a-3p)

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
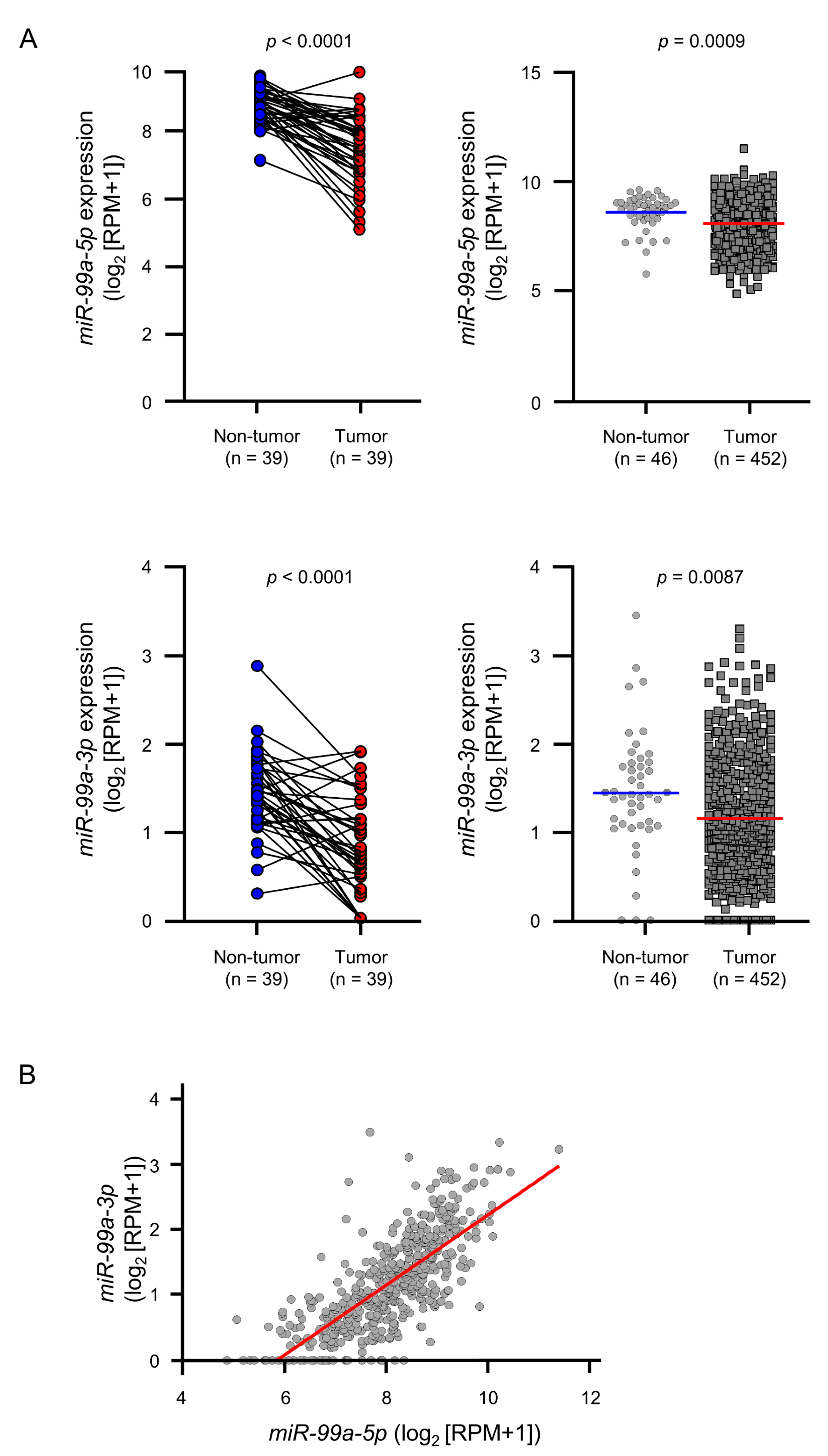
2.1. Data Mining of miRNA Target Genes and Their Expression in LUAD Clinical Specimens
2.2. Transfection of miRNAs, siRNAs and Plasmid Vectors into LUAD Cells and Functional Assays
2.3. Plasmid Construction and Dual-Luciferase Reporter Assays
2.4. Immunohistochemistry
2.5. Stastistical Analyses
3. Results
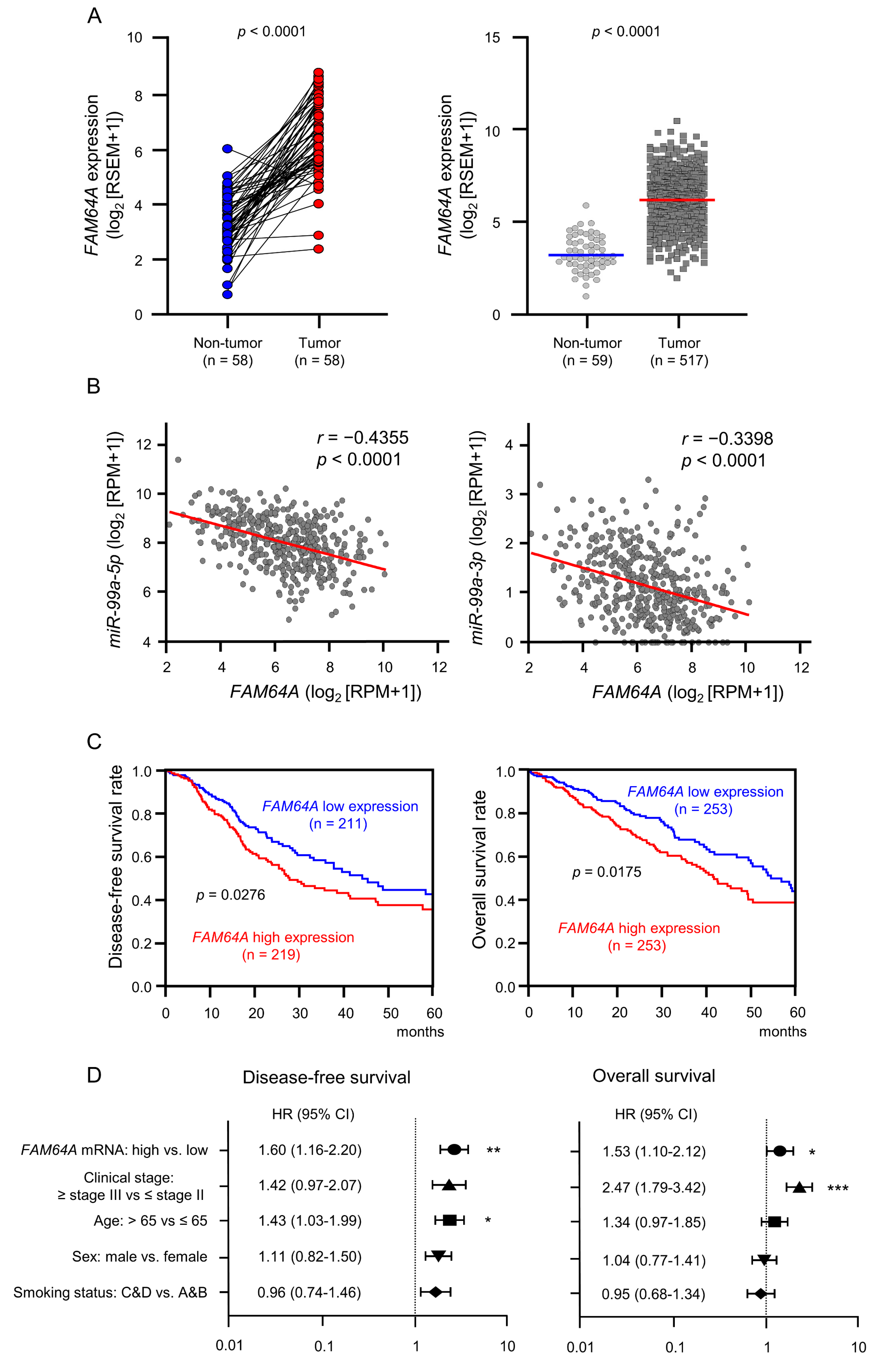
3.1. Downregulation of miR-99a-5p and miR-99a-3p in LUAD Clinical Specimens and Their Clinical Significance
3.2. Tumor-Suppressive Functions of miR-99a-5p and miR-99a-3p Assessed by Ectopic Expression Assays
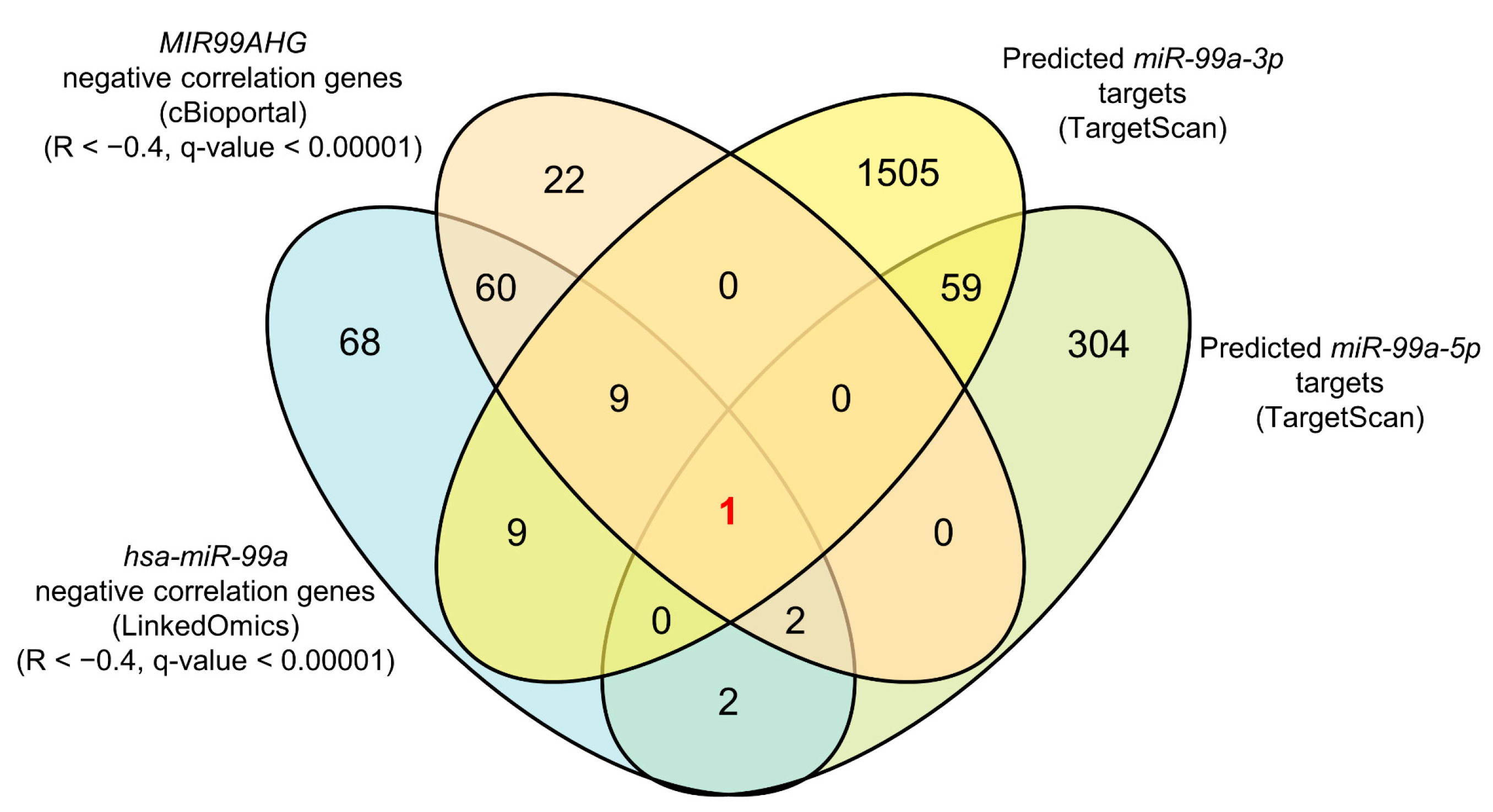
3.3. Identification of miR-99a-5p and miR-99a-3p Target Genes in LUAD
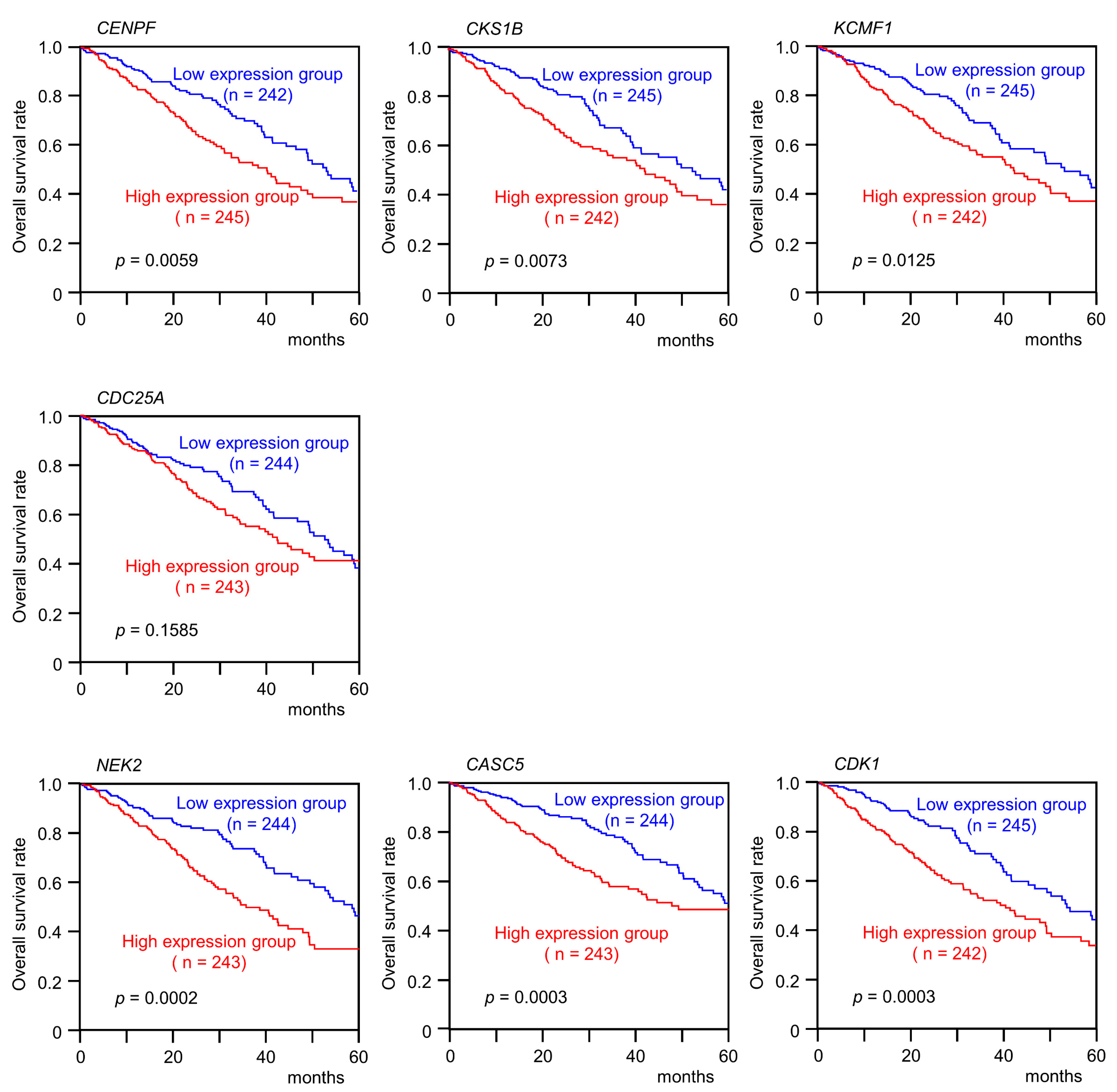
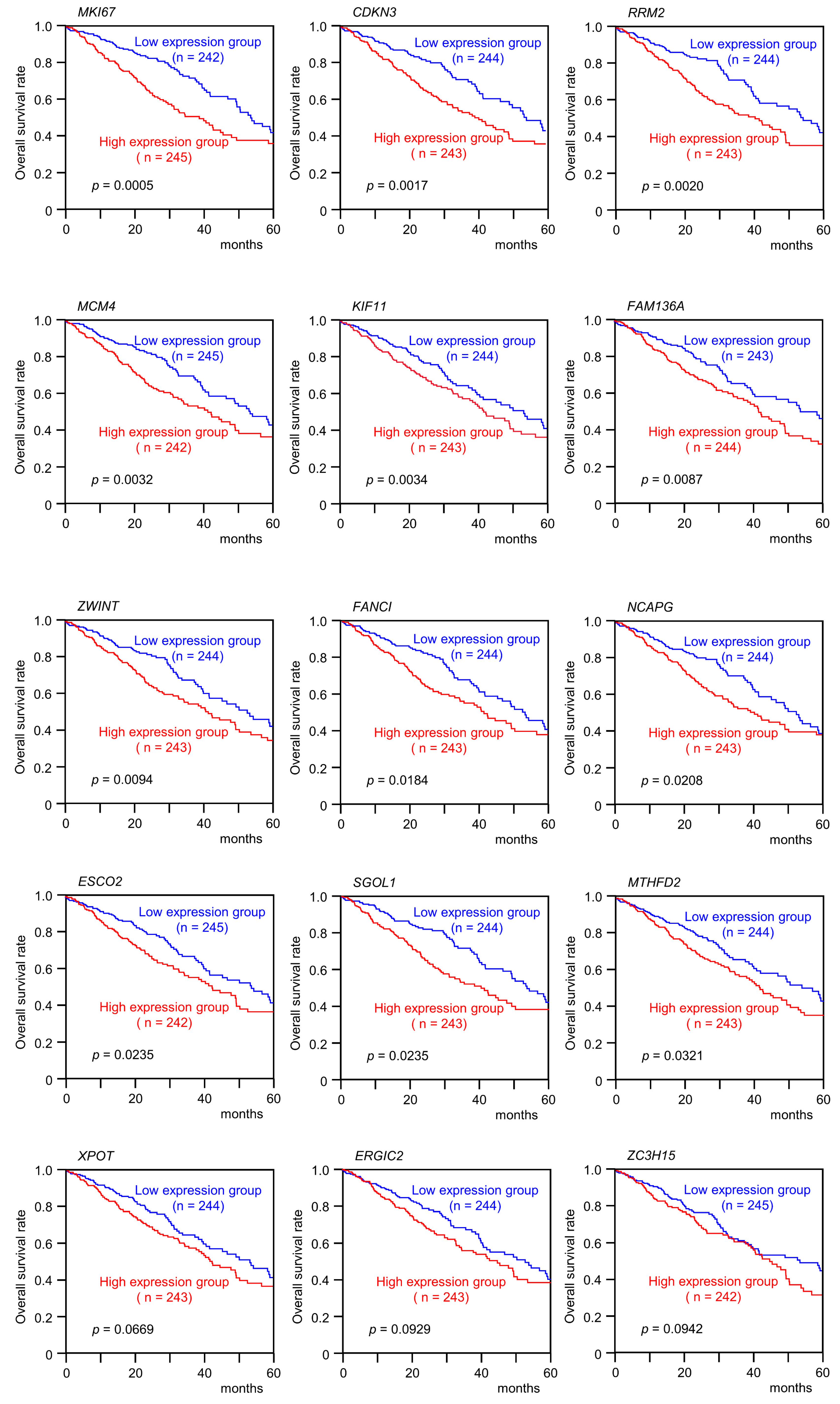
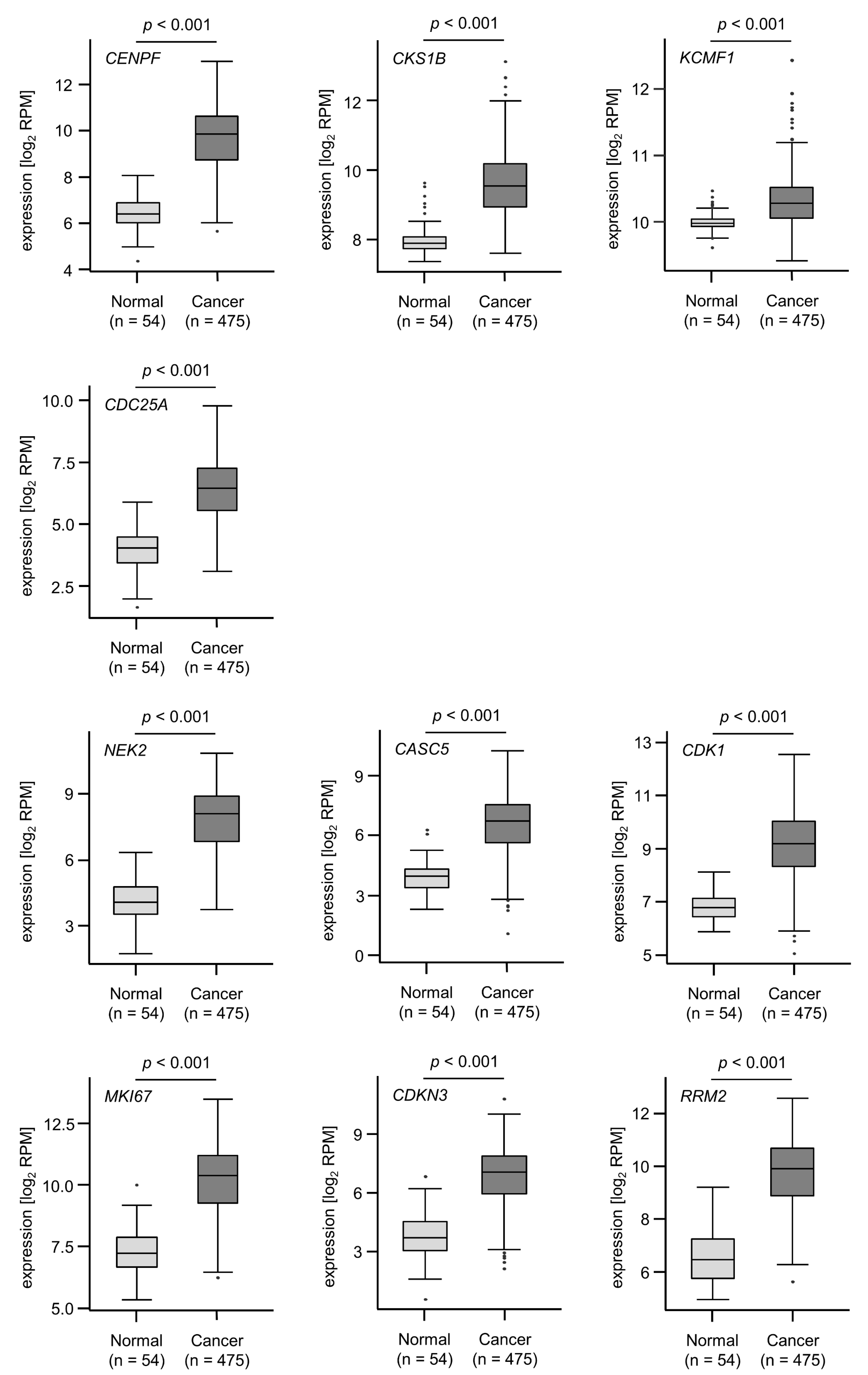
3.4. Clinical Significance of miR-99a-5p and miR-99a-3p Target Genes in LUAD Pathogenesis
3.5. Clinical Significance of FAM64A in LUAD Pathogenesis
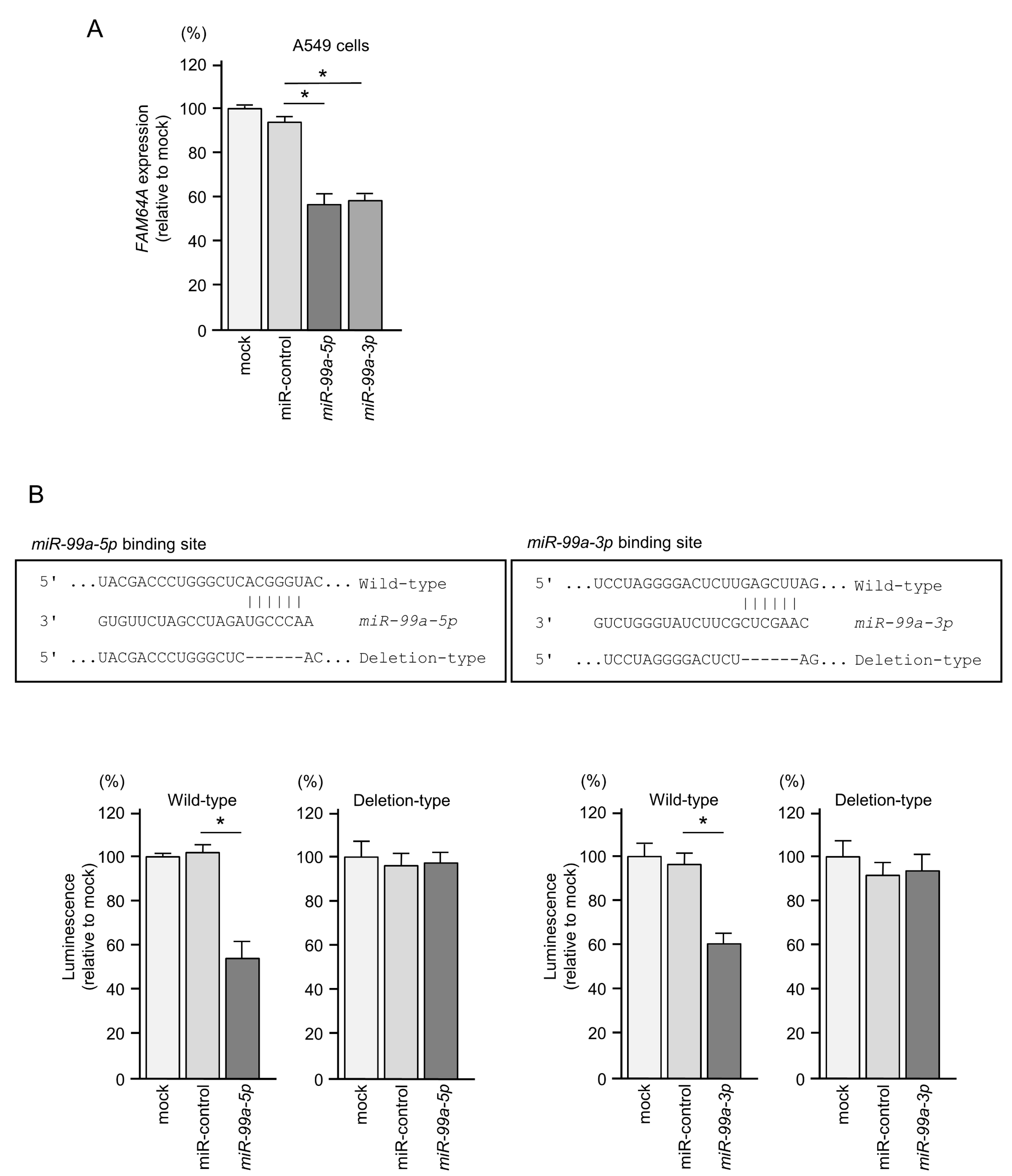
3.6. Direct Regulation of FAM64A by miR-99a-5p and miR-99a-3p in LUAD Cells
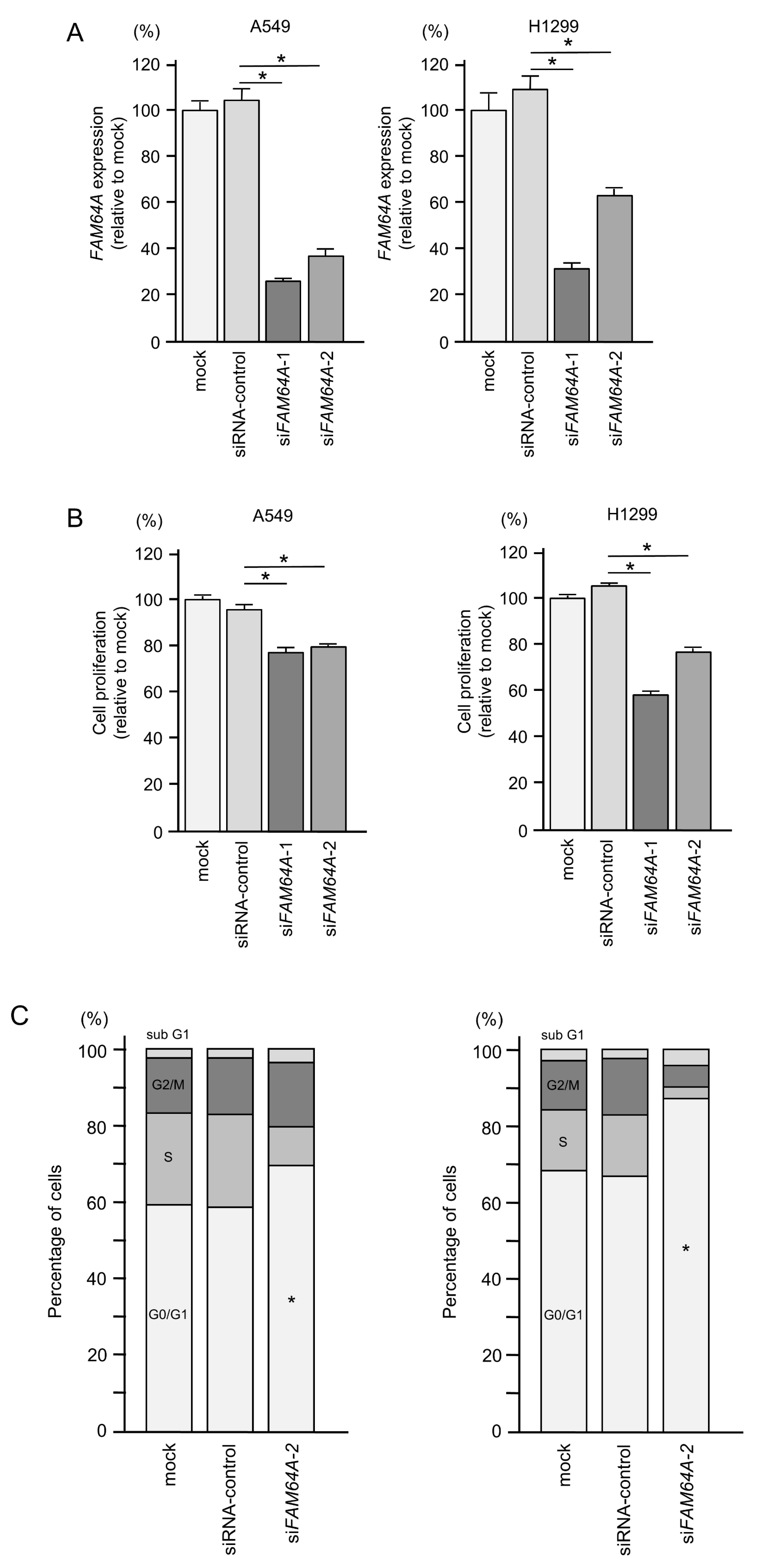
3.7. Effects of FAM64A Knockdown on Cell Proliferation and Cell Cycle in LUAD Cells
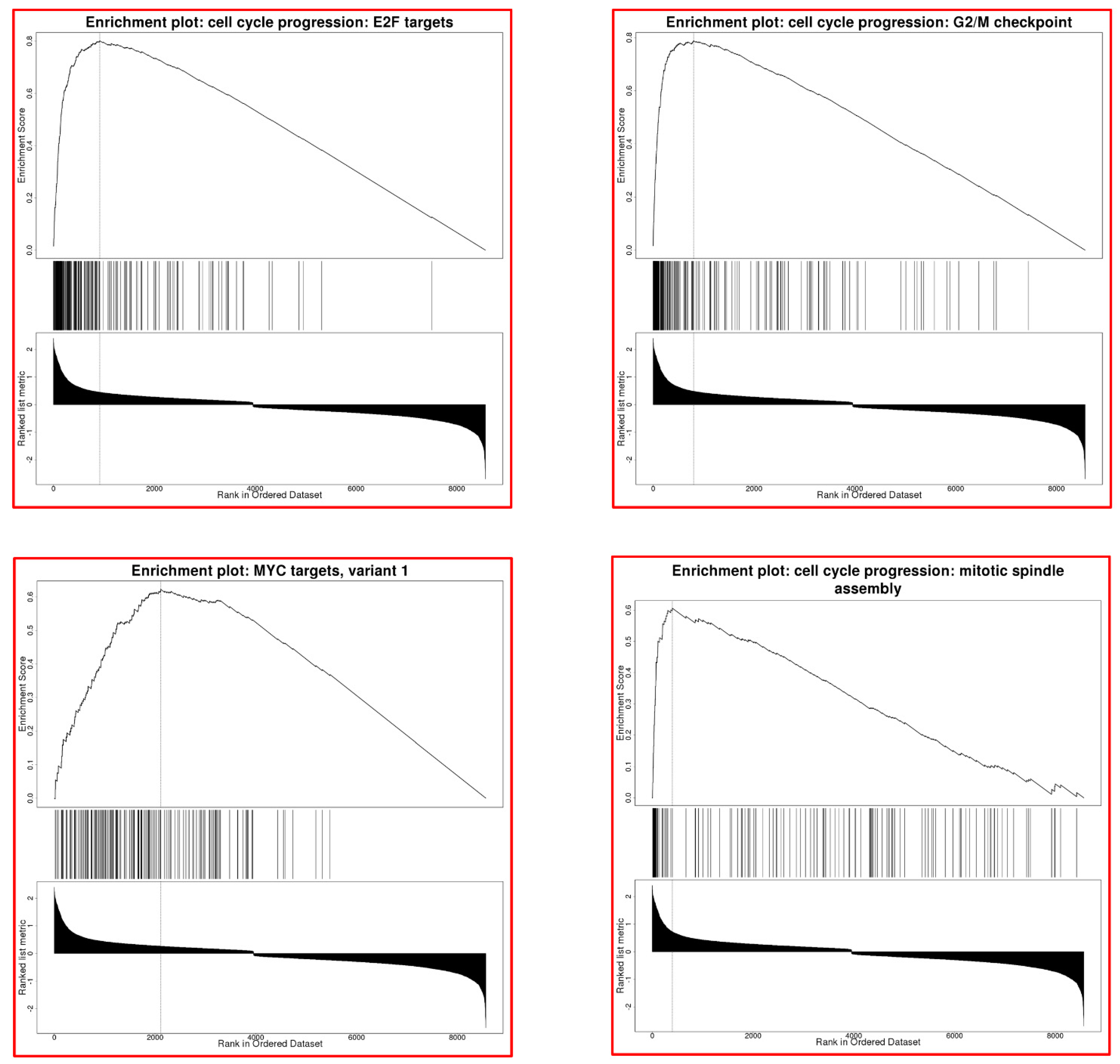
3.8. FAM64A Effects on Molecular Pathways in LUAD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Travis, W.D.; Brambilla, E.; Nicholson, A.G.; Yatabe, Y.; Austin, J.H.M.; Beasley, M.B.; Chirieac, L.R.; Dacic, S.; Duhig, E.; Flieder, D.B.; et al. The 2015 World Health Organization Classification of Lung Tumors: Impact of Genetic, Clinical and Radiologic Advances Since the 2004 Classification. J. Thorac. Oncol. 2015, 10, 1243–1260. [Google Scholar] [CrossRef]
- Goldstraw, P.; Chansky, K.; Crowley, J.; Rami-Porta, R.; Asamura, H.; Eberhardt, W.E.; Nicholson, A.G.; Groome, P.; Mitchell, A.; Bolejack, V.; et al. The IASLC Lung Cancer Staging Project: Proposals for Revision of the TNM Stage Groupings in the Forthcoming (Eighth) Edition of the TNM Classification for Lung Cancer. J. Thorac. Oncol. 2016, 11, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, L.; Rodríguez-Abreu, D.; Gadgeel, S.; Esteban, E.; Felip, E.; De Angelis, F.; Domine, M.; Clingan, P.; Hochmair, M.J.; Powell, S.F.; et al. Pembrolizumab plus Chemotherapy in Metastatic Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2018, 378, 2078–2092. [Google Scholar] [CrossRef] [PubMed]
- Hida, T.; Nokihara, H.; Kondo, M.; Kim, Y.H.; Azuma, K.; Seto, T.; Takiguchi, Y.; Nishio, M.; Yoshioka, H.; Imamura, F.; et al. Alectinib versus crizotinib in patients with ALK-positive non-small-cell lung cancer (J-ALEX): An open-label, randomised phase 3 trial. Lancet 2017, 390, 29–39. [Google Scholar] [CrossRef]
- Ramalingam, S.S.; Vansteenkiste, J.; Planchard, D.; Cho, B.C.; Gray, J.E.; Ohe, Y.; Zhou, C.; Reungwetwattana, T.; Cheng, Y.; Chewaskulyong, B.; et al. Overall Survival with Osimertinib in Untreated, EGFR-Mutated Advanced NSCLC. N. Engl. J. Med. 2020, 382, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Rosell, R.; Moran, T.; Queralt, C.; Porta, R.; Cardenal, F.; Camps, C.; Majem, M.; Lopez-Vivanco, G.; Isla, D.; Provencio, M.; et al. Screening for epidermal growth factor receptor mutations in lung cancer. N. Engl. J. Med. 2009, 361, 958–967. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Hobert, O. Gene regulation by transcription factors and microRNAs. Science 2008, 319, 1785–1786. [Google Scholar] [CrossRef]
- Iorio, M.V.; Croce, C.M. MicroRNAs in cancer: Small molecules with a huge impact. J. Clin. Oncol. 2009, 27, 5848–5856. [Google Scholar] [CrossRef] [PubMed]
- Baer, C.; Claus, R.; Plass, C. Genome-wide epigenetic regulation of miRNAs in cancer. Cancer Res. 2013, 73, 473–477. [Google Scholar] [CrossRef]
- Wu, S.G.; Chang, T.H.; Liu, Y.N.; Shih, J.Y. MicroRNA in Lung Cancer Metastasis. Cancers 2019, 11, 265. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.L.; Tsai, Y.M.; Lien, C.T.; Kuo, P.L.; Hung, A.J. The Roles of MicroRNA in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 1611. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Fukumoto, I.; Katada, K.; Okamoto, Y.; Seki, N. Deep sequencing-based microRNA expression signatures in head and neck squamous cell carcinoma: Dual strands of pre-miR-150 as antitumor miRNAs. Oncotarget 2017, 8, 30288–30304. [Google Scholar] [CrossRef] [PubMed]
- Yonemori, K.; Seki, N.; Idichi, T.; Kurahara, H.; Osako, Y.; Koshizuka, K.; Arai, T.; Okato, A.; Kita, Y.; Arigami, T.; et al. The microRNA expression signature of pancreatic ductal adenocarcinoma by RNA sequencing: Anti-tumour functions of the microRNA-216 cluster. Oncotarget 2017, 8, 70097–70115. [Google Scholar] [CrossRef]
- Toda, H.; Kurozumi, S.; Kijima, Y.; Idichi, T.; Shinden, Y.; Yamada, Y.; Arai, T.; Maemura, K.; Fujii, T.; Horiguchi, J.; et al. Molecular pathogenesis of triple-negative breast cancer based on microRNA expression signatures: Antitumor miR-204-5p targets AP1S3. J. Hum. Genet. 2018, 63, 1197–1210. [Google Scholar] [CrossRef]
- Toda, H.; Seki, N.; Kurozumi, S.; Shinden, Y.; Yamada, Y.; Nohata, N.; Moriya, S.; Idichi, T.; Maemura, K.; Fujii, T.; et al. RNA-sequence-based microRNA expression signature in breast cancer: Tumor-suppressive miR-101-5p regulates molecular pathogenesis. Mol. Oncol. 2020, 14, 426–446. [Google Scholar] [CrossRef]
- Wada, M.; Goto, Y.; Tanaka, T.; Okada, R.; Moriya, S.; Idichi, T.; Noda, M.; Sasaki, K.; Kita, Y.; Kurahara, H.; et al. RNA sequencing-based microRNA expression signature in esophageal squamous cell carcinoma: Oncogenic targets by antitumor miR-143-5p and miR-143-3p regulation. J. Hum. Genet. 2020. [Google Scholar] [CrossRef]
- Mataki, H.; Seki, N.; Chiyomaru, T.; Enokida, H.; Goto, Y.; Kumamoto, T.; Machida, K.; Mizuno, K.; Nakagawa, M.; Inoue, H. Tumor-suppressive microRNA-206 as a dual inhibitor of MET and EGFR oncogenic signaling in lung squamous cell carcinoma. Int. J. Oncol. 2015, 46, 1039–1050. [Google Scholar] [CrossRef]
- Uchida, A.; Seki, N.; Mizuno, K.; Misono, S.; Yamada, Y.; Kikkawa, N.; Sanada, H.; Kumamoto, T.; Suetsugu, T.; Inoue, H. Involvement of dual-strand of the miR-144 duplex and their targets in the pathogenesis of lung squamous cell carcinoma. Cancer Sci. 2019, 110, 420–432. [Google Scholar] [CrossRef]
- Misono, S.; Seki, N.; Mizuno, K.; Yamada, Y.; Uchida, A.; Arai, T.; Kumamoto, T.; Sanada, H.; Suetsugu, T.; Inoue, H. Dual strands of the miR-145 duplex (miR-145-5p and miR-145-3p) regulate oncogenes in lung adenocarcinoma pathogenesis. J. Hum. Genet. 2018, 63, 1015–1028. [Google Scholar] [CrossRef] [PubMed]
- Sanada, H.; Seki, N.; Mizuno, K.; Misono, S.; Uchida, A.; Yamada, Y.; Moriya, S.; Kikkawa, N.; Machida, K.; Kumamoto, T.; et al. Involvement of Dual Strands of miR-143 (miR-143-5p and miR-143-3p) and Their Target Oncogenes in the Molecular Pathogenesis of Lung Adenocarcinoma. Int. J. Mol. Sci. 2019, 20, 4482. [Google Scholar] [CrossRef] [PubMed]
- Misono, S.; Seki, N.; Mizuno, K.; Yamada, Y.; Uchida, A.; Sanada, H.; Moriya, S.; Kikkawa, N.; Kumamoto, T.; Suetsugu, T.; et al. Molecular Pathogenesis of Gene Regulation by the miR-150 Duplex: miR-150-3p Regulates TNS4 in Lung Adenocarcinoma. Cancers 2019, 11, 601. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA. 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2017, 46, D956–D963. [Google Scholar] [CrossRef] [PubMed]
- Rotow, J.; Bivona, T.G. Understanding and targeting resistance mechanisms in NSCLC. Nat. Rev. Cancer 2017, 17, 637–658. [Google Scholar] [CrossRef] [PubMed]
- Quintanal-Villalonga, Á.; Chan, J.M.; Yu, H.A.; Pe’er, D.; Sawyers, C.L.; Sen, T.; Rudin, C.M. Lineage plasticity in cancer: A shared pathway of therapeutic resistance. Nat. Rev. Clin. Oncol. 2020, 17, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Adams, C.M.; Jiang, W.; Greenawalt, E.; Eischen, C.M. Pan-cancer analysis reveals cooperativity of both strands of microRNA that regulate tumorigenesis and patient survival. Nat. Commun. 2020, 11, 968. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.H.; Zhang, C.L.; Dong, F.S.; Zhang, Y.M. miR-99a suppresses the metastasis of human non-small cell lung cancer cells by targeting AKT1 signaling pathway. J. Cell. Biochem. 2015, 116, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Ma, J.; Chen, L.; Piao, S.; Zhang, Y.; Zhang, S.; Ma, H.; Li, Y.; Qu, Y.; Wang, X.; et al. MiR-99a Enhances the Radiation Sensitivity of Non-Small Cell Lung Cancer by Targeting mTOR. Cell. Physiol. Biochem. 2018, 46, 471–481. [Google Scholar] [CrossRef]
- Arai, T.; Okato, A.; Yamada, Y.; Sugawara, S.; Kurozumi, A.; Kojima, S.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Regulation of NCAPG by miR-99a-3p (passenger strand) inhibits cancer cell aggressiveness and is involved in CRPC. Cancer Med. 2018, 7, 1988–2002. [Google Scholar] [CrossRef]
- Okada, R.; Koshizuka, K.; Yamada, Y.; Moriya, S.; Kikkawa, N.; Kinoshita, T.; Hanazawa, T.; Seki, N. Regulation of Oncogenic Targets by miR-99a-3p (Passenger Strand of miR-99a-Duplex) in Head and Neck Squamous Cell Carcinoma. Cells 2019, 8, 1535. [Google Scholar] [CrossRef]
- Huszar, D.; Theoclitou, M.E.; Skolnik, J.; Herbst, R. Kinesin motor proteins as targets for cancer therapy. Cancer Metastasis Rev. 2009, 28, 197–208. [Google Scholar] [CrossRef]
- Rath, O.; Kozielski, F. Kinesins and cancer. Nat. Rev. Cancer 2012, 12, 527–539. [Google Scholar] [CrossRef]
- Archangelo, L.F.; Gläsner, J.; Krause, A.; Bohlander, S.K. The novel CALM interactor CATS influences the subcellular localization of the leukemogenic fusion protein CALM/AF10. Oncogene 2006, 25, 4099–4109. [Google Scholar] [CrossRef]
- Archangelo, L.F.; Greif, P.A.; Hölzel, M.; Harasim, T.; Kremmer, E.; Przemeck, G.K.; Eick, D.; Deshpande, A.J.; Buske, C.; de Angelis, M.H.; et al. The CALM and CALM/AF10 interactor CATS is a marker for proliferation. Mol. Oncol. 2008, 2, 356–367. [Google Scholar] [CrossRef]
- Caudell, D.; Aplan, P.D. The role of CALM-AF10 gene fusion in acute leukemia. Leukemia 2008, 22, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Barbutti, I.; Xavier-Ferrucio, J.M.; Machado-Neto, J.A.; Ricon, L.; Traina, F.; Bohlander, S.K.; Saad, S.T.; Archangelo, L.F. CATS (FAM64A) abnormal expression reduces clonogenicity of hematopoietic cells. Oncotarget 2016, 7, 68385–68396. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Kodama, A.; Honda, T.; Hanashima, A.; Ujihara, Y.; Murayama, T.; Nishimatsu, S.I.; Mohri, S. Fam64a is a novel cell cycle promoter of hypoxic fetal cardiomyocytes in mice. Sci. Rep. 2017, 7, 4486. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Zheng, X.; Lu, S.; He, Z.; Miao, Y.; Huang, H.; Chu, X.; Cai, C.; Zou, F. Knockdown of FAM64A suppresses proliferation and migration of breast cancer cells. Breast Cancer 2019, 26, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yuan, H.; Li, Z.; Zhang, J.; Wu, J.; Chen, Y.; Shi, Q.; Ren, W.; Shao, N.; Ying, X. Transcriptional response profiles of paired tumor-normal samples offer novel perspectives in pan-cancer analysis. Oncotarget 2017, 8, 41334–41347. [Google Scholar] [CrossRef]
- Jiang, L.; Ren, L.; Zhang, X.; Chen, H.; Chen, X.; Lin, C.; Wang, L.; Hou, N.; Pan, J.; Zhou, Z.; et al. Overexpression of PIMREG promotes breast cancer aggressiveness via constitutive activation of NF-κB signaling. EBioMedicine 2019, 43, 188–200. [Google Scholar] [CrossRef]
- Xu, Z.S.; Zhang, H.X.; Li, W.W.; Ran, Y.; Liu, T.T.; Xiong, M.G.; Li, Q.L.; Wang, S.Y.; Wu, M.; Shu, H.B.; et al. FAM64A positively regulates STAT3 activity to promote Th17 differentiation and colitis-associated carcinogenesis. Proc. Natl. Acad. Sci. USA 2019, 116, 10447–10452. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entrez Gene ID | Gene Symbol | Gene Name | Location | Total Sites | 5-Year OS p-Value |
|---|---|---|---|---|---|
| Putative targets by miR-99a-5p regulation in LUAD cells | |||||
| 1063 | CENPF | Centromere protein F | 1q41 | 1 | 0.0059 |
| 1163 | CKS1B | CDC28 protein kinase regulatory subunit 1B | 1q21.3 | 1 | 0.0073 |
| 56888 | KCMF1 | Potassium channel modulatory factor 1 | 2p11.2 | 1 | 0.0125 |
| 417685 | FAM64A | Family with sequence similarity 64 member A | 17p13.2 | 1 | 0.0176 |
| 993 | CDC25A | Cell division cycle 25A | 3p21.31 | 1 | 0.1585 |
| Putative targets by miR-99a-3p regulation in LUAD cells | |||||
| 4751 | NEK2 | NIMA related kinase 2 | 1q32.3 | 1 | 0.0002 |
| 57082 | CASC5 | Cancer susceptibility candidate 5 | 15q15.1 | 1 | 0.0003 |
| 983 | CDK1 | Cyclin dependent kinase 1 | 10q21.2 | 1 | 0.0003 |
| 4288 | MKI67 | Marker of proliferation Ki-67 | 10q26.2 | 1 | 0.0005 |
| 1033 | CDKN3 | Cyclin dependent kinase inhibitor 3 | 14q22.2 | 1 | 0.0017 |
| 6241 | RRM2 | Ribonucleotide reductase regulatory subunit M2 | 2p25.1 | 1 | 0.002 |
| 4173 | MCM4 | Minichromosome maintenance complex component 4 | 8q11.21 | 1 | 0.0032 |
| 3832 | KIF11 | Kinesin family member 11 | 10q23.33 | 1 | 0.0034 |
| 84908 | FAM136A | Family with sequence similarity 136 member A | 2p13.3 | 1 | 0.0087 |
| 11130 | ZWINT | ZW10 interacting kinetochore protein | 10q21.1 | 1 | 0.0094 |
| 55215 | FANCI | FA complementation group I | 15q26.1 | 1 | 0.0108 |
| 64151 | NCAPG | Non-SMC condensin I complex subunit G | 4p15.31 | 1 | 0.0208 |
| 157570 | ESCO2 | Establishment of sister chromatid cohesion N-acetyltransferase 2 | 8p21.1 | 1 | 0.0235 |
| 151648 | SGOL1 | Shugoshin 1 | 3p24.3 | 1 | 0.0235 |
| 10797 | MTHFD2 | Methylenetetrahydrofolate dehydrogenase (NADP+ dependent) 2, methenyltetrahydrofolate cyclohydrolase | 2p13.1 | 1 | 0.0321 |
| 11260 | XPOT | Exportin for tRNA | 12q14.2 | 1 | 0.0669 |
| 417685 | FAM64A | Family with sequence similarity 64 member A | 17p13.2 | 1 | 0.0756 |
| 51290 | ERGIC2 | ERGIC and golgi 2 | 12p11.22 | 1 | 0.0929 |
| 55854 | ZC3H15 | Zinc finger CCCH-type containing 15 | 2q32.1 | 1 | 0.0942 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizuno, K.; Tanigawa, K.; Nohata, N.; Misono, S.; Okada, R.; Asai, S.; Moriya, S.; Suetsugu, T.; Inoue, H.; Seki, N. FAM64A: A Novel Oncogenic Target of Lung Adenocarcinoma Regulated by Both Strands of miR-99a (miR-99a-5p and miR-99a-3p). Cells 2020, 9, 2083. https://doi.org/10.3390/cells9092083
Mizuno K, Tanigawa K, Nohata N, Misono S, Okada R, Asai S, Moriya S, Suetsugu T, Inoue H, Seki N. FAM64A: A Novel Oncogenic Target of Lung Adenocarcinoma Regulated by Both Strands of miR-99a (miR-99a-5p and miR-99a-3p). Cells. 2020; 9(9):2083. https://doi.org/10.3390/cells9092083
Chicago/Turabian StyleMizuno, Keiko, Kengo Tanigawa, Nijiro Nohata, Shunsuke Misono, Reona Okada, Shunichi Asai, Shogo Moriya, Takayuki Suetsugu, Hiromasa Inoue, and Naohiko Seki. 2020. "FAM64A: A Novel Oncogenic Target of Lung Adenocarcinoma Regulated by Both Strands of miR-99a (miR-99a-5p and miR-99a-3p)" Cells 9, no. 9: 2083. https://doi.org/10.3390/cells9092083
APA StyleMizuno, K., Tanigawa, K., Nohata, N., Misono, S., Okada, R., Asai, S., Moriya, S., Suetsugu, T., Inoue, H., & Seki, N. (2020). FAM64A: A Novel Oncogenic Target of Lung Adenocarcinoma Regulated by Both Strands of miR-99a (miR-99a-5p and miR-99a-3p). Cells, 9(9), 2083. https://doi.org/10.3390/cells9092083