Experimental Models for Fungal Keratitis: An Overview of Principles and Protocols

Abstract
1. Introduction to Fungal Keratitis
1.1. Corneal Structure and Function
1.2. Fungal Keratitis Pathogenesis
2. Experimental Methods Available for FK Research
3. In Vivo Models
3.1. Mice
3.1.1. Mouse Strains
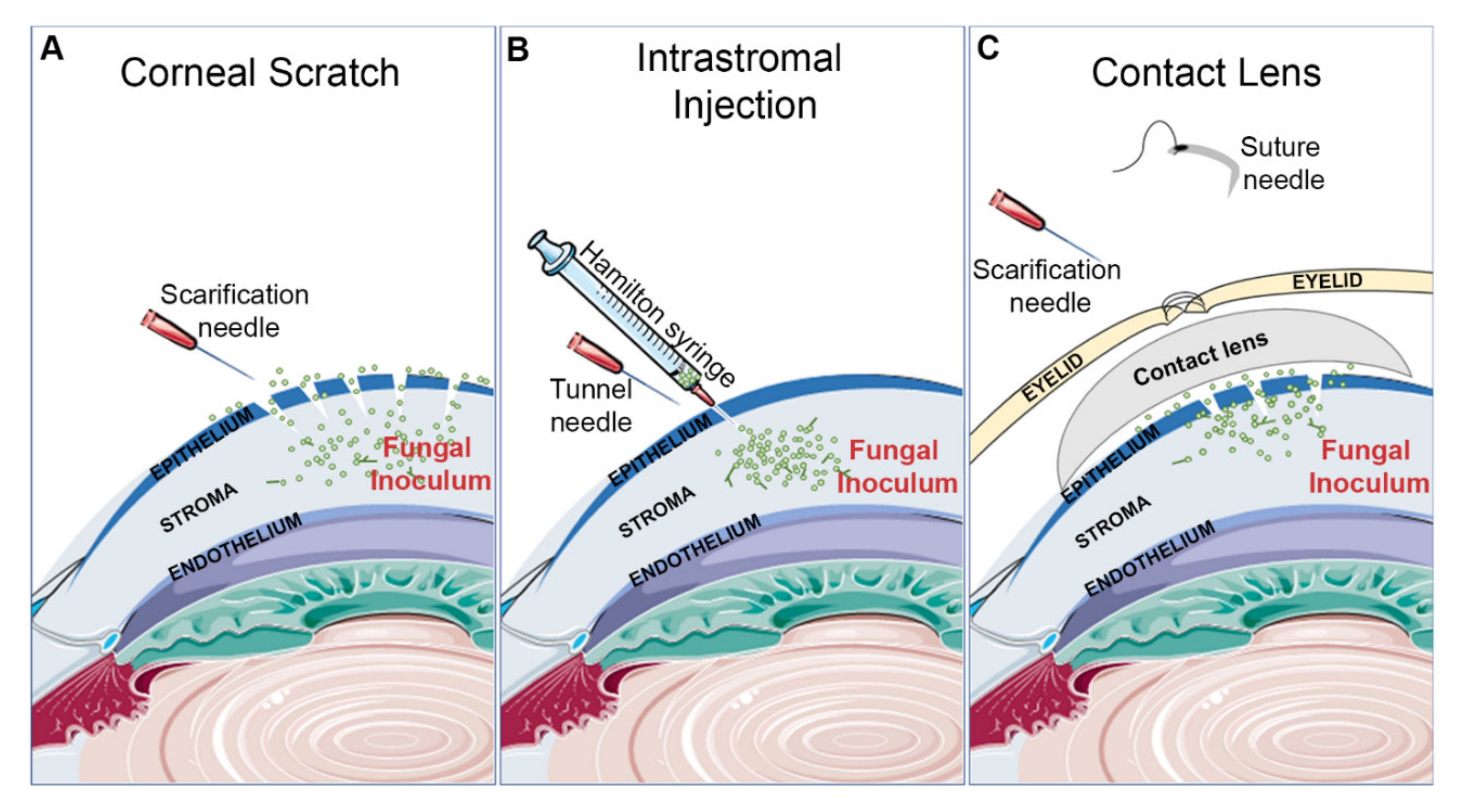
3.1.2. Inoculation Procedures
Corneal Scratch and Topical Inoculation
Intrastromal Injection
Contact Lens
3.1.3. Immunosuppression
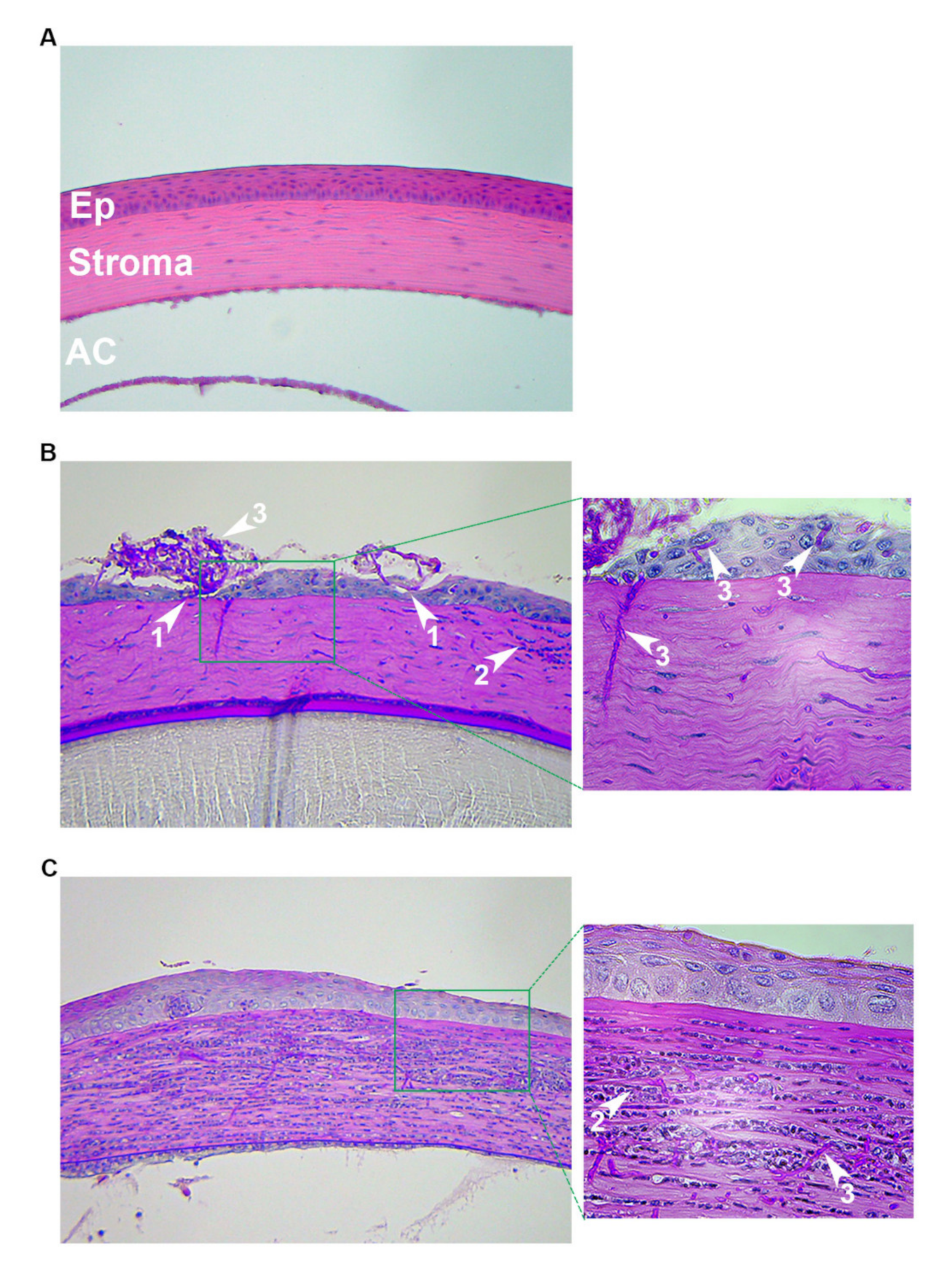
3.1.4. Disease Assessment
3.1.5. Antifungal Therapy
3.2. Rats
3.3. Rabbits
4. Ex Vivo Models
5. In Vitro Models
6. Conclusions
7. Experimental Methods Utilized in This Study
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thomas, P.A.; Kaliamurthy, J. Mycotic Keratitis: Epidemiology, Diagnosis and Management. Clin. Microbiol. Infect. 2013, 19, 210–220. [Google Scholar] [CrossRef]
- Saha, S.; Banerjee, D.; Khetan, A.; Sengupta, J. Epidemiological Profile of Fungal Keratitis in Urban Population of West Bengal, India. Oman J. Ophthalmol. 2009, 2, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.H.; Miller, D.; Alfonso, E.C. The Changing Spectrum of Fungal Keratitis in South Florida. Ophthalmology 1994, 101, 1005–1013. [Google Scholar] [CrossRef]
- Qiao, G.L.; Ling, J.; Wong, T.; Yeung, S.N.; Iovieno, A. Candida Keratitis: Epidemiology, Management, and Clinical Outcomes. Cornea 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.C.; Grant, G.B.; O’Donnell, K.; Wannemuehler, K.A.; Noble-Wang, J.; Rao, C.Y.; Jacobson, L.M.; Crowell, C.S.; Sneed, R.S.; Lewis, F.M.T.; et al. Multistate Outbreak of Fusarium Keratitis Associated with Use of a Contact Lens Solution. JAMA 2006, 296, 953–963. [Google Scholar] [CrossRef]
- Short, D.P.; O’Donnell, K.; Thrane, U.; Nielsen, K.F.; Zhang, N.; Juba, J.H.; Geiser, D.M. Phylogenetic relationships among members of the Fusarium solani species complex in human infections and the descriptions of F. keratoplasticum sp. nov. and F. petroliphilum stat. nov. Fungal Genet. Biol. 2013, 53, 59–70. [Google Scholar] [CrossRef]
- Spadea, L.; Maraone, G.; Verboschi, F.; Vingolo, E.M.; Tognetto, D. Effect of Corneal Light Scatter on Vision: A Review of the Literature. Int. J. Ophthalmol. 2016, 9, 459–464. [Google Scholar] [CrossRef]
- Meek, K.M.; Knupp, C. Corneal Structure and Transparency. Prog. Retin. Eye Res. 2015, 49, 1–16. [Google Scholar] [CrossRef]
- McDermott, A.M. Antimicrobial Compounds in Tears. Exp. Eye Res. 2013, 117, 53–61. [Google Scholar] [CrossRef]
- Han, J.; Jyoti, M.A.; Song, H.-Y.; Jang, W.S. Antifungal Activity and Action Mechanism of Histatin 5-Halocidin Hybrid Peptides against Candida Ssp. PLoS ONE 2016, 11, e0150196. [Google Scholar] [CrossRef]
- Brissette-Storkus, C.S.; Reynolds, S.M.; Lepisto, A.J.; Hendricks, R.L. Identification of a Novel Macrophage Population in the Normal Mouse Corneal Stroma. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2264–2271. [Google Scholar]
- Fukuda, K.; Ishida, W.; Fukushima, A.; Nishida, T. Corneal Fibroblasts as Sentinel Cells and Local Immune Modulators in Infectious Keratitis. Int. J. Mol. Sci. 2017, 18, 1831. [Google Scholar] [CrossRef] [PubMed]
- Hori, J. Mechanisms of Immune Privilege in the Anterior Segment of the Eye: What We Learn from Corneal Transplantation. J. Ocul. Biol. Dis. Inform. 2008, 1, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Hori, J.; Yamaguchi, T.; Keino, H.; Hamrah, P.; Maruyama, K. Immune Privilege in Corneal Transplantation. Prog. Retin. Eye Res. 2019, 72, 100758. [Google Scholar] [CrossRef]
- Bourne, W.M. Biology of the Corneal Endothelium in Health and Disease. Eye 2003, 17, 912–918. [Google Scholar] [CrossRef]
- Ansari, Z.; Miller, D.; Galor, A. Current Thoughts in Fungal Keratitis: Diagnosis and Treatment. Curr. Fungal Infect. Rep. 2013, 7, 209–218. [Google Scholar] [CrossRef]
- Ou, J.I.; Acharya, N.R. Epidemiology and Treatment of Fungal Corneal Ulcers. Int. Ophthalmol. Clin. 2007, 47, 7–16. [Google Scholar] [CrossRef]
- Klotz, S.A.; Penn, C.C.; Negvesky, G.J.; Butrus, S.I. Fungal and Parasitic Infections of the Eye. Clin. Microbiol. Rev. 2000, 13, 662–685. [Google Scholar] [CrossRef]
- Radfar, L.; Shea, Y.; Fischer, S.H.; Sankar, V.; Leakan, R.A.; Baum, B.J.; Pillemer, S.R. Fungal load and candidiasis in Sjogren’s syndrome. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2003, 96, 283–287. [Google Scholar] [CrossRef]
- Zhu, W.-S.; Wojdyla, K.; Donlon, K.; Thomas, P.A.; Eberle, H.I. Extracellular Proteases of Aspergillus flavus: Fungal Keratitis, Proteases, and Pathogenesis. Diagn. Microbiol. Infect. Dis. 1990, 13, 491–497. [Google Scholar] [CrossRef]
- Gopinathan, U.; Ramakrishna, T.; Willcox, M.; Rao, C.M.; Balasubramanian, D.; Kulkarni, A.; Vemuganti, G.K.; Rao, G.N. Enzymatic, Clinical and Histologic Evaluation of Corneal Tissues in Experimental Fungal Keratitis in Rabbits. Exp. Eye Res. 2001, 72, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, C.; Kuchler, K. Fungal Pathogens—A Sweet and Sour Treat for Toll-like Receptors. Front. Cell. Infect. Microbiol. 2012, 2, 142. [Google Scholar] [CrossRef]
- Kumar, A.; Yu, F.-S. Toll-Like Receptors and Corneal Innate Immunity. Curr. Mol. Med. 2006, 6, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Leal, S.M.; Cowden, S.; Hsia, Y.-C.; Ghannoum, M.A.; Momany, M.; Pearlman, E. Distinct Roles for Dectin-1 and TLR4 in the Pathogenesis of Aspergillus fumigatus Keratitis. PLoS Pathog. 2010, 6, e1000976. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.K.; Dohlman, T.H.; Dana, R. Corneal Lymphatics: Role in Ocular Inflammation as Inducer and Responder of Adaptive Immunity. J. Clin. Cell. Immunol. 2014, 5, 1000256. [Google Scholar] [CrossRef] [PubMed]
- Menda, S.A.; Das, M.; Panigrahi, A. Association of Postfungal Keratitis Corneal Scar Features with Visual Acuity. JAMA Ophthalmol. 2020, 138, 113–119. [Google Scholar] [CrossRef]
- Yuan, X.; Wilhelmus, K.R. Corneal neovascularization during experimental fungal keratitis. Mol. Vis. 2009, 15, 1988–1996. [Google Scholar]
- Hazlett, L.; Suvas, S.; McClellan, S.; Ekanayaka, S. Challenges of Corneal Infections. Expert Rev. Ophthalmol. 2016, 11, 285–297. [Google Scholar] [CrossRef]
- Henriksson, J.T.; McDermott, A.M.; Bergmanson, J.P.G. Dimensions and Morphology of the Cornea in Three Strains of Mice. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3648–3654. [Google Scholar] [CrossRef]
- Pal-Ghosh, S.; Tadvalkar, G.; Jurjus, R.A.; Zieske, J.D.; Stepp, M.A. BALB/c and C57BL6 Mouse Strains Vary in Their Ability to Heal Corneal Epithelial Debridement Wounds. Exp. Eye Res. 2008, 87, 478–486. [Google Scholar] [CrossRef]
- Li, C.-Y.; Li, C.; Li, H.; Zhao, G.-Q.; Lin, J.; Wang, Q.; Peng, X.-D.; Xu, Q.; Zhu, G.-Q.; Jiang, J.-Q. Disparate Expression of Autophagy in Corneas of C57BL/6 Mice and BALB/c Mice after Aspergillus fumigatus Infection. Int. J. Ophthalmol. 2019, 12, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Huang, W.; Deng, Q.; Wu, M.; Jiang, H.; Lin, X.; Sun, Y.; Huang, X.; Yuan, J. Inhibition of TREM-1 and Dectin-1 Alleviates the Severity of Fungal Keratitis by Modulating Innate Immune Responses. PLoS ONE 2016, 11, e0150114. [Google Scholar] [CrossRef]
- Wu, T.G.; Wilhelmus, K.R.; Mitchell, B.M. Experimental Keratomycosis in a Mouse Model. Investig. Ophthalmol. Vis. Sci. 2003, 44, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Yu, F.-S.X. Chitinase 3-Like 1 Promotes Candida albicans Killing and Preserves Corneal Structure and Function by Controlling Host Antifungal Responses. Infect. Immun. 2015, 83, 4154–4164. [Google Scholar] [CrossRef]
- Liu, X.; Gao, N.; Dong, C.; Zhou, L.; Mi, Q.-S.; Standiford, T.J.; Yu, F.-S.X. Flagellin-Induced Expression of CXCL10 Mediates Direct Fungal Killing and Recruitment of NK Cells to the Cornea in Response to Candida albicans Infection. Eur. J. Immunol. 2014, 44, 2667–2679. [Google Scholar] [CrossRef]
- Gao, N.; Kumar, A.; Guo, H.; Wu, X.; Wheater, M.; Yu, F.-S.X. Topical Flagellin-Mediated Innate Defense against Candida albicans Keratitis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3074–3082. [Google Scholar] [CrossRef]
- Mitchell, B.M.; Wu, T.G.; Jackson, B.E.; Wilhelmus, K.R. Candida albicans Strain-Dependent Virulence and Rim13p-Mediated Filamentation in Experimental Keratomycosis. Investig. Ophthalmol. Vis. Sci. 2007, 48, 774–780. [Google Scholar] [CrossRef]
- Onyewu, C.; Afshari, N.A.; Heitman, J. Calcineurin Promotes Infection of the Cornea by Candida albicans and Can Be Targeted to Enhance Fluconazole Therapy. Antimicrob. Agents Chemother. 2006, 50, 3963–3965. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, K.; Zhao, G. Targeting STAT3 Signaling Alleviates Severity of Fungal Keratitis by Suppressing ICAM-1 and IL-1B. Int. J. Clin. Exp. Med. 2018, 11, 8094–8101. [Google Scholar]
- Dong, C.; Gao, N.; Ross, B.X.; Yu, F.-S.X. ISG15 in Host Defense Against Candida albicans Infection in a Mouse Model of Fungal Keratitis. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2948–2958. [Google Scholar] [CrossRef]
- Qin, X.-H.; Ma, X.; Fang, S.-F.; Zhang, Z.-Z.; Lu, J.-M. IL-17 Produced by Th17 Cells Alleviates the Severity of Fungal Keratitis by Suppressing CX43 Expression in Corneal Peripheral Vascular Endothelial Cells. Cell Cycle 2019, 18, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.G.; Keasler, V.V.; Mitchell, B.M.; Wilhelmus, K.R. Immunosuppression Affects the Severity of Experimental Fusarium solani Keratitis. J. Infect. Dis. 2004, 190, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.M.; Wu, T.G.; Chong, E.-M.; Pate, J.C.; Wilhelmus, K.R. Expression of Matrix Metalloproteinases 2 and 9 in Experimental Corneal Injury and Fungal Keratitis. Cornea 2007, 26, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, H.; Yue, J.; Liu, S.; Li, Z.; Wang, L. Antimicrobial Efficacy of Corneal Cross-Linking in Vitro and in Vivo for Fusarium solani: A Potential New Treatment for Fungal Keratitis. BMC Ophthalmol. 2018, 18, 65. [Google Scholar] [CrossRef]
- Li, X.; Yuan, M.; Yin, R.; Liu, X.; Zhang, Y.; Sun, S.; Han, L.; He, S. Histone Deacetylase Inhibitor Attenuates Experimental Fungal Keratitis in Mice. Sci. Rep. 2019, 9, 9859. [Google Scholar] [CrossRef]
- He, S.; Zhang, H.; Liu, S.; Liu, H.; Chen, G.; Xie, Y.; Zhang, J.; Sun, S.; Li, Z.; Wang, L. Γδ T Cells Regulate the Expression of Cytokines but Not the Manifestation of Fungal Keratitis. Exp. Eye Res. 2015, 135, 93–101. [Google Scholar] [CrossRef]
- Lee, M.H.; Wiedman, G.; Park, S.; Mustaev, A.; Zhao, Y.; Perlin, D.S. A Novel, Tomographic Imaging Probe for Rapid Diagnosis of Fungal Keratitis. Med. Mycol. 2018, 56, 796–802. [Google Scholar] [CrossRef]
- Zhong, J.; Peng, L.; Wang, B.; Zhang, H.; Li, S.; Yang, R.; Deng, Y.; Huang, H.; Yuan, J. Tacrolimus Interacts with Voriconazole to Reduce the Severity of Fungal Keratitis by Suppressing IFN-Related Inflammatory Responses and Concomitant FK506 and Voriconazole Treatment Suppresses Fungal Keratitis. Mol. Vis. 2018, 24, 187–200. [Google Scholar]
- Liang, L.; Shi, W.-Y. The Effects of FK506 Combined with Natamycin in the Treatment of Experimental Fungal Keratitis by Suppressing NLRP3 Inflammasome Activation. Eur. J. Inflamm. 2017, 15, 192–199. [Google Scholar] [CrossRef]
- Yuan, K.; Zhao, G.; Che, C.; Li, C.; Lin, J.; Zhu, G.; He, K. Dectin-1 Is Essential for IL-1β Production through JNK Activation and Apoptosis in Aspergillus fumigatus Keratitis. Int. Immunopharmacol. 2017, 52, 168–175. [Google Scholar] [CrossRef]
- Zhang, S.; Meng, P.; Liu, G.; Liu, K.; Che, C. ATF4 Involvement in TLR4 and LOX-1-Induced Host Inflammatory Response to Aspergillus fumigatus Keratitis. J. Ophthalmol. 2018, 2018, 5830202. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Abbondante, S.; Karmakar, M.; de Jesus Carrion, S.; Che, C.; Hise, A.G.; Pearlman, E. Neutrophil Caspase-11 Is Required for Cleavage of Caspase-1 and Secretion of IL-1β in Aspergillus fumigatus Infection. J. Immunol. 2018, 201, 2767–2775. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Zhao, G.-Q.; Lin, J.; Hu, L.-T.; Che, C.-Y.; Li, C.; Wang, Q.; Xu, Q.; Zhang, J.; Peng, X.-D. Expression of Indoleamine 2,3-Dioxygenase in a Murine Model of Aspergillus fumigatus Keratitis. Int. J. Ophthalmol. 2016, 9, 491–496. [Google Scholar] [CrossRef]
- Liu, M.; Li, C.; Zhao, G.-Q.; Lin, J.; Che, C.-Y.; Xu, Q.; Wang, Q.; Xu, R.; Niu, Y.-W. Boxb Mediate BALB/c Mice Corneal Inflammation through a TLR4/MyD88-Dependent Signaling Pathway in Aspergillus fumigatus Keratitis. Int. J. Ophthalmol. 2018, 11, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chandra, J.; Mukherjee, P.; Szczotka-Flynn, L.; Ghannoum, M.A.; Pearlman, E. A Murine Model of Contact Lens–Associated Fusarium Keratitis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-F.; Zhong, J.; Chen, G.-P.; Lin, Z.-T.; Deng, Y.; Liu, Y.-L.; Cao, P.-Y.; Wang, B.; Wei, Y.; Wu, T.; et al. A Hydrogel-Based Hybrid Theranostic Contact Lens for Fungal Keratitis. ACS Nano 2016, 10, 6464–6473. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Y.; Wang, S.; Qin, F.; Wang, L. MSCs Helped Reduce Scarring in the Cornea after Fungal Infection When Combined with Anti-Fungal Treatment. BMC Ophthalmol. 2019, 19, 226. [Google Scholar] [CrossRef]
- Taylor, P.R.; Leal, S.M.; Sun, Y.; Pearlman, E. Aspergillus and Fusarium Corneal Infections Are Regulated by Th17 Cells and IL-17–Producing Neutrophils. J. Immunol. 2014, 192, 3319–3327. [Google Scholar] [CrossRef]
- Zhao, W.; Che, C.; Liu, K.; Zhang, J.; Jiang, N.; Yuan, K.; Zhao, G. Fenretinide Inhibits Neutrophil Recruitment and IL-1β Production in Aspergillus fumigatus Keratitis. Cornea 2018, 37, 1579–1585. [Google Scholar] [CrossRef]
- Watanabe, H.; Numata, K.; Ito, T.; Takagi, K.; Matsukawa, A. Innate Immune Response In Th1- and Th2-Dominant Mouse Strains. Shock 2004, 22, 460–466. [Google Scholar] [CrossRef]
- Kaiko, G.E.; Horvat, J.C.; Beagley, K.W.; Hansbro, P.M. Immunological Decision-Making: How Does the Immune System Decide to Mount a Helper T-Cell Response? Immunology 2008, 123, 326–338. [Google Scholar] [CrossRef] [PubMed]
- O’Dea, E.M.; Amarsaikhan, N.; Li, H.; Downey, J.; Steele, E.; Van Dyken, S.J.; Locksley, R.M.; Templeton, S.P. Eosinophils are recruited in response to chitin exposure and enhance Th2-mediated immune pathology in Aspergillus fumigatus infection. Infect. Immun. 2014, 82, 3199–3205. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.; Giovannini, G.; De Luca, A.; D’Angelo, C.; Casagrande, A.; Iannitti, R.G.; Ricci, G.; Cunha, C.; Romani, L. Dectin-1 Isoforms Contribute to Distinct Th1/Th17 Cell Activation in Mucosal Candidiasis. Cell. Mol. Immunol. 2012, 9, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Rebong, R.A.; Santaella, R.M.; Goldhagen, B.E.; Majka, C.P.; Perfect, J.R.; Steinbach, W.J.; Afshari, N.A. Polyhexamethylene biguanide and calcineurin inhibitors as novel antifungal treatments for Aspergillus keratitis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7309–7315. [Google Scholar] [CrossRef] [PubMed]
- Abou Shousha, M.; Santos, A.R.; Oechsler, R.; Iovieno, A.; Maestre-Mesa, J.; Ruggeri, M.; Echegaray, J.; Dubovy, S.; Perez, V.; Miller, D.; et al. A Novel Rat Contact Lens Model for Fusarium Keratitis. Mol. Vis. 2013, 19, 2596–2605. [Google Scholar]
- Zhang, Z.-H.; Teng, F.; Sun, Q.-X.; Wang, S.-Z.; Liu, C.; Zhao, G.-Q. Rapamycin Liposome Gutta Inhibiting Fungal Keratitis of Rats. Int. J. Ophthalmol. 2019, 12, 536–541. [Google Scholar] [CrossRef]
- Vasseneix, C.; Gargala, G.; François, A.; Hellot, M.F.; Duclos, C.; Muraine, M.; Benichou, J.; Ballet, J.J.; Brasseur, G.; Favennec, L. A Keratitis Rat Model for Evaluation of Anti-Acanthamoeba polyphaga Agents. Cornea 2006, 25, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, S.M.; Rao, C.M. Condition Responsive Nanoparticles for Managing Infection and Inflammation in Keratitis. Nanoscale 2017, 9, 9946–9959. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, G.-Q.; Qu, J.; Lin, J.; Che, C.-Y.; Yang, X.-J. Early Expression of PTX3 in Aspergillus fumigatus Infected Rat Cornea. Int. J. Ophthalmol. 2018, 11, 1084–1089. [Google Scholar] [CrossRef]
- Zhao, G.; Xu, Q.; Lin, J.; Chen, W.; Cui, T.; Hu, L.; Jiang, N. The Role of Mincle in Innate Immune to Fungal Keratitis. J. Infect. Dev. Ctries 2017, 11, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wu, X.Y.; Yu, F.S. Activation of Toll-like receptors 2 and 4 in Aspergillus fumigatus keratitis. Innate Immun. 2009, 15, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Zernii, E.Y.; Baksheeva, V.E.; Iomdina, E.N.; Averina, O.A.; Permyakov, S.E.; Philippov, P.P.; Zamyatnin, A.A., Jr.; Senin, I.I. Rabbit Models of Ocular Diseases: New Relevance for Classical Approaches. CNS Neurol. Disord. Drug Targets 2016, 15, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.L.; Herreras, J.M.; Dios, E.; Argueso, P.; Almaraz, A. Evaluation of Lectin Staining in the Diagnosis of Fungal Keratitis in an Experimental Rabbit Model. Mol. Vis. 2002, 8, 10–16. [Google Scholar] [PubMed]
- Ghosh, A.K.; Rudramurthy, S.M.; Gupta, A.; Choudhary, H.; Singh, S.; Thakur, A.; Jatana, M. Evaluation of Liposomal and Conventional Amphotericin B in Experimental Fungal Keratitis Rabbit Model. Transl. Vis. Sci. Technol. 2019, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.A.; Takata, T.T.; Shiguematsu, A.I.; Melo Júnior, L.A.S.; Gompertz, O.F.; de Sousa, L.B.; Mannis, M.J. Effect of Topical 0.5% Povidone-Iodine Compared to 5% Natamycin in Fungal Keratitis Caused by Fusarium solani in a Rabbit Model: A Pilot Study. Arq. Bras. Oftalmol. 2008, 71, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Goldblum, D.; Frueh, B.E.; Sarra, G.-M.; Katsoulis, K.; Zimmerli, S. Topical Caspofungin for Treatment of Keratitis Caused by Candida albicans in a Rabbit Model. Antimicrob. Agents Chemother. 2005, 49, 1359–1363. [Google Scholar] [CrossRef]
- Qu, L.; Li, L.; Xie, H. Toxicity and Pharmacokinetics of Intrastromal Injection of Amphotericin B in a Rabbit Model. Curr. Eye Res. 2014, 39, 340–347. [Google Scholar] [CrossRef]
- Sponsel, W.; Chen, N.; Dang, D.; Paris, G.; Graybill, J.; Najvar, L.K.; Zhou, L.; Lam, K.-W.; Glickman, R.; Scribbick, F. Topical Voriconazole as a Novel Treatment for Fungal Keratitis. Antimicrob. Agents Chemother. 2006, 50, 262–268. [Google Scholar] [CrossRef]
- Avunduk, A.M.; Beuerman, R.W.; Warnel, E.D.; Kaufman, H.E.; Greer, D. Comparison of Efficacy of Topical and Oral Fluconazole Treatment in Experimental Aspergillus Keratitis. Curr. Eye Res. 2003, 26, 113–117. [Google Scholar] [CrossRef]
- Galperin, G.; Berra, M.; Tau, J.; Boscaro, G.; Zarate, J.; Berra, A. Treatment of Fungal Keratitis from Fusarium Infection by Corneal Cross-Linking. Cornea 2012, 31, 176–180. [Google Scholar] [CrossRef]
- Hao, Z.-Q.; Song, J.-X.; Pan, S.-Y.; Zhang, L.; Cheng, Y.; Liu, X.-N.; Wu, J.; Xiao, X.-H.; Gao, W.; Zhu, H.-F. Corneal Collagen Cross-Linking and Liposomal Amphotericin B Combination Therapy for Fungal Keratitis in Rabbits. Int. J. Ophthalmol. 2016, 9, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, W.; Gao, M.; Belin, M.W.; Yu, H.; Yu, J. Experimental Study on Cryotherapy for Fungal Corneal Ulcer. BMC Ophthalmol. 2015, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Pinnock, A.; Shivshetty, N.; Roy, S.; Rimmer, S.; Douglas, I.; MacNeil, S.; Garg, P. Ex Vivo Rabbit and Human Corneas as Models for Bacterial and Fungal Keratitis. Graefes Arch. Clin. Exp. Ophthalmol. 2017, 255, 333–342. [Google Scholar] [CrossRef]
- Hua, X.; Yuan, X.; Di Pietro, A.; Wilhelmus, K.R. The Molecular Pathogenicity of Fusarium Keratitis. Cornea 2010, 29, 1440–1444. [Google Scholar] [CrossRef] [PubMed]
- Madhu, S.N.; Jha, K.K.; Karthyayani, A.P.; Gajjar, D.U. Ex vivo caprine model to study virulence factors in keratitis. J. Ophthalmic Vis. Res. 2018, 13, 383–391. [Google Scholar] [PubMed]
- Hua, X.; Yuan, X.; Wilhelmus, K.R. A Fungal PH-Responsive Signaling Pathway Regulating Aspergillus Adaptation and Invasion into the Cornea. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1517–1523. [Google Scholar] [CrossRef]
- Blanco, A.R.; Nostro, A.; D’Angelo, V.; D’Arrigo, M.; Mazzone, M.G.; Marino, A. Efficacy of a Fixed Combination of Tetracycline, Chloramphenicol, and Colistimethate Sodium for Treatment of Candida albicans Keratitis. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4292–4298. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, J.M.; Caballero-Lima, D.; Hillarby, M.C.; Shawcross, S.G.; Brahma, A.; Carley, F.; Read, N.D.; Radhakrishnan, H. Evaluation of Corneal Cross-Linking for Treatment of Fungal Keratitis: Using Confocal Laser Scanning Microscopy on an Ex Vivo Human Corneal Model. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6367–6373. [Google Scholar] [CrossRef]
- Li, C.; Zhao, G.-Q.; Che, C.-Y.; Li, N.; Lin, J.; Xu, Q.; Wang, Q.; Liu, Y.; Qiu, S. Expression of Dectin-1 during Fungus Infection in Human Corneal Epithelial Cells. Int. J. Ophthalmol. 2014, 7, 34–37. [Google Scholar] [CrossRef]
- Zhu, C.-C.; Zhao, G.-Q.; Lin, J.; Hu, L.-T.; Xu, Q.; Peng, X.-D.; Wang, X.; Qiu, S. Dectin-1 Agonist Curdlan Modulates Innate Immunity to Aspergillus fumigatus in Human Corneal Epithelial Cells. Int. J. Ophthalmol. 2015, 8, 690–696. [Google Scholar] [CrossRef]
- Li, C.; Zhao, G.; Che, C.; Lin, J.; Li, N.; Hu, L.; Jiang, N.; Liu, Y. The Role of LOX-1 in Innate Immunity to Aspergillus fumigatus in Corneal Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3593–3603. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.-Q.; Lin, J.; Hu, L.-T.; Yin, X.-N.; Wang, Q.; Xu, Q.; Li, H. The Role of Dectin-1/Raf-1 Signal Cascade in Innate Immune of Human Corneal Epithelial Cells against Aspergillus fumigatus Infection. Int. J. Ophthalmol. 2016, 9, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.-Q.; Qiu, X.-Y.; Lin, J.; Li, Q.; Hu, L.-T.; Wang, Q.; Li, H. Co-Regulation of Dectin-1 and TLR2 in Inflammatory Response of Human Corneal Epithelial Cells Induced by Aspergillus fumigatus. Int. J. Ophthalmol. 2016, 9, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Chi, W.; Su, L.; Li, J.; Zhang, Z.; Yuan, X. ROS-Induced Oxidative Injury Involved in Pathogenesis of Fungal Keratitis via P38 MAPK Activation. Sci. Rep. 2017, 7, 10421. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, A.; Swioklo, S.; Connon, C.J. 3D Bioprinting of a Corneal Stroma. Equivalent. Exp. Eye Res. 2018, 17, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, S.; Nicholas, S.E.; Karamichos, D. 3D Stacked Construct: A Novel Substitute for Corneal Tissue Engineering. Methods Mol. Biol. 2018, 1697, 173–180. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fungal Species (Strain ID) | Mouse Strain | Inoculation Procedure | Inoculum Size | Time Points | Immune Status | Cite # |
|---|---|---|---|---|---|---|
| A. fumigatus (3.0772) | C57BL/6, BALB/c | Central cornea scraped, inoculum added, soft contact lens added, then eyelids sutured. | 5 μL of 1 × 108 CFU/mL | day 1, 3, and 5 p.i. | Immunocompetent | [31] |
| A. fumigatus (3.0772) | C57BL/6 | 30G needle used to create tunnel into cornea stroma, then 33G Hamilton syringe was inserted into tunnel and inoculum injected. | 2 μL of 1 × 105 solution | day 1, 3, and 5 p.i. | Immunocompetent | [32] |
| C. albicans (ATCC 32354) | NIH Swiss, BALB/c | Hypodermic needle used to scarify cornea surface with 30 scratches in a crosshatching grid formation, inoculum applied. | 5 μL containing 102, 104, 106, or 108 CFU | 6 h; day 1, 4, and 8 p.i. | Immunocompromised with methylprednisolone and cyclophosphamide | [33] |
| C. albicans (SC5314) | C57BL/6 | Corneal surface scratched 3 times, 2 mm deep, with a 26G needle, then inoculum added. | 5 μL of 1 × 105 CFU solution | 6 h; day 1 and 3 p.i. | Immunocompetent | [34] |
| C. albicans (SC5314) | C57BL/6, C57BL/6.129S7-Rag1tm1Mom/J | Corneal surface scratched 3 times with 1 mm incisions using a 26G needle, inoculum added | 5 μL containing 1 × 105 CFU | day 1, 3, 5, and 7 p.i. | Immunocompetent | [35] |
| C. albicans (SC5314) | C57BL/6, C57BL/6 TR5 −/−, C57BL/6 Camp -/- | Corneal surface scratched 3 times with 1 mm incisions using a 26G needle, inoculum added | 5 μL of 1 × 104 to 106 CFU | 6 h; day 1, 3, and 5 p.i. | Immunocompetent | [36] |
| C. albicans (ATCC 32354. SC5314, VE175, Tn7-rim13, BWP17, DAY28) | BALB/c | Hypodermic needle used to scarify cornea surface with 30 scratches in a crosshatching grid formation, inoculum applied. | 5 μL containing 1 × 105 or 1 · 106 CFU | 6 h, day 1, 4, and 8 p.i. | Immunocompromised with cyclophosphamide | [37] |
| C. albicans (SC5314) | BALB/c, C57BL/6 | Hypodermic needle used to scarify cornea surface with 30 scratches in a crosshatching grid formation, inoculum applied. | 5 μL containing 1 × 106 CFU | day 1, 3, and 7 p.i. | Immunocompetent | [27] |
| C. albicans (SC5314) | BALB/c | Corneal surface scarified with a 28.5G needle, inoculum applied. | 5 μL containing 1 ×106 CFU | day 1, 2, 3, and 4 p.i. | Immunosuppressed with methylprednisolone | [38] |
| C. albicans | C57BL/6 | Hypodermic needle used to scarify cornea surface with 30 scratches in a crosshatching grid formation, inoculum applied. | 5 μL containing 1 × 106 CFU | day 1 and 5 p.i. | Immunocompetent | [39] |
| C. albicans (SC5314) | C57BL/6 | Corneal surface scratched 3 times with 1mm incisions using a 26G needle, inoculum added. | 5 μL containing 1 × 105 CFU | day 1, 3, 5, and 7 p.i. | Immunocompetent | [40] |
| C. albicans (MYA-2876) | BALB/c | Hypodermic needle used to scarify cornea surface with 30 scratches in a crosshatching grid formation, inoculum applied. | 5 μL containing 1 ×106 CFU | day 1–8 p.i. | Immunocompetent | [41] |
| F. solani (SRL-F2) | BALB/c | Hypodermic needle used to scarify cornea surface with 30 scratches in a crosshatching grid formation, inoculum applied. | 5 μL containing 1 × 101, 1 × 104, or 1 × 105 CFU | 6 h; days 1, 4, 8, and 14 p.i. | Immunosuppressed with cyclophosphamide | [42] |
| F. solani (SRL-F2) | BALB/c | Hypodermic needle used to scarify cornea surface with 30 scratches in a crosshatching grid formation, inoculum applied. | 5 μL containing 1 × 105 CFU | 1.5 and 6 h; day 1, 4, and 8 p.i. | Immunosuppressed with cyclophosphamide | [43] |
| F. solani (No.3.1791) | C57BL/6 | a 2mm trephine marked the central cornea and a sterile scalpel blade scratched the area. A sharpened bamboo toothpick scraped the area 2-3 times and fungi was then smeared onto the central cornea. | Fungal hyphae ground with a glass rod for fungal suspension, solution adjusted by turbidimeter to get 0.5 Mx suspenion. | day 1, 3, 7, and 10 p.i. | Immunocompetent | [44] |
| F. solani cultures from Henan Eye Institute | C57BL/6 | a 2mm trephine marked the central cornea and a sterile scalpel blade scratched the area. A sharpened bamboo toothpick was used to smear fungi onto the central cornea. | Not indicated. | Sacrificed after formation of corneal lesion; varied. | Immunocompetent | [45] |
| F. solani | C57BL/6 | a 2mm trephine marked the central cornea and a sterile scalpel blade scratched the area. A sharpened bamboo toothpick scraped the area 2-3 times and fungi was then smeared onto the central cornea. | Not indicated. | 6, 12, 18, 24, 36, 72, and 120 h p.i. | Immunocompetent | [46] |
| A. fumigatus (R21), A. flavus (DPL9), A. niger (DPL29), F. solani (DPL114) | SKH1 | 30G needle used to create tunnel into cornea stroma, then 33G Hamilton syringe was inserted into tunnel and inoculum injected. | 2 μL containing 5 × 106 CFU of A. fumigatus, A. flavus, A. niger, or F. solani | 24 h p.i. | Immunocompetent | [47] |
| A. fumigatus (3.0772) | C57BL/6 | 30G needle used to create tunnel into cornea stroma, then 33G Hamilton syringe was inserted into tunnel and inoculum injected. | 2 μL containing 1 × 105 CFU | 8 h; day 1, 3, 5, and 6 p.i. | Immunocompetent | [48] |
| A. flavus and F. solani | C57BL/6 | 30G needle used to abrade the corneal surface, then 33G Hamilton syringe was inserted into the stroma and inoculum injected. | 2 μL containing 1 × 105 CFU | 48 h p.i. | Immunocompetent | [49] |
| A. fumigatus (3.0772) | C57BL/6 | 30G needle used to abrade the corneal surface, then 33G Hamilton syringe was inserted into the stroma and inoculum injected. | 2 μL of a 5 × 104 conidial/μL solution | 12 h; day 1 and 2 p.i. | Immunocompetent | [50] |
| A. fumigatus (3.0772) | C57BL/6 | Central cornea scratched with a 255/8G needle and inoculum added. A soft contact lens was applied to the corneal surface and the eyelids were sutured shut. | 5 μL of 1 × 108 conidia/μL | 12 h; days 1–3, 5, 7, 10, and 14 p.i. | Immunocompetent | [51] |
| A. fumigatus (Af293) | C57BL/6, caspase-1/11 −/−, caspase-11 −/−, IFNAR1 −/−, Dectin-1 −/−, IL-1β −/−, NLRP3 −/−, and ASC −/− | 30G needle used to create tunnel into cornea stroma, then 33G Hamilton syringe was inserted into tunnel and inoculum injected. | 2 μL containing 1 × 105 CFU | day 1 and 2 p.i. | Immunocompetent | [52] |
| A. fumigatus (3.0772) | C57BL/6 | Central cornea was scraped with 30G needle then smeared with fungal colonies. It was then covered with a contact lens and eyelids were sutured shut. Contact lenses were removed after 24 h. | Not indicated. | day 1, 3, and 5 p.i. | Immunocompetent | [53] |
| A. fumigatus (3.0772) | BALB/c | A 2mm scratch was made to the central cornea, covered in inoculum, and contact lens placed. Eyelids were sewn shut. | Not indicated. | day 1 p.i. | Immunocompetent | [54] |
| F. oxysporum (MRL8996) | C57BL/6, IL-1R1 −/−, TLR2 −/−, TLR4 −/−, MYD88 −/− | Lotrafilcon A contact lenses were incubated with conidia, washed, and then incubated further to establish biofilm. The central cornea was abraded and a 2mm punch of the biolfilm contact lens was placed on the central cornea. After 2 h, the contact lenses were removed. | 1 × 106 conidia were incubated with the contact lenses. | 90 min; 2, 24, and 48 h; day 1, 2, 3, and 4 p.i. | Immunosuppressed with cyclophosphamide | [55] |
| F. solani, A. fumigatus (AS 3.772) | C57BL/6 | 30G needle used to create tunnel into cornea stroma, then 33G Hamilton syringe was inserted into tunnel and inoculum injected. After 12 h, a contact lens fragment was placed on the eye. | 2 μL of a 1 × 108 CFU/mL suspension | day 1, 3, and 5 p.i. | Immunocompetent | [56] |
| F. oxysporum (No.3.791) | C57BL/6 | a 2mm trephine marked the central cornea and a sterile scalpel blade scratched the area. A sharpened bamboo toothpick scraped the area 2-3 times and fungi was then smeared onto the central cornea. | Not indicated. | day 14, 21, and 28 p.i. | Immunocompetent | [57] |
| A. fumigatus (Af-BP), F. oxysporum (8996) | C57BL/6. Rag2 −/−, IL17 −/− | 30G needle used to abrade the corneal surface, then 33G Hamilton syringe was inserted into the stroma and inoculum injected. | 2 μL containing 1 × 105 conidia | 24, 48, and 72 h p.i. | Immunocompetent | [58] |
| A. fumigatus (3.0772) | C57BL/6 | Cornea stromas were injected with inoculum. | Volume not specified, 0.5 × 105 conidia/μL | day 1 p.i. | Immunocomptent | [59] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montgomery, M.L.; Fuller, K.K. Experimental Models for Fungal Keratitis: An Overview of Principles and Protocols. Cells 2020, 9, 1713. https://doi.org/10.3390/cells9071713
Montgomery ML, Fuller KK. Experimental Models for Fungal Keratitis: An Overview of Principles and Protocols. Cells. 2020; 9(7):1713. https://doi.org/10.3390/cells9071713
Chicago/Turabian StyleMontgomery, Micaela L., and Kevin K. Fuller. 2020. "Experimental Models for Fungal Keratitis: An Overview of Principles and Protocols" Cells 9, no. 7: 1713. https://doi.org/10.3390/cells9071713
APA StyleMontgomery, M. L., & Fuller, K. K. (2020). Experimental Models for Fungal Keratitis: An Overview of Principles and Protocols. Cells, 9(7), 1713. https://doi.org/10.3390/cells9071713