Phosphoinositide Profile of the Mouse Retina

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals
2.3. Phosphatidylinositol Extraction and Detection with Electroconductivity Method
2.4. Phosphatidylinositol Extraction and Detection with Mass Spectrometry
2.5. Distribution of PI(4)P and PI(4,5)P2 in Rod Photoreceptors
2.6. Statistical Analysis
3. Results
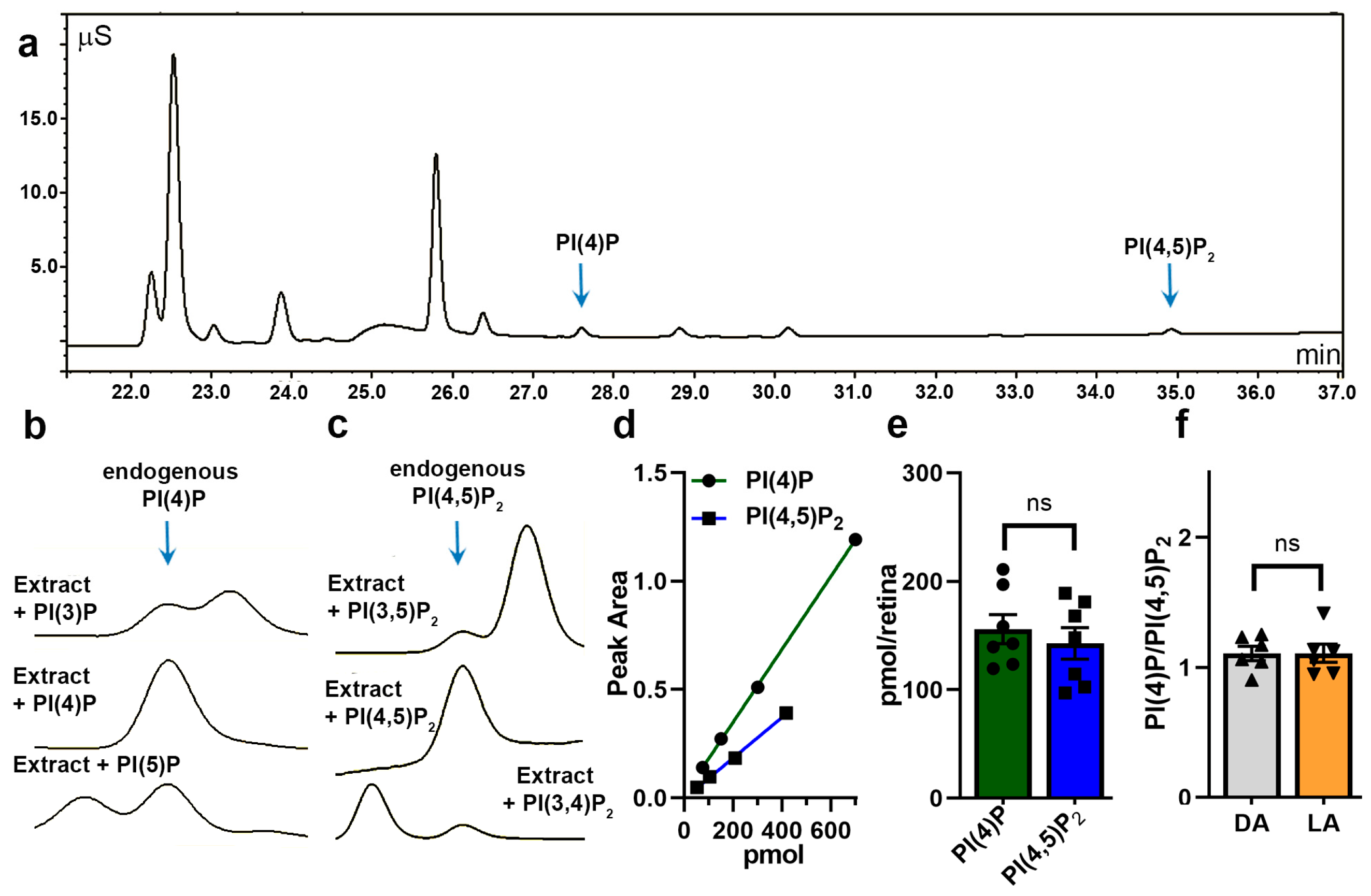
3.1. Retinal Composition of Major Phosphoinositide Headgroups
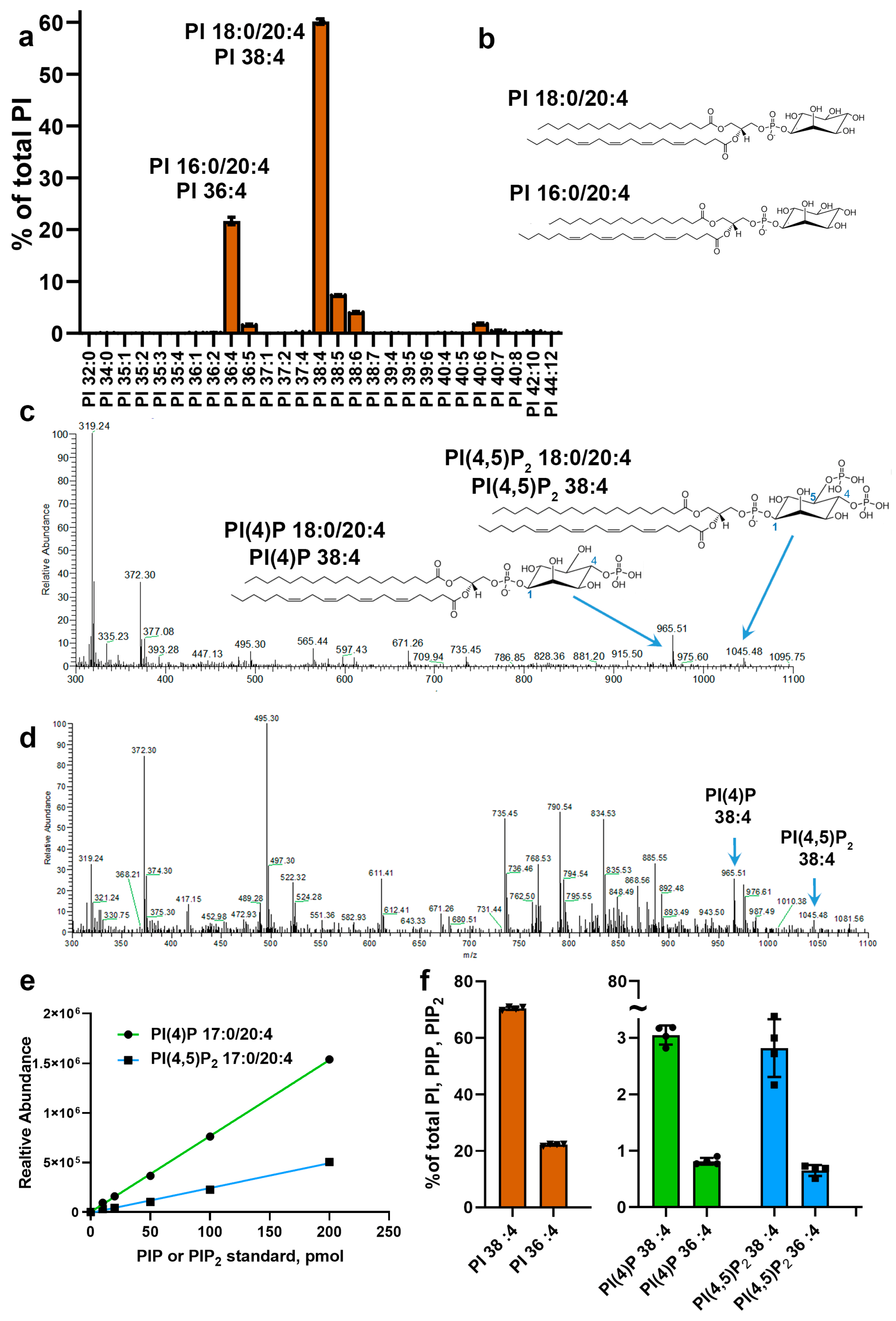
3.2. Fatty Acid Composition of Major Retinal Phosphoinositides
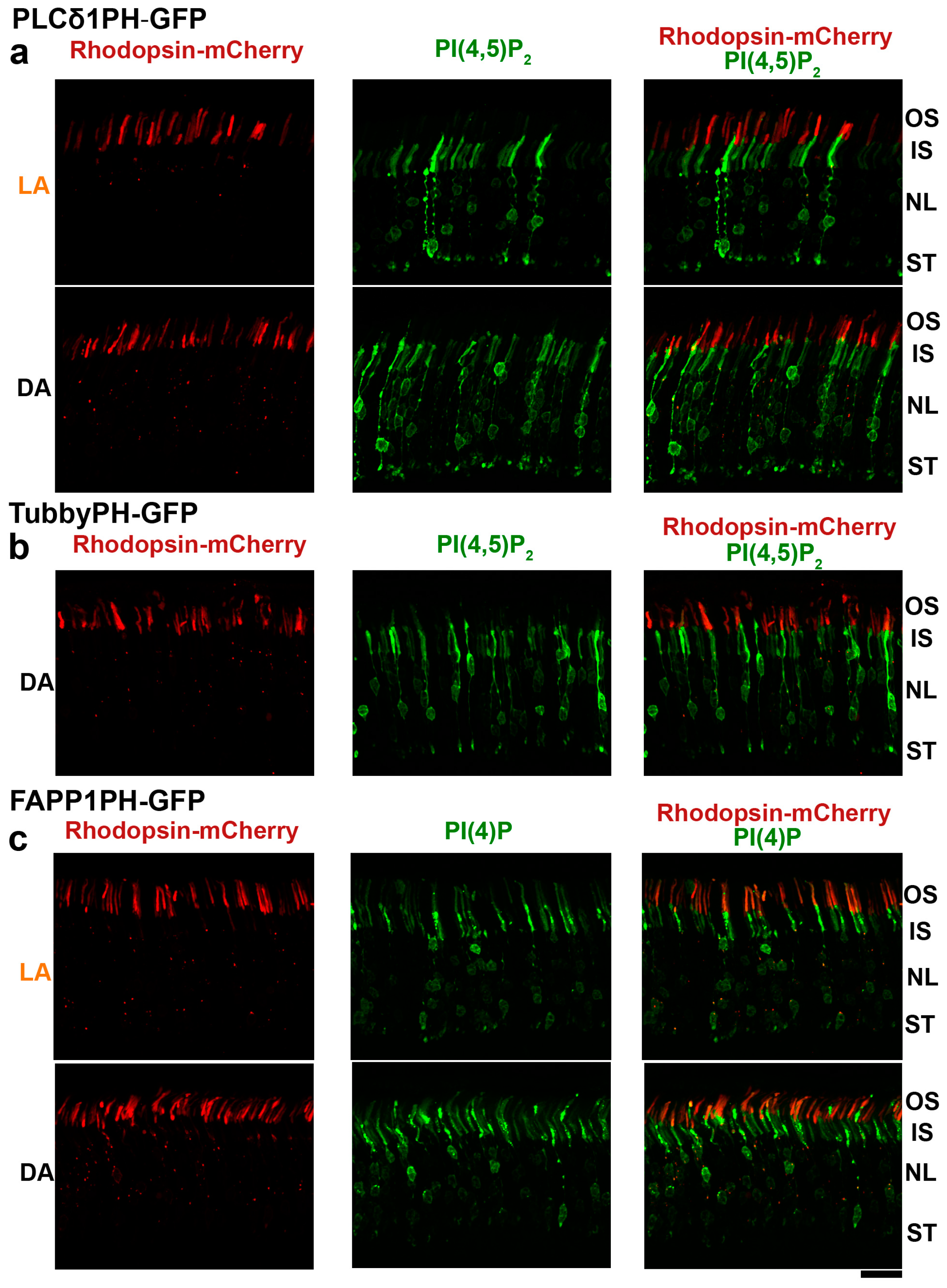
3.3. Subcellular Distribution of PI(4,5)P2 and PI(4)P in Mouse Rod Photoreceptors
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| C | chloroform |
| DA | dark-adapted |
| FAPP1 | four-phosphate-adaptor protein 1 |
| GFP | green fluorescent protein |
| HCl | hydrochloric acid |
| HPLC | high-performance liquid chromatography |
| INPP5E | inositol polyphosphate-5-phosphatase E |
| LA | light-adapted |
| M | methanol |
| NaOH | sodium hydroxide |
| PBS | phosphate-buffered saline |
| PH | pleckstrin homology domain |
| PI | phosphatidylinositol |
| PI(4)P | phosphatidylinositol 4-monophosphate |
| PI(4,5)P2 | phosphatidylinositol 4,5-bisphosphate |
| PI3K | phosphoinositide 3-kinase |
| PLCδ | phospholipase Cδ |
| ROS | rod outer segments |
| SynJ1 | synaptojanin 1 |
| Vps34 | vacuolar protein sorting 34 |
References
- de Craene, J.-O.; Bertazzi, D.L.; Bar, S.; Friant, S. Phosphoinositides, Major Actors in Membrane Trafficking and Lipid Signaling Pathways. Int. J. Mol. Sci. 2017, 18, 634. [Google Scholar] [CrossRef] [PubMed]
- Schink, K.O.; Tan, K.-W.; Stenmark, H. Phosphoinositides in Control of Membrane Dynamics. Annu. Rev. Cell Dev. Boil. 2016, 32, 143–171. [Google Scholar] [CrossRef] [PubMed]
- Balla, T. Phosphoinositides: Tiny lipids with giant impact on cell regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef] [PubMed]
- Hammond, G.R.; Burke, J.E. Novel roles of phosphoinositides in signaling, lipid transport, and disease. Curr. Opin. Cell Boil. 2020, 63, 57–67. [Google Scholar] [CrossRef] [PubMed]
- George, A.A.; Hayden, S.; Holzhausen, L.C.; Ma, E.Y.; Suzuki, S.C.; Brockerhoff, S.E. Synaptojanin 1 Is Required for Endolysosomal Trafficking of Synaptic Proteins in Cone Photoreceptor Inner Segments. PLoS ONE 2014, 9, e84394. [Google Scholar] [CrossRef] [Green Version]
- George, A.A.; Hayden, S.; Stanton, G.R.; Brockerhoff, S.E. Arf6 and the 5’phosphatase of synaptojanin 1 regulate autophagy in cone photoreceptors. Bioessays 2016, 38, 119–135. [Google Scholar] [CrossRef]
- He, F.; Agosto, M.; Anastassov, I.; Tse, D.Y.; Wu, S.M.; Wensel, T.G. Phosphatidylinositol-3-phosphate is light-regulated and essential for survival in retinal rods. Sci. Rep. 2016, 6, 26978. [Google Scholar] [CrossRef]
- Rajala, R.V.S.; Ranjo-Bishop, M.; Wang, Y.; Rajala, A.; Anderson, R.E. The p110α isoform of phosphoinositide 3-kinase is essential for cone photoreceptor survival. Biochimie 2015, 112, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Brooks, B.P.; Zein, W.M.; Thompson, A.; Mokhtarzadeh, M.; Doherty, D.; Parisi, M.; Glass, I.A.; Malicdan, M.C.; Vilboux, T.; Vemulapalli, M.; et al. Joubert Syndrome: Ophthalmological Findings in Correlation with Genotype and Hepatorenal Disease in 99 Patients Prospectively Evaluated at a Single Center. Ophthalmology 2018, 125, 1937–1952. [Google Scholar] [CrossRef] [Green Version]
- Travaglini, L.; Brancati, F.; Silhavy, J.; Iannicelli, M.; Nickerson, E.; Elkhartoufi, N.; Scott, E.; Spencer, E.; Gabriel, S.; Thomas, S.; et al. Phenotypic spectrum and prevalence of INPP5E mutations in Joubert Syndrome and related disorders. Eur. J. Hum. Genet. 2013, 21, 1074–1078. [Google Scholar] [CrossRef] [Green Version]
- Hagstrom, S.A.; Duyao, M.; North, M.A.; Li, T. Retinal degeneration in tulp1-/- mice: Vesicular accumulation in the interphotoreceptor matrix. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2795–2802. [Google Scholar]
- Hagstrom, S.A.; North, M.A.; Nishina, P.M.; Berson, E.L.; Dryja, T.P. Recessive mutations in the gene encoding the tubby-like protein TULP1 in patients with Retinitis pigmentosa. Nat. Genet. 1998, 18, 174–176. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, A.; Naggert, J.K.; Nishina, P.M. Genetic Modification of Retinal Degeneration in Tubby Mice. Exp. Eye Res. 2002, 74, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Chuang, J.-Z.; Zhao, Y.; Sung, C.-H. SARA-regulated vesicular targeting underlies formation of the light-sensing organelle in mammalian rods. Cell 2007, 130, 535–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deretic, D.; Traverso, V.; Parkins, N.; Jackson, F.; de Turco, E.B.R.; Ransom, N. Phosphoinositides, Ezrin/Moesin, and rac1 Regulate Fusion of Rhodopsin Transport Carriers in Retinal Photoreceptors. Mol. Boil. Cell 2004, 15, 359–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giusto, N.; Pasquaré, S.J.; A Salvador, G.; I Castagnet, P.; E Roque, M.; De Boschero, M.G.I. Lipid metabolism in vertebrate retinal rod outer segments. Prog. Lipid Res. 2000, 39, 315–391. [Google Scholar] [CrossRef]
- Wensel, T.G. Phosphoinositides in Retinal Function and Disease. Cells 2020, 9, 866. [Google Scholar] [CrossRef] [Green Version]
- Lobanova, E.S.; Finkelstein, S.; Li, J.; Travis, A.M.; Hao, Y.; Klingeborn, M.; Skiba, N.P.; Deshaies, R.J.; Arshavsky, V.Y. Increased proteasomal activity supports photoreceptor survival in inherited retinal degeneration. Nat. Commun. 2018, 9, 1738. [Google Scholar] [CrossRef]
- Lobanova, E.S.; Herrmann, R.; Finkelstein, S.; Reidel, B.; Skiba, N.P.; Deng, W.-T.; Jo, R.; Weiss, E.R.; Hauswirth, W.; Arshavsky, V.Y. Mechanistic basis for the failure of cone transducin to translocate: Why cones are never blinded by light. J. Neurosci. 2010, 30, 6815–6824. [Google Scholar] [CrossRef]
- Nasuhoglu, C.; Feng, S.; Mao, J.; Yamamoto, M.; Yin, H.L.; Earnest, S.; Barylko, B.; Albanesi, J.P.; Hilgemann, D.W. Nonradioactive Analysis of Phosphatidylinositides and Other Anionic Phospholipids by Anion-Exchange High-Performance Liquid Chromatography with Suppressed Conductivity Detection. Anal. Biochem. 2002, 301, 243–254. [Google Scholar] [CrossRef]
- Rouser, G.; Fkeischer, S.; Yamamoto, A. Two dimensional then layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 1970, 5, 494–496. [Google Scholar] [CrossRef] [PubMed]
- Milne, S.B.; Ivanova, P.T.; de Camp, D.; Hsueh, R.C.; Brown, H.A. A targeted mass spectrometric analysis of phosphatidylinositol phosphate species. J. Lipid Res. 2005, 46, 1796–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, R.; Schwudke, D.; Schuhmann, K.; Sampaio, J.; Bornstein, S.; Schroeder, M.; Shevchenko, A. A novel informatics concept for high-throughput shotgun lipidomics based on the molecular fragmentation query language. Genome Boil. 2011, 12, R8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobanova, E.S.; Schuhmann, K.; Finkelstein, S.; Lewis, T.; Cady, M.A.; Hao, Y.; Keuthan, C.; Ash, J.D.; Burns, M.E.; Shevchenko, A.; et al. Disrupted Blood-Retina Lysophosphatidylcholine Transport Impairs Photoreceptor Health But Not Visual Signal Transduction. J. Neurosci. 2019, 39, 9689–9701. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Cepko, C.L. Electroporation and RNA interference in the rodent retina in vivo and in vitro. Proc. Natl. Acad. Sci. USA 2003, 101, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Balla, T.; Tuymetova, G.; Tsiomenko, A.; Várnai, P.; Balla, T. A Plasma Membrane Pool of Phosphatidylinositol 4-Phosphate Is Generated by Phosphatidylinositol 4-Kinase Type-III Alpha: Studies with the PH Domains of the Oxysterol Binding Protein and FAPP1. Mol. Boil. Cell 2005, 16, 1282–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, G.R.; Fischer, M.J.; Anderson, K.E.; Holdich, J.; Koteci, A.; Balla, T.; Irvine, R.F. PI4P and PI (4,5) P2 Are Essential but Independent Lipid Determinants of Membrane Identity. Science 2012, 337, 727–730. [Google Scholar] [CrossRef] [Green Version]
- Szentpetery, Z.; Balla, T.; Kim, Y.J.; Lemmon, M.A.; Balla, T. Live cell imaging with protein domains capable of recognizing phosphatidylinositol 4,5-bisphosphate; A comparative study. BMC Cell Boil. 2009, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- Balla, T.; Várnai, P. Visualization of Cellular Phosphoinositide Pools with GFP-Fused Protein-Domains. Curr. Protoc. Cell Boil. 2009, 42. [Google Scholar] [CrossRef] [Green Version]
- Řezanka, T.; Sigler, K. Odd-numbered very-long-chain fatty acids from the microbial, animal and plant kingdoms. Prog. Lipid Res. 2009, 48, 206–238. [Google Scholar] [CrossRef]
- Zhang, L.-S.; Liang, S.; Zong, M.-H.; Yang, J.-G.; Lou, W.-Y. Microbial synthesis of functional odd-chain fatty acids: A review. World J. Microbiol. Biotechnol. 2020, 36, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Wenk, M.R.; Lucast, L.; di Paolo, G.; Romanelli, A.J.; Suchy, S.F.; Nussbaum, R.L.; Cline, G.W.; Shulman, G.I.; McMurray, W.; de Camilli, P. Phosphoinositide profiling in complex lipid mixtures using electrospray ionization mass spectrometry. Nat. Biotechnol. 2003, 21, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Rajala, R.V.S.; Rajala, A.; Morris, A.J.; Anderson, R.E. Phosphoinositides: Minor Lipids Make a Major Impact on Photoreceptor Cell Functions. Sci. Rep. 2014, 4, 5463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Mao, M.; Wensel, T.G. Enhancement of Phototransduction G Protein-Effector Interactions by Phosphoinositides. J. Boil. Chem. 2003, 279, 8986–8990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, R.V.S.; Anderson, R.E. Focus on Molecules: Phosphatidylinositol-4,5-bisphosphate (PIP2). Exp. Eye Res. 2010, 91, 324–325. [Google Scholar] [CrossRef] [Green Version]
- Brockerhoff, E.S. Phosphoinositides and photoreceptors. Mol. Neurobiol. 2011, 44, 420–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghalayini, A.; Anderson, R.E. Phosphatidylinositol 4,5-bisphosphate: Light-mediated breakdown in the vertebrate retina. Biochem. Biophys. Res. Commun. 1984, 124, 503–506. [Google Scholar] [CrossRef]
- Ghalayini, A.J.; Weber, N.R.; Rundle, D.R.; A Koutz, C.; Lambert, D.; Guo, X.X.; E Anderson, R. Phospholipase Cgamma1 in bovine rod outer segments: Immunolocalization and light-dependent binding to membranes. J. Neurochem. 1998, 70, 171–178. [Google Scholar] [CrossRef]
- Ghalayini, A.J.; Anderson, R.E. Light adaptation of bovine retinas in situ stimulates phosphatidylinositol synthesis in rod outer segments in vitro. Curr. Eye Res. 1995, 14, 1025–1029. [Google Scholar] [CrossRef]
- He, F.; Nichols, R.M.; Kailasam, L.; Wensel, T.G.; Agosto, M.A. Critical Role for Phosphatidylinositol-3 Kinase Vps34/PIK3C3 in ON-Bipolar Cells. Invest Ophthalmol. Vis. Sci. 2019, 60, 2861–2874. [Google Scholar] [CrossRef] [Green Version]
- Raghu, P.; Joseph, A.; Krishnan, H.; Singh, P.; Saha, S. Phosphoinositides: Regulators of Nervous System Function in Health and Disease. Front. Mol. Neurosci. 2019, 12, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waugh, M.G. PIPs in neurological diseases. Biochim. Biophys. Acta Mol. Cell Boil. Lipids 2015, 1851, 1066–1082. [Google Scholar] [CrossRef] [PubMed]
- Barneda, D.; Cosulich, S.; Stephens, L.; Hawkins, P. How is the acyl chain composition of phosphoinositides created and does it matter? Biochem. Soc. Trans. 2019, 47, 1291–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, T.; Takatori, S.; Fujimoto, T. Definition of phosphoinositide distribution in the nanoscale. Curr. Opin. Cell Boil. 2019, 57, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Wakelam, M. The uses and limitations of the analysis of cellular phosphoinositides by lipidomic and imaging methodologies. Biochim. Biophys. Acta Mol. Cell Boil. Lipids 2014, 1841, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Mehta, Z.B.; Piętka, G.; Lowe, M. The Cellular and Physiological Functions of the Lowe Syndrome Protein OCRL1. Traffic 2014, 15, 471–487. [Google Scholar] [CrossRef] [Green Version]
- Bujakowska, K.M.; Liu, Q.; A Pierce, E. Photoreceptor Cilia and Retinal Ciliopathies. Cold Spring Harb. Perspect. Boil. 2017, 9, a028274. [Google Scholar] [CrossRef]
- Verbakel, S.K.; van Huet, R.A.; Boon, C.J.; Hollander, A.I.D.; Collin, R.W.; Klaver, C.C.W.; Hoyng, C.; Roepman, R.; Klevering, B.J. Non-syndromic retinitis pigmentosa. Prog. Retin. Eye Res. 2018, 66, 157–186. [Google Scholar] [CrossRef]
- Daiger, S.P.; Sullivan, L.S.; Bowne, S.J. Genes and mutations causing retinitis pigmentosa. Clin. Genet. 2013, 84, 132–141. [Google Scholar] [CrossRef]
- Daiger, S.P.; Bowne, S.J.; Sullivan, L.S. Genes and Mutations Causing Autosomal Dominant Retinitis Pigmentosa. Cold Spring Harb. Perspect Med. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- di Carlo, J.E.; Mahajan, V.B.; Tsang, S.H. Gene therapy and genome surgery in the retina. J. Clin. Investig. 2018, 128, 2177–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baehr, W.; Hanke-Gogokhia, C.; Sharif, A.; Reed, M.; Dahl, T.; Frederick, J.M.; Ying, G. Insights into photoreceptor ciliogenesis revealed by animal models. Prog. Retin. Eye Res. 2018, 71, 26–56. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finkelstein, S.; Gospe, S.M., III; Schuhmann, K.; Shevchenko, A.; Arshavsky, V.Y.; Lobanova, E.S. Phosphoinositide Profile of the Mouse Retina. Cells 2020, 9, 1417. https://doi.org/10.3390/cells9061417
Finkelstein S, Gospe SM III, Schuhmann K, Shevchenko A, Arshavsky VY, Lobanova ES. Phosphoinositide Profile of the Mouse Retina. Cells. 2020; 9(6):1417. https://doi.org/10.3390/cells9061417
Chicago/Turabian StyleFinkelstein, Stella, Sidney M. Gospe, III, Kai Schuhmann, Andrej Shevchenko, Vadim Y. Arshavsky, and Ekaterina S. Lobanova. 2020. "Phosphoinositide Profile of the Mouse Retina" Cells 9, no. 6: 1417. https://doi.org/10.3390/cells9061417
APA StyleFinkelstein, S., Gospe, S. M., III, Schuhmann, K., Shevchenko, A., Arshavsky, V. Y., & Lobanova, E. S. (2020). Phosphoinositide Profile of the Mouse Retina. Cells, 9(6), 1417. https://doi.org/10.3390/cells9061417