Characterization of Plasma-Derived Small Extracellular Vesicles Indicates Ongoing Endothelial and Platelet Activation in Patients with Thrombotic Antiphospholipid Syndrome

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Controls
2.2. Blood Collection
2.3. Biochemical Analysis
2.4. aPL Determination
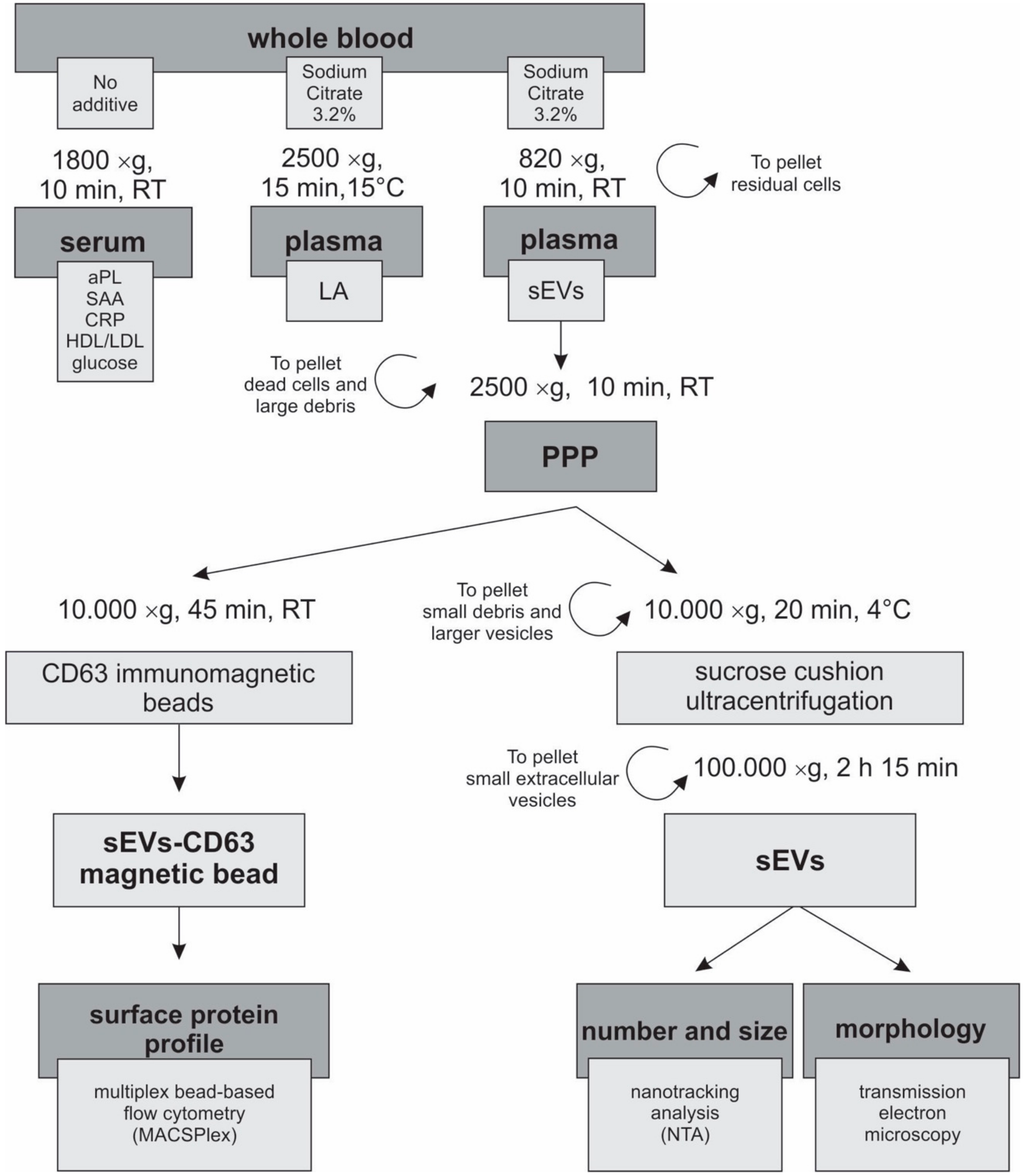
2.5. Preparation of Plasma for EV Isolation
2.6. Sucrose Cushion Ultracentrifugation
2.7. Quantification of the Concentration and Size of the sEVs
2.8. CD63 Immunomagnetic Isolation of sEVs
2.9. Multiplex Bead-Based Flow Cytometry Analysis of the sEVs
2.10. Transmission Electron Microscopy
2.11. Statistical Analysis
3. Results
3.1. Subject Characteristics That Could Confound the sEV Analysis Were Mostly Comparable between Study Groups
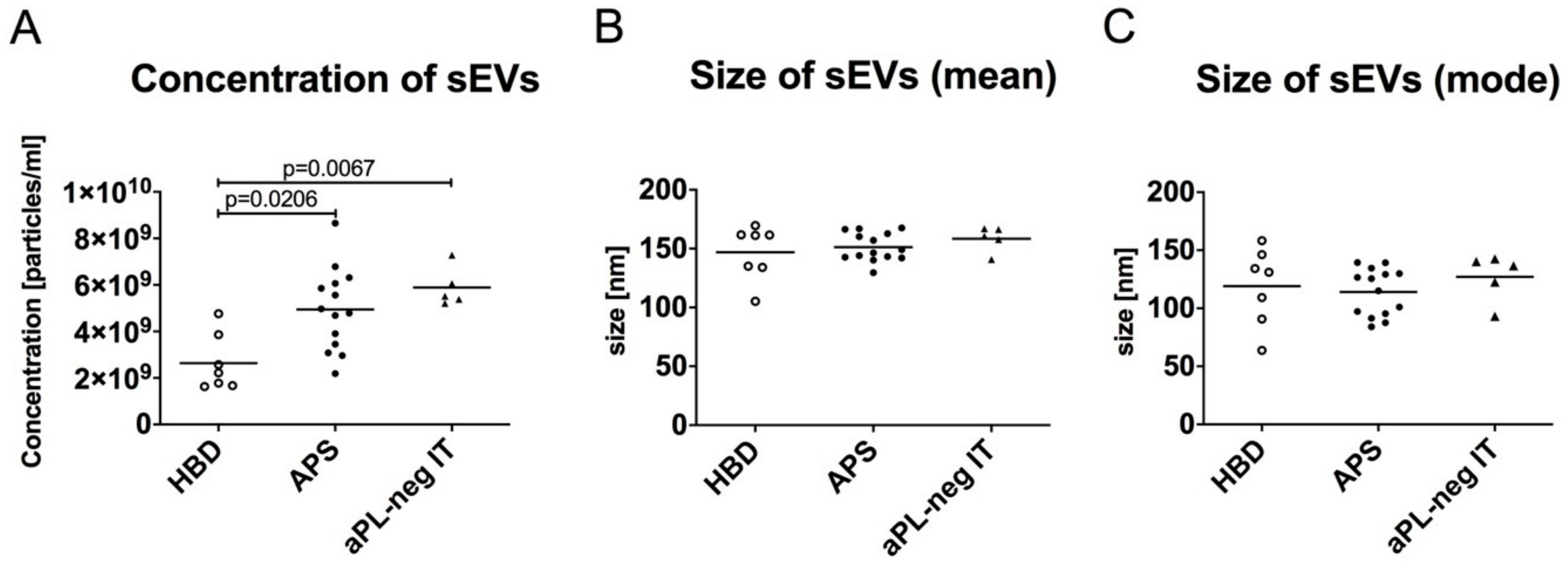
3.2. The Quantity of sEVs Is Increased in Patients with APS-Associated Thrombosis and aPL-Negative Idiopathic Thrombosis
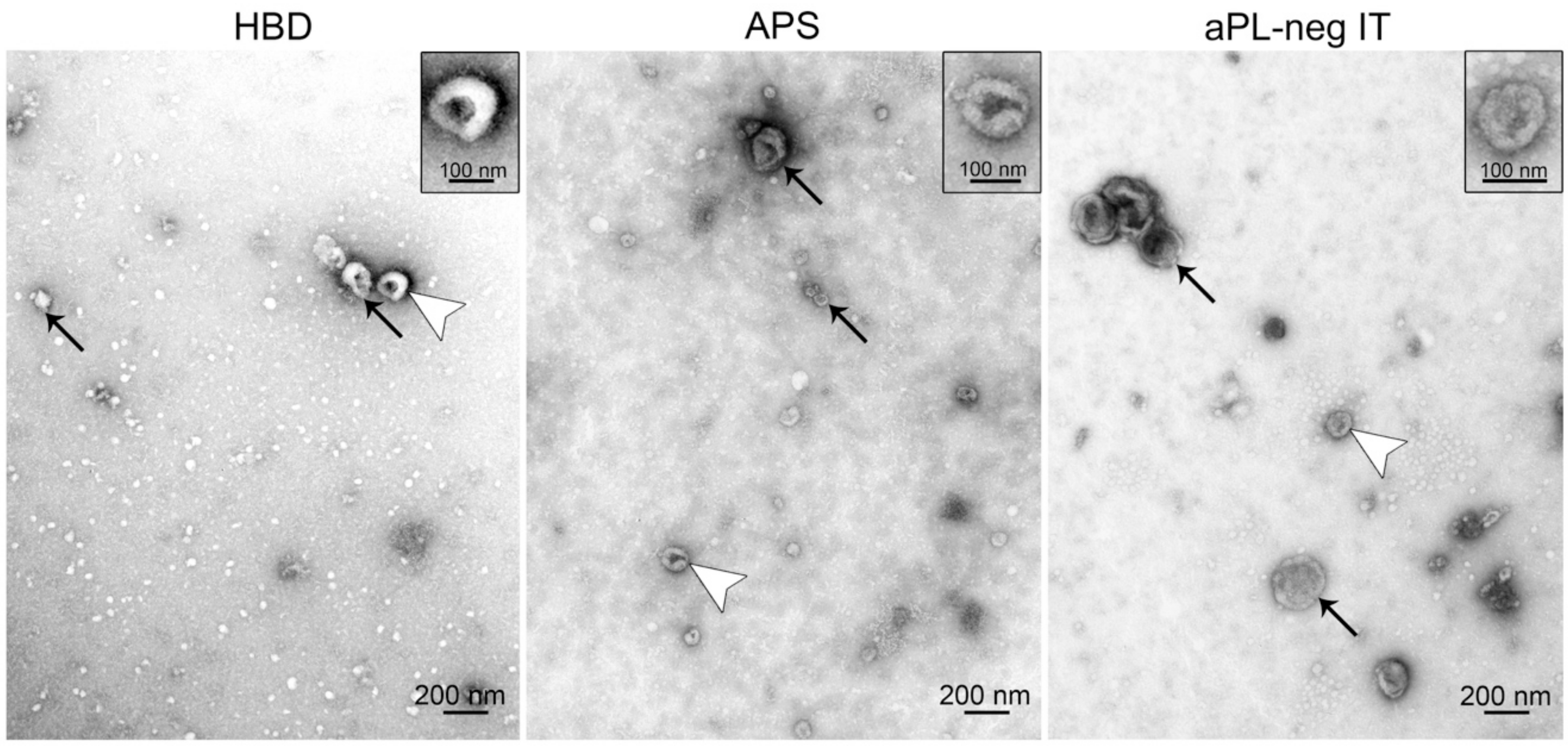
3.3. Transmission Electron Microscopy Confirms the Presence of sEVs in Plasma Isolates
3.4. MACSPlex Technology Enables Detection of Multiple Surface Protein Markers on sEVs Isolated from Human Plasma
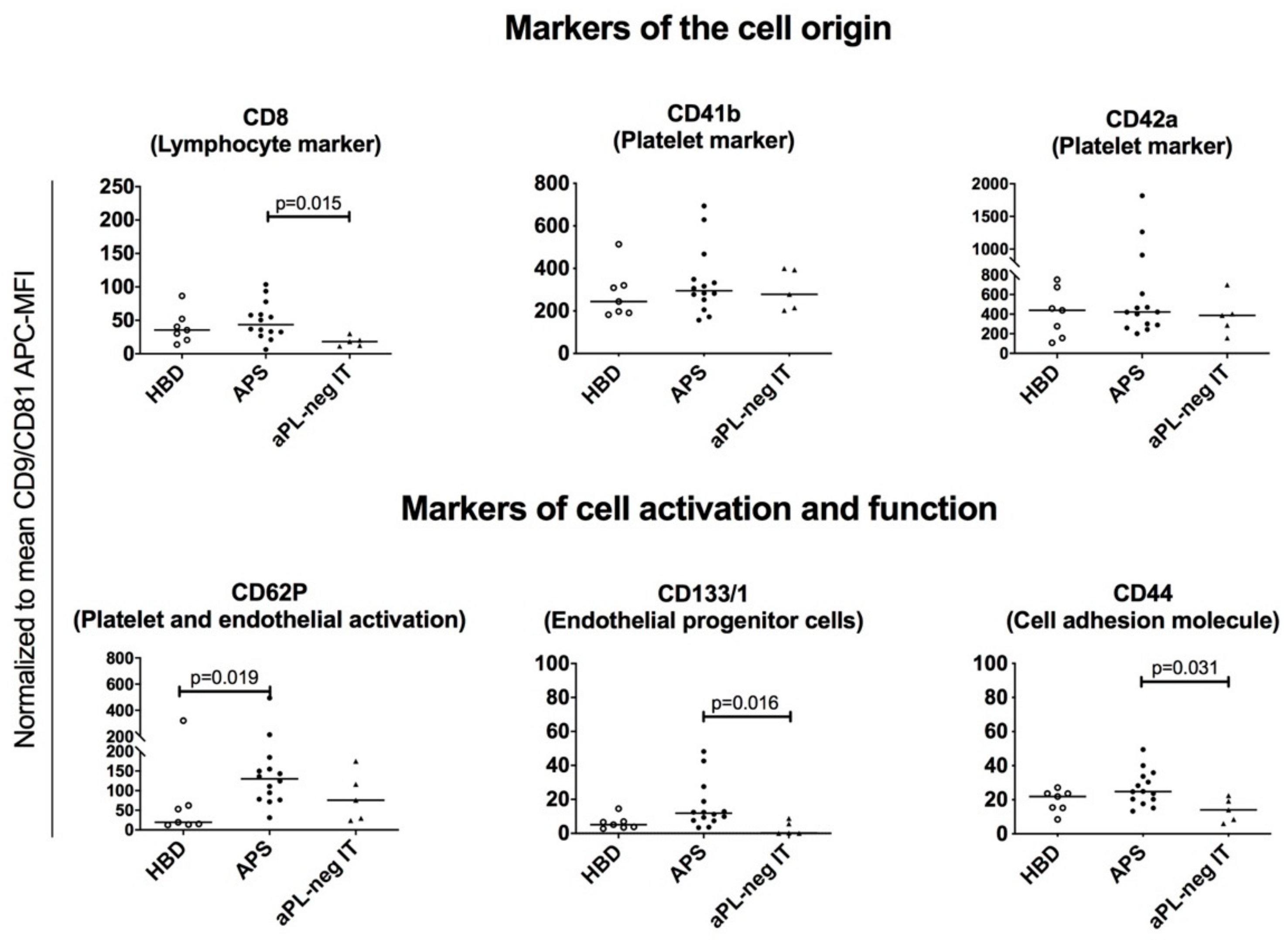
3.5. Platelet-, Lymphocyte-, Leukocyte- and Endothelial-Derived sEVs are Present in Plasma of Patients with APS, aPL-Negative Idiopathic Thrombosis and Healthy Blood Donors
3.6. sEVs from Plasma of Patients with APS Are Enriched for the Surface Expression of CD62P and CD133/1, Indicating Endothelial Activation/Damage and Platelet Activation in APS
3.7. EVs Isolated from Human Plasma Express Several Adhesion Molecules Involved in Cell-to-Cell and Cell-to-Matrix Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miyakis, S.; Lockshin, M.D.; Atsumi, T.; Branch, D.W.; Brey, R.L.; Cervera, R.; Derksen, R.H.; PG, D.E.G.; Koike, T.; Meroni, P.L.; et al. International consensus statement on an update of the classification criteria for definite antiphospholipid syndrome (APS). J. Thromb. Haemost. 2006, 4, 295–306. [Google Scholar] [CrossRef]
- Sammaritano, L.R. Antiphospholipid syndrome. Best Pract. Res. Clin. Rheumatol. 2019, 101463. [Google Scholar] [CrossRef]
- Giacomelli, R.; Afeltra, A.; Alunno, A.; Bartoloni-Bocci, E.; Berardicurti, O.; Bombardieri, M.; Bortoluzzi, A.; Caporali, R.; Caso, F.; Cervera, R.; et al. Guidelines for biomarkers in autoimmune rheumatic diseases - evidence based analysis. Autoimmun. Rev. 2019, 18, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, S.; Alluri, R.; McCrae, K.R. Extracellular Vesicles in the Antiphospholipid Syndrome. Semin. Thromb. Hemost. 2018, 44, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Veh. 2018, 7, 1535750. [Google Scholar] [CrossRef]
- O’Loghlen, A. Role for extracellular vesicles in the tumour microenvironment. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2018, 373, 20160488. [Google Scholar] [CrossRef]
- Combes, V.; Simon, A.C.; Grau, G.E.; Arnoux, D.; Camoin, L.; Sabatier, F.; Mutin, M.; Sanmarco, M.; Sampol, J.; Dignat-George, F. In vitro generation of endothelial microparticles and possible prothrombotic activity in patients with lupus anticoagulant. J. Clin. Investig. 1999, 104, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Dignat-George, F.; Camoin-Jau, L.; Sabatier, F.; Arnoux, D.; Anfosso, F.; Bardin, N.; Veit, V.; Combes, V.; Gentile, S.; Moal, V.; et al. Endothelial microparticles: A potential contribution to the thrombotic complications of the antiphospholipid syndrome. Thromb. Haemost. 2004, 91, 667–673. [Google Scholar] [CrossRef]
- Joseph, J.E.; Harrison, P.; Mackie, I.J.; Isenberg, D.A.; Machin, S.J. Increased circulating platelet-leucocyte complexes and platelet activation in patients with antiphospholipid syndrome, systemic lupus erythematosus and rheumatoid arthritis. Br. J. Haematol. 2001, 115, 451–459. [Google Scholar] [CrossRef]
- Jy, W.; Tiede, M.; Bidot, C.J.; Horstman, L.L.; Jimenez, J.J.; Chirinos, J.; Ahn, Y.S. Platelet activation rather than endothelial injury identifies risk of thrombosis in subjects positive for antiphospholipid antibodies. Thromb. Res. 2007, 121, 319–325. [Google Scholar] [CrossRef]
- Vikerfors, A.; Mobarrez, F.; Bremme, K.; Holmstrom, M.; Agren, A.; Eelde, A.; Bruzelius, M.; Antovic, A.; Wallen, H.; Svenungsson, E. Studies of microparticles in patients with the antiphospholipid syndrome (APS). Lupus 2012, 21, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Niccolai, E.; Squatrito, D.; Emmi, G.; Silvestri, E.; Emmi, L.; Ciucciarelli, L.; Ricci, F.; Manganaro, D.; Amedei, A.; Prisco, D. A new cytofluorimetric approach to evaluate the circulating microparticles in subjects with antiphospholipid antibodies. Thromb. Res. 2015, 136, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Breen, K.A.; Sanchez, K.; Kirkman, N.; Seed, P.T.; Parmar, K.; Moore, G.W.; Hunt, B.J. Endothelial and platelet microparticles in patients with antiphospholipid antibodies. Thromb. Res. 2015, 135, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, S.; Cockrell, E.; Espinola, R.; Hsi, L.; Fulton, S.; Khan, M.; Li, L.; Fonseca, F.; Kundu, S.; McCrae, K.R. Circulating microparticles in patients with antiphospholipid antibodies: Characterization and associations. Thromb. Res. 2015, 135, 102–108. [Google Scholar] [CrossRef]
- Amengual, O.; Forastiero, R.; Sugiura-Ogasawara, M.; Otomo, K.; Oku, K.; Favas, C.; Delgado Alves, J.; Zigon, P.; Ambrozic, A.; Tomsic, M.; et al. Evaluation of phosphatidylserine-dependent antiprothrombin antibody testing for the diagnosis of antiphospholipid syndrome: Results of an international multicentre study. Lupus 2017, 26, 266–276. [Google Scholar] [CrossRef]
- Bozic, B.; Kveder, T.; Stegnar, M.; Morosini-Berus, E.; Kos-Golja, M.; Peternel, P.; Rozman, B. Influence of degraded phosphatidylserine on binding of antiphospholipid antibodies. Int. Arch. Allergy Immunol. 1997, 112, 19–26. [Google Scholar] [CrossRef]
- Zigon, P.; Ambrozic, A.; Cucnik, S.; Kveder, T.; Rozman, B.; Bozic, B. Modified phosphatidylserine-dependent antiprothrombin ELISA enables identification of patients negative for other antiphospholipid antibodies and also detects low avidity antibodies. Clin. Chem. Lab. Med. 2011, 49, 1573. [Google Scholar]
- Sciascia, S.; Sanna, G.; Murru, V.; Roccatello, D.; Khamashta, M.A.; Bertolaccini, M.L. GAPSS: The Global Anti-Phospholipid Syndrome Score. Rheumatology (Oxf.) 2013, 52, 1397–1403. [Google Scholar] [CrossRef]
- Serrano-Pertierra, E.; Oliveira-Rodriguez, M. Characterization of Plasma-Derived Extracellular Vesicles Isolated by Different Methods: A Comparison Study. Bioengineering 2019, 6, 8. [Google Scholar] [CrossRef]
- Zahra, S.; Anderson, J.A.; Stirling, D.; Ludlam, C.A. Microparticles, malignancy and thrombosis. Br. J. Haematol. 2011, 152, 688–700. [Google Scholar] [CrossRef]
- Simak, J.; Gelderman, M.P.; Yu, H.; Wright, V.; Baird, A.E. Circulating endothelial microparticles in acute ischemic stroke: A link to severity, lesion volume and outcome. J. Thromb. Haemost. 2006, 4, 1296–1302. [Google Scholar] [CrossRef] [PubMed]
- Pierangeli, S.S.; Espinola, R.G.; Liu, X.; Harris, E.N. Thrombogenic effects of antiphospholipid antibodies are mediated by intercellular cell adhesion molecule-1, vascular cell adhesion molecule-1, and P-selectin. Circ. Res. 2001, 88, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Wu, H.; Liu, Y.; Zhao, M.; Li, D.; Lu, Q. Recent advances of exosomes in immune modulation and autoimmune diseases. Autoimmunity 2016, 49, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Niccolai, E.; Emmi, G.; Squatrito, D.; Silvestri, E.; Emmi, L.; Amedei, A.; Prisco, D. Microparticles: Bridging the Gap between Autoimmunity and Thrombosis. Semin. Thromb. Hemost. 2015, 41, 413–422. [Google Scholar] [CrossRef]
- Koliha, N. Analysis of the microRNA profile and Origin of Exosomes in Plasma of Melanoma Patients and Healthy Individuals. Ph.D. Thesis, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany, 6 October 2016. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HBD (n = 7) | APS (n = 14) | aPL-Neg IT (n = 5) | p Value | |
|---|---|---|---|---|
| Mean age (range) | 49 (28–67) | 47 (28–74) | 52 (38–73) | 0.322 |
| Sex (F:M) | 6:1 | 9:5 | 3:2 | 0.534 |
| Smoking | 2/7 (29 %) | 2/14 (14%) | 1/5 (20%) | 0.233 |
| Fasting | 1/7 (14%) | 5/14 (36%) | 2/5 (40%) | 0.641 |
| BMI (kg2) | 24.1 ± 6.8 | 26.7 ± 4.0 | 24.3 ± 4.8 | 0.324 |
| Systolic blood pressure (mmHg) | 114.5 ± 9.4 | 137.8 ± 17.9 | 115 ± 15.2 | 0.005 |
| Diastolic blood pressure (mmHg) | 75.2 ± 5.2 | 84.5 ± 7.7 | 75.6 ± 11.0 | 0.029 |
| Arterial thrombosis n (%) | 0 | 6/14 (43%) | 1/5 (20%) | 0.363 |
| Venous thrombosis n (%) | 0 | 9/14 (64%) | 4/5 (80%) | 0.516 |
| Microthrombosis n (%) | 0 | 2/14 (14%) | 0 | 0.372 |
| Obstetric complications n (%) | 0 | 3/14 (21%) | 1/5 (20%) | ns |
| Diabetes n (%) | 1/7 (14%) | 2/14 (14%) | 0 | 0.646 |
| Anticoagulant therapy n (%) | 0 | 12/14 (86%) | 2/5 (40%) | 0.005 |
| Anti-aggregation therapy n (%) | 0 | 4/14 (29%) | 1/5 (20%) | 0.471 |
| Antimalarics n (%) | 0 | 2/14 (14%) | 0 | 0.266 |
| Hormonal contraceptives n (%) | 2/7 (29%) | 4/14 (29%) | 2/5 (40%) | 0.624 |
| aCL (G/M/A) n (%) | 0 | 10/14 (71%) | 0 | 0.001 |
| IgG (<10 AU neg) | <5 | 20.9 ± 12.8 | <5 | 0.001 |
| IgM (<10 AU neg) | <5 | 10.4 ± 10.3 | <5 | 0.157 |
| IgA (<10 AU neg) | <5 | 4.4 ± 2.5 | <5 | 0.145 |
| anti-β2GPI (G/M/A) n (%) | 0 | 11/14 (79%) | 0 | <0.001 |
| IgG (<2 AU neg) | <2 | 10.9 ± 6.7 | <2 | 0.002 |
| IgM (<2 AU neg) | <2 | 2.21 ± 2.2 | <2 | 0.081 |
| IgA (<2 AU neg) | <2 | 1.9 ± 1.6 | <2 | 0.333 |
| aPS/PT (G/M/A) n (%) | 0 | 11/14 (79%) | 0 | 0.010 |
| IgG (<5 AU neg) | <5 | 41.5 ± 44.6 | <5 | 0.001 |
| IgM (<5 AU neg) | <5 | 21.4 ± 29.1 | <5 | 0.007 |
| IgA (<5 AU neg) | <5 | 6.7 ± 5.6 | <5 | 0.043 |
| LA n (%) | / | 10/14 (71%) | 0 | 0.006 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štok, U.; Blokar, E.; Lenassi, M.; Holcar, M.; Frank-Bertoncelj, M.; Erman, A.; Resnik, N.; Sodin-Šemrl, S.; Čučnik, S.; Perdan Pirkmajer, K.; et al. Characterization of Plasma-Derived Small Extracellular Vesicles Indicates Ongoing Endothelial and Platelet Activation in Patients with Thrombotic Antiphospholipid Syndrome. Cells 2020, 9, 1211. https://doi.org/10.3390/cells9051211
Štok U, Blokar E, Lenassi M, Holcar M, Frank-Bertoncelj M, Erman A, Resnik N, Sodin-Šemrl S, Čučnik S, Perdan Pirkmajer K, et al. Characterization of Plasma-Derived Small Extracellular Vesicles Indicates Ongoing Endothelial and Platelet Activation in Patients with Thrombotic Antiphospholipid Syndrome. Cells. 2020; 9(5):1211. https://doi.org/10.3390/cells9051211
Chicago/Turabian StyleŠtok, Ula, Elizabeta Blokar, Metka Lenassi, Marija Holcar, Mojca Frank-Bertoncelj, Andreja Erman, Nataša Resnik, Snežna Sodin-Šemrl, Saša Čučnik, Katja Perdan Pirkmajer, and et al. 2020. "Characterization of Plasma-Derived Small Extracellular Vesicles Indicates Ongoing Endothelial and Platelet Activation in Patients with Thrombotic Antiphospholipid Syndrome" Cells 9, no. 5: 1211. https://doi.org/10.3390/cells9051211
APA StyleŠtok, U., Blokar, E., Lenassi, M., Holcar, M., Frank-Bertoncelj, M., Erman, A., Resnik, N., Sodin-Šemrl, S., Čučnik, S., Perdan Pirkmajer, K., Ambrožič, A., & Žigon, P. (2020). Characterization of Plasma-Derived Small Extracellular Vesicles Indicates Ongoing Endothelial and Platelet Activation in Patients with Thrombotic Antiphospholipid Syndrome. Cells, 9(5), 1211. https://doi.org/10.3390/cells9051211