Functional Consequences of the Variable Stoichiometry of the Kv1.3-KCNE4 Complex

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
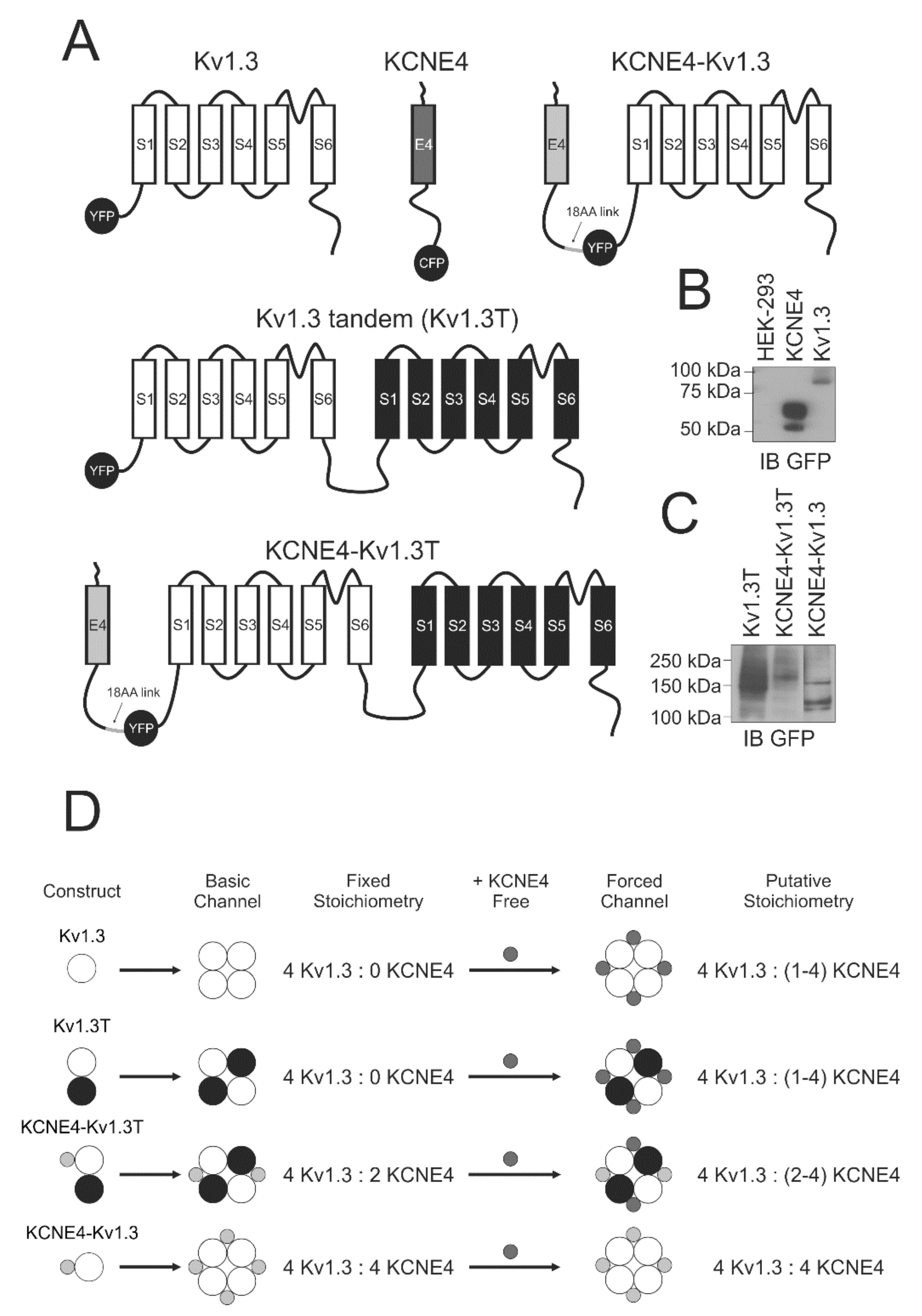
2.1. Expression Plasmids, Chimeric Channels and Site-Directed Mutagenesis
2.2. Cell Culture and Transient Transfection
2.3. Protein Extraction, Coimmunoprecipitation and Western Blotting
2.4. Confocal Microscopy and Image Analysis
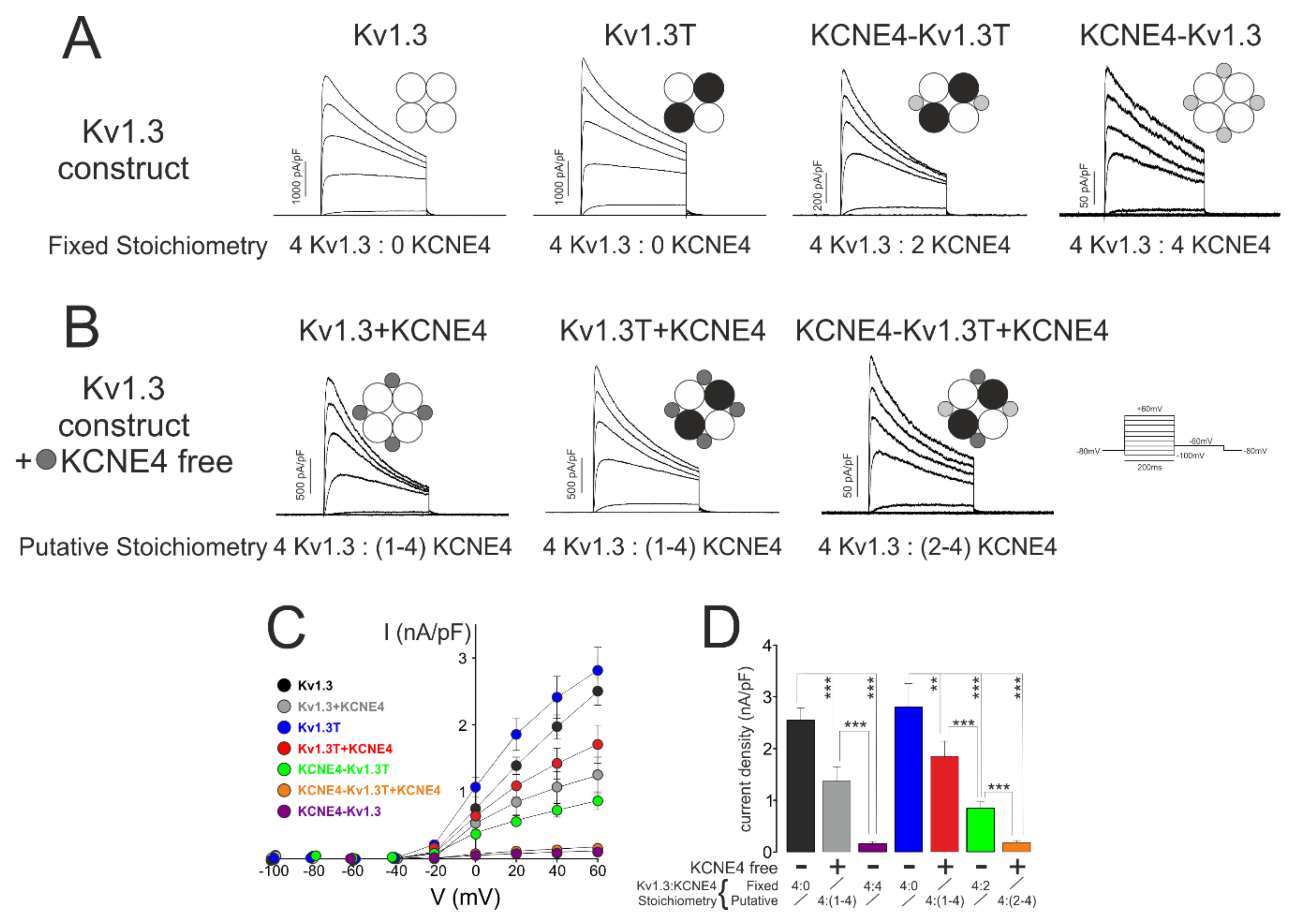
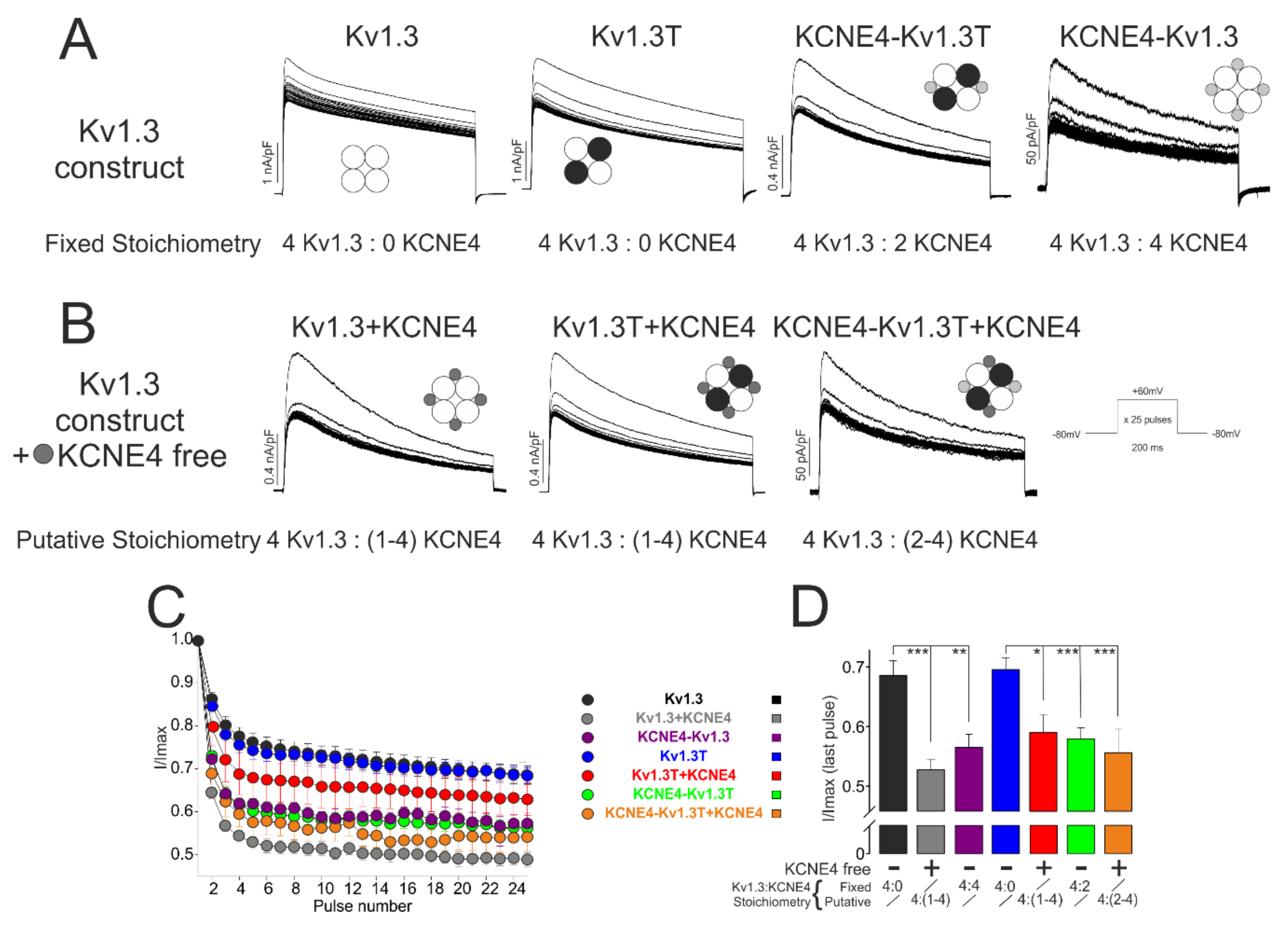
2.5. Electrophysiology
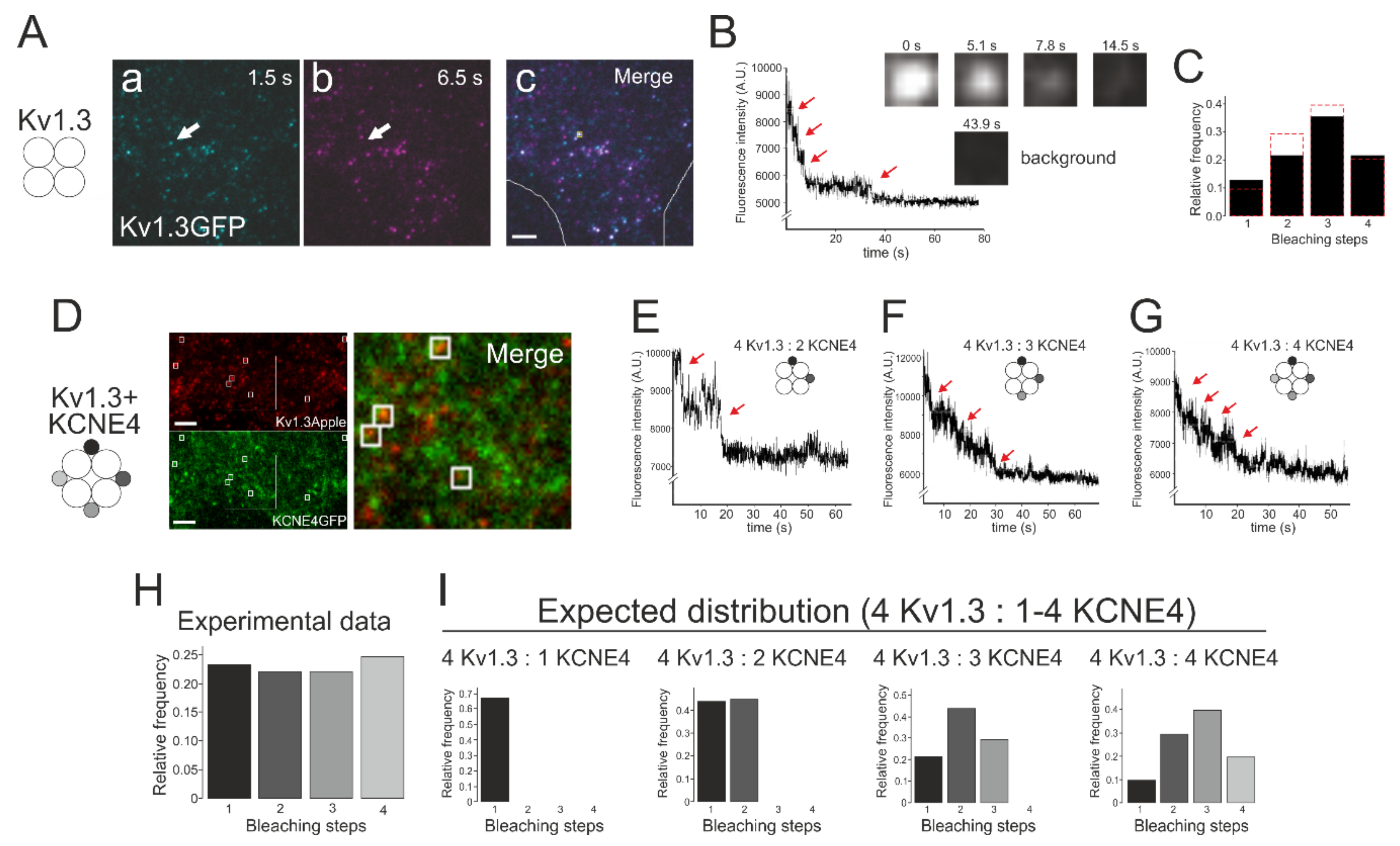
2.6. TIRF and Bleaching Steps of the Single Fluorescent Protein Complexes
2.7. Statistics
3. Results
3.1. KCNE4 Specifically Modulates the Traffic and Activity of Kv1.3
3.2. Kv1.3 Activity Depends on the Variations in Kv1.3-KCNE4 Stoichiometry
3.3. Bleaching Steps of KCNE4-GFP Suggested that as Many as Four KCNE4 Proteins Associate with a Kv1.3 Functional Tetrameric Channel
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chandy, K.G.; Norton, R.S. Peptide blockers of Kv1.3 channels in T cells as therapeutics for autoimmune disease. Curr. Opin. Chem. Biol. 2017, 38, 97–107. [Google Scholar] [CrossRef]
- Perez-Verdaguer, M.; Capera, J.; Serrano-Novillo, C.; Estadella, I.; Sastre, D.; Felipe, A. The voltage-gated potassium channel Kv1.3 is a promising multitherapeutic target against human pathologies. Expert Opin. Ther. Targets 2016, 20, 577–591. [Google Scholar] [CrossRef]
- Serrano-Albarras, A.; Cirera-Rocosa, S.; Sastre, D.; Estadella, I.; Felipe, A. Fighting rheumatoid arthritis: Kv1.3 as a therapeutic target. Biochem. Pharmacol. 2019, 165, 214–220. [Google Scholar] [CrossRef]
- Wulff, H.; Christophersen, P.; Colussi, P.; Chandy, K.G.; Yarov-Yarovoy, V. Antibodies and venom peptides: New modalities for ion channels. Nat. Rev. Drug Discov. 2019, 18, 339–357. [Google Scholar] [CrossRef]
- Vicente, R.; Escalada, A.; Villalonga, N.; Texido, L.; Roura-Ferrer, M.; Martin-Satue, M.; Lopez-Iglesias, C.; Soler, C.; Solsona, C.; Tamkun, M.M.; et al. Association of Kv1.5 and Kv1.3 contributes to the major voltage-dependent K+ channel in macrophages. J. Biol. Chem. 2006, 281, 37675–37685. [Google Scholar] [CrossRef]
- Villalonga, N.; Escalada, A.; Vicente, R.; Sanchez-Tillo, E.; Celada, A.; Solsona, C.; Felipe, A. Kv1.3/Kv1.5 heteromeric channels compromise pharmacological responses in macrophages. Biochem. Biophys. Res. Commun. 2007, 352, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Perez-Verdaguer, M.; Capera, J.; Martinez-Marmol, R.; Camps, M.; Comes, N.; Tamkun, M.M.; Felipe, A. Caveolin interaction governs Kv1.3 lipid raft targeting. Sci. Rep. 2016, 6, 22453. [Google Scholar] [CrossRef]
- Martinez-Marmol, R.; Styrczewska, K.; Perez-Verdaguer, M.; Vallejo-Gracia, A.; Comes, N.; Sorkin, A.; Felipe, A. Ubiquitination mediates Kv1.3 endocytosis as a mechanism for protein kinase C-dependent modulation. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, O.; Boratko, A.; Panyi, G.; Hajdu, P. The role of PSD-95 in the rearrangement of Kv1.3 channels to the immunological synapse. Pflug. Arch. Eur. J. Physiol. 2013, 465, 1341–1353. [Google Scholar] [CrossRef] [PubMed]
- McCormack, T.; McCormack, K.; Nadal, M.S.; Vieira, E.; Ozaita, A.; Rudy, B. The effects of Shaker beta-subunits on the human lymphocyte K+ channel Kv1.3. J. Biol. Chem. 1999, 274, 20123–20126. [Google Scholar] [CrossRef] [PubMed]
- Vicente, R.; Escalada, A.; Soler, C.; Grande, M.; Celada, A.; Tamkun, M.M.; Solsona, C.; Felipe, A. Pattern of Kv beta subunit expression in macrophages depends upon proliferation and the mode of activation. J. Immunol. 2005, 174, 4736–4744. [Google Scholar] [CrossRef] [PubMed]
- Sole, L.; Vallejo-Gracia, A.; Roig, S.R.; Serrano-Albarras, A.; Marruecos, L.; Manils, J.; Gomez, D.; Soler, C.; Felipe, A. KCNE gene expression is dependent on the proliferation and mode of activation of leukocytes. Channels 2013, 7, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Sole, L.; Felipe, A. Does a physiological role for KCNE subunits exist in the immune system? Commun. Integr. Biol. 2010, 3, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Sole, L.; Roura-Ferrer, M.; Perez-Verdaguer, M.; Oliveras, A.; Calvo, M.; Fernandez-Fernandez, J.M.; Felipe, A. KCNE4 suppresses Kv1.3 currents by modulating trafficking, surface expression and channel gating. J. Cell Sci. 2009, 122, 3738–3748. [Google Scholar] [CrossRef]
- Grunnet, M.; Rasmussen, H.B.; Hay-Schmidt, A.; Rosenstierne, M.; Klaerke, D.A.; Olesen, S.P.; Jespersen, T. KCNE4 is an inhibitory subunit to Kv1.1 and Kv1.3 potassium channels. Biophys. J. 2003, 85, 1525–1537. [Google Scholar] [CrossRef]
- Abbott, G.W. KCNE4 and KCNE5: K(+) channel regulation and cardiac arrhythmogenesis. Gene 2016, 593, 249–260. [Google Scholar] [CrossRef]
- Jepps, T.A.; Carr, G.; Lundegaard, P.R.; Olesen, S.P.; Greenwood, I.A. Fundamental role for the KCNE4 ancillary subunit in Kv7.4 regulation of arterial tone. J. Physiol. 2015, 593, 5325–5340. [Google Scholar] [CrossRef]
- Wrobel, E.; Tapken, D.; Seebohm, G. The KCNE Tango—How KCNE1 Interacts with Kv7.1. Front. Pharmacol. 2012, 3, 142. [Google Scholar] [CrossRef]
- Sole, L.; Roig, S.R.; Sastre, D.; Vallejo-Gracia, A.; Serrano-Albarras, A.; Ferrer-Montiel, A.; Fernandez-Ballester, G.; Tamkun, M.M.; Felipe, A. The calmodulin-binding tetraleucine motif of KCNE4 is responsible for association with Kv1.3. FASEB J. 2019, 33, 8263–8279. [Google Scholar] [CrossRef]
- Sole, L.; Roig, S.R.; Vallejo-Gracia, A.; Serrano-Albarras, A.; Martinez-Marmol, R.; Tamkun, M.M.; Felipe, A. The C-terminal domain of Kv1.3 regulates functional interactions with the KCNE4 subunit. J. Cell Sci. 2016, 129, 4265–4277. [Google Scholar] [CrossRef]
- Freidin, M.B.; Bragina, E.; Saltykova, I.V.; Deeva, E.V.; Ogorodova, L.M.; Puzyrev, V.P. Effect of additional disease (comorbidity) on association of allergic rhinitis with KCNE4 gene rs12621643 variant. Genetika 2013, 49, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Trevino, L.R.; Yang, W.; French, D.; Hunger, S.P.; Carroll, W.L.; Devidas, M.; Willman, C.; Neale, G.; Downing, J.; Raimondi, S.C.; et al. Germline genomic variants associated with childhood acute lymphoblastic leukemia. Nat. Genet. 2009, 41, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Nakajo, K.; Ulbrich, M.H.; Kubo, Y.; Isacoff, E.Y. Stoichiometry of the KCNQ1—KCNE1 ion channel complex. Proc. Natl. Acad. Sci. USA 2010, 107, 18862–18867. [Google Scholar] [CrossRef] [PubMed]
- Tamkun, M.M.; O’Connell, K.M.; Rolig, A.S. A cytoskeletal-based perimeter fence selectively corrals a sub-population of cell surface Kv2.1 channels. J. Cell Sci. 2007, 120, 2413–2423. [Google Scholar] [CrossRef]
- Ulbrich, M.H.; Isacoff, E.Y. Subunit counting in membrane-bound proteins. Nat. Methods 2007, 4, 319–321. [Google Scholar] [CrossRef]
- Ciampa, E.J.; Welch, R.C.; Vanoye, C.G.; George, A.L., Jr. KCNE4 juxtamembrane region is required for interaction with calmodulin and for functional suppression of KCNQ1. J. Biol. Chem. 2011, 286, 4141–4149. [Google Scholar] [CrossRef]
- Thompson, E.; Eldstrom, J.; Westhoff, M.; McAfee, D.; Fedida, D. The IKs Channel Response to cAMP Is Modulated by the KCNE1:KCNQ1 Stoichiometry. Biophys. J. 2018, 115, 1731–1740. [Google Scholar] [CrossRef]
- Wang, Y.; Eldstrom, J.; Fedida, D. The IKs ion channel activator, mefenamic acid, requires KCNE1 and modulates channel gating in a subunit-dependent manner. Mol. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Bendahhou, S.; Marionneau, C.; Haurogne, K.; Larroque, M.M.; Derand, R.; Szuts, V.; Escande, D.; Demolombe, S.; Barhanin, J. In vitro molecular interactions and distribution of KCNE family with KCNQ1 in the human heart. Cardiovasc. Res. 2005, 67, 529–538. [Google Scholar] [CrossRef]
- Roura-Ferrer, M.; Etxebarria, A.; Sole, L.; Oliveras, A.; Comes, N.; Villarroel, A.; Felipe, A. Functional implications of KCNE subunit expression for the Kv7.5 (KCNQ5) channel. Cell. Physiol. Biochem. 2009, 24, 325–334. [Google Scholar] [CrossRef]
- Tombola, F.; Ulbrich, M.H.; Isacoff, E.Y. The voltage-gated proton channel Hv1 has two pores, each controlled by one voltage sensor. Neuron 2008, 58, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ulbrich, M.H.; Li, M.H.; Buraei, Z.; Chen, X.Z.; Ong, A.C.; Tong, L.; Isacoff, E.Y.; Yang, J. Structural and molecular basis of the assembly of the TRPP2/PKD1 complex. Proc. Natl. Acad. Sci. USA 2009, 106, 11558–11563. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Sanguinetti, M.C. Molecular Basis of Cardiac Delayed Rectifier Potassium Channel Function and Pharmacology. Card. Electrophysiol. Clin. 2016, 8, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Manderfield, L.J.; George, A.L., Jr. KCNE4 can co-associate with the I(Ks) (KCNQ1-KCNE1) channel complex. FEBS J. 2008, 275, 1336–1349. [Google Scholar] [CrossRef]
- Chen, Y.H.; Xu, S.J.; Bendahhou, S.; Wang, X.L.; Wang, Y.; Xu, W.Y.; Jin, H.W.; Sun, H.; Su, X.Y.; Zhuang, Q.N.; et al. KCNQ1 gain-of-function mutation in familial atrial fibrillation. Science 2003, 299, 251–254. [Google Scholar] [CrossRef]
- Wang, K.; Terrenoire, C.; Sampson, K.J.; Iyer, V.; Osteen, J.D.; Lu, J.; Keller, G.; Kotton, D.N.; Kass, R.S. Biophysical properties of slow potassium channels in human embryonic stem cell derived cardiomyocytes implicate subunit stoichiometry. J. Physiol. 2011, 589, 6093–6104. [Google Scholar] [CrossRef]
- Wang, W.; Xia, J.; Kass, R.S. MinK-KvLQT1 fusion proteins, evidence for multiple stoichiometries of the assembled IsK channel. J. Biol. Chem. 1998, 273, 34069–34074. [Google Scholar] [CrossRef]
- Toyoda, F.; Ueyama, H.; Ding, W.G.; Matsuura, H. Modulation of functional properties of KCNQ1 channel by association of KCNE1 and KCNE2. Biochem. Biophys. Res. Commun. 2006, 344, 814–820. [Google Scholar] [CrossRef]
- Chen, H.; Kim, L.A.; Rajan, S.; Xu, S.; Goldstein, S.A. Charybdotoxin binding in the I(Ks) pore demonstrates two MinK subunits in each channel complex. Neuron 2003, 40, 15–23. [Google Scholar] [CrossRef]
- Morin, T.J.; Kobertz, W.R. Counting membrane-embedded KCNE beta-subunits in functioning K+ channel complexes. Proc. Natl. Acad. Sci. USA 2008, 105, 1478–1482. [Google Scholar] [CrossRef]
- Rasmusson, R.L.; Morales, M.J.; Wang, S.; Liu, S.; Campbell, D.L.; Brahmajothi, M.V.; Strauss, H.C. Inactivation of voltage-gated cardiac K+ channels. Circ. Res. 1998, 82, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Eghbali, M.; Olcese, R.; Zarei, M.M.; Toro, L.; Stefani, E. External pore collapse as an inactivation mechanism for Kv4.3 K+ channels. J. Membr. Biol. 2002, 188, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Cook, K.K.; Fadool, D.A. Two adaptor proteins differentially modulate the phosphorylation and biophysics of Kv1.3 ion channel by SRC kinase. J. Biol. Chem. 2002, 277, 13268–13280. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.S.; Zhang, M.; Jiang, M.; Wu, D.M.; Tseng, G.N. Probing the interaction between KCNE2 and KCNQ1 in their transmembrane regions. J. Membr. Biol. 2007, 216, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Melman, Y.F.; Domenech, A.; de la Luna, S.; McDonald, T.V. Structural determinants of KvLQT1 control by the KCNE family of proteins. J. Biol. Chem. 2001, 276, 6439–6444. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solé, L.; Sastre, D.; Colomer-Molera, M.; Vallejo-Gracia, A.; Roig, S.R.; Pérez-Verdaguer, M.; Lillo, P.; Tamkun, M.M.; Felipe, A. Functional Consequences of the Variable Stoichiometry of the Kv1.3-KCNE4 Complex. Cells 2020, 9, 1128. https://doi.org/10.3390/cells9051128
Solé L, Sastre D, Colomer-Molera M, Vallejo-Gracia A, Roig SR, Pérez-Verdaguer M, Lillo P, Tamkun MM, Felipe A. Functional Consequences of the Variable Stoichiometry of the Kv1.3-KCNE4 Complex. Cells. 2020; 9(5):1128. https://doi.org/10.3390/cells9051128
Chicago/Turabian StyleSolé, Laura, Daniel Sastre, Magalí Colomer-Molera, Albert Vallejo-Gracia, Sara R. Roig, Mireia Pérez-Verdaguer, Pilar Lillo, Michael M. Tamkun, and Antonio Felipe. 2020. "Functional Consequences of the Variable Stoichiometry of the Kv1.3-KCNE4 Complex" Cells 9, no. 5: 1128. https://doi.org/10.3390/cells9051128
APA StyleSolé, L., Sastre, D., Colomer-Molera, M., Vallejo-Gracia, A., Roig, S. R., Pérez-Verdaguer, M., Lillo, P., Tamkun, M. M., & Felipe, A. (2020). Functional Consequences of the Variable Stoichiometry of the Kv1.3-KCNE4 Complex. Cells, 9(5), 1128. https://doi.org/10.3390/cells9051128