Toxic-Metal-Induced Alteration in miRNA Expression Profile as a Proposed Mechanism for Disease Development

 ,
,  ,
,  ,
,  , , , , and
, , , , and 
Abstract
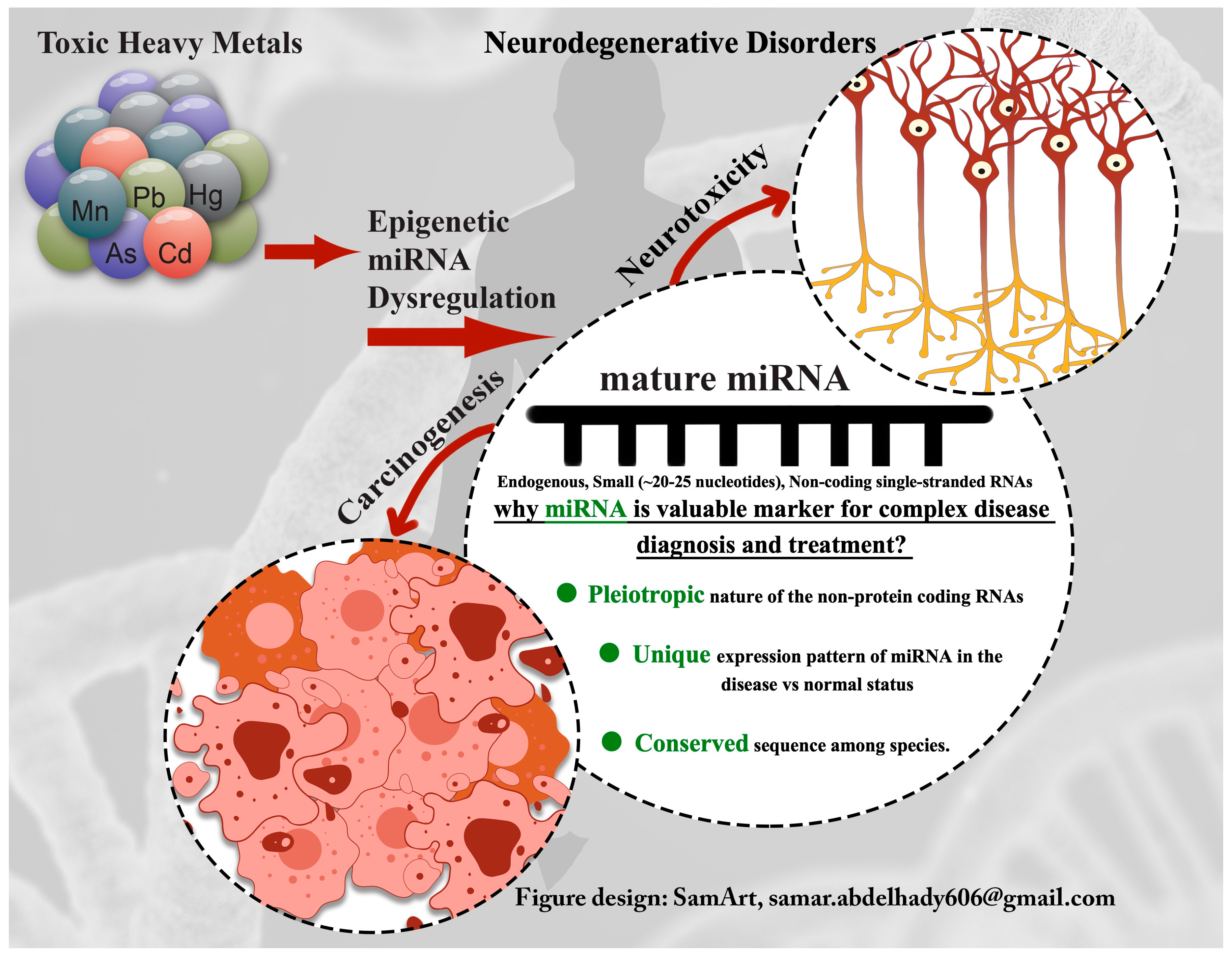
1. Introduction
1.1. Toxic Metals
1.2. Toxicoepigenomics and Environmentally-Associated Diseases
1.3. Epigenetics miRNA Signaling
2. Incorporating Toxicoepigenomic Data into the Risk Assessment
3. Determining Causal Toxic Metals and miRNA Signaling Cascade Associated with Disease Pathway
3.1. Cadmium-Associated Changes in miRNA Expression
3.2. Arsenic-Associated Changes in miRNA Expression
3.3. Lead-Associated Changes in miRNA Expression
3.4. Mercury-Associated Changes in miRNA Expression
3.5. Manganese-Associated Changes in miRNA Expression
4. Therapeutic Implications of miRNA in Human Diseases
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Wallace, D.R. Environmental Pesticides and Heavy Metals—Role in Breast Cancer. In Toxicity and Hazard of Agrochemicals; Larramendy, M.L., Soloneski, S., Eds.; InTech Open: London, UK, 2015; pp. 39–70. ISBN 978-953-51-2145-9. [Google Scholar]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. EXS 2012, 101, 133–164. [Google Scholar] [PubMed]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Gimeno-Garcia, E.; Andreu, V.; Boluda, R. Heavy Metals Incidence in the Application of Inorganic Fertilizers and Pesticides to Rice Farming Soils. Environ. Pollut. 1996, 92, 19–25. [Google Scholar] [CrossRef]
- Defarge, N.; Spiroux de Vendômois, J.; Séralini, G.E. Toxicity of formulants and heavy metals in glyphosate-based herbicides and other pesticides. Toxicol. Rep. 2018, 5, 156–163. [Google Scholar] [CrossRef]
- Shahid, M.; Khalid, S.; Abbas, G.; Shahid, N.; Nadeem, M.; Sabir, M.; Aslam, M.; Dumat, C. Heavy Metal Stress and Crop Productivity. In Crop Production and Global Environmental Issues; Hakeem, K., Ed.; Springe: Cham, Switzerland, 2015; pp. 1–25. [Google Scholar]
- Xin, J.; Huang, B.; Yang, Z.; Yuan, J.; Dai, H.; Qiu, Q. Responses of different water spinach cultivars and their hybrid to Cd, Pb and Cd-Pb exposures. J. Hazard. Mater. 2010, 175, 468–476. [Google Scholar] [CrossRef]
- Shen, C.; Huang, Y.Y.; He, C.T.; Zhou, Q.; Chen, J.X.; Tan, X.; Mubeen, S.; Yuan, J.G.; Yang, Z.Y. Comparative analysis of cadmium responsive microRNAs in roots of two Ipomoea aquatica Forsk. cultivars with different cadmium accumulation capacities. Plant Physiol. Biochem. 2017, 111, 329–339. [Google Scholar] [CrossRef]
- Gupta, O.P.; Sharma, P.; Gupta, R.K.; Sharma, I. MicroRNA mediated regulation of metal toxicity in plants: Present status and future perspectives. Plant Mol. Biol. 2014, 84, 1–18. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry Interaction Profile for: Arsenic, Cadmium, Chromium, and Lead. Available online: https://www.atsdr.cdc.gov/interactionprofiles/ip04.html (accessed on 7 April 2020).
- Nedelescu, M.; Baconi, D.; Neagoe, A.; Iordache, V.; Stan, M.; Constantinescu, P.; Ciobanu, A.M.; Vardavas, A.I.; Vinceti, M.; Tsatsakis, A.M. Environmental metal contamination and health impact assessment in two industrial regions of Romania. Sci. Total Environ. 2017, 580, 984–995. [Google Scholar] [CrossRef]
- Bjørklund, G.; Dadar, M.; Mutter, J.; Aaseth, J. The toxicology of mercury: Current research and emerging trends. Environ. Res. 2017, 159, 545–554. [Google Scholar] [CrossRef]
- Bjørklund, G.; Crisponi, G.; Nurchi, V.M.; Buha Djordjevic, A.; Aaseth, J. A Review on Coordination Properties of Thiol-Containing Chelating Agents towards Mercury, Cadmium, and Lead. Molecules 2019, 24, 3247. [Google Scholar] [CrossRef]
- Satarug, S. Dietary Cadmium Intake and Its Effects on Kidneys. Toxics 2018, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Casas, S.J.; Sordo, J. Lead Chemistry, Analytical aspects, Environmental Impact and Health Effects; Elsevier: Amsterdam, The Netherlands, 2006; ISBN 9780444529459. [Google Scholar]
- Renieri, E.A.; Safenkova, I.V.; Alegakis, A.; Slutskaya, E.S.; Kokaraki, V.; Kentouri, M.; Dzantiev, B.B.; Tsatsakis, A.M. Cadmium, lead and mercury in muscle tissue of gilthead seabream and seabass: Risk evaluation for consumers. Food Chem. Toxicol. 2019, 124, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Karaulov, A.V.; Renieri, E.A.; Smolyagin, A.I.; Mikhaylova, I.V.; Stadnikov, A.A.; Begun, D.N.; Tsarouhas, K.; Buha Djordjevic, A.; Hartung, T.; Tsatsakis, A. Long-term effects of chromium on morphological and immunological parameters of Wistar rats. Food Chem. Toxicol. 2019, 133, 110748. [Google Scholar] [CrossRef] [PubMed]
- Firestone, M.; Kavlock, R.; Zenick, H.; Kramer, M. The U.S. environmental protection agency strategic plan for evaluating the toxicity of chemicals. J. Toxicol. Environ. Health Part B Crit. Rev. 2010, 13, 139–162. [Google Scholar] [CrossRef]
- Ray, P.D.; Yosim, A.; Fry, R.C. Incorporating epigenetic data into the risk assessment process for the toxic metals arsenic, cadmium, chromium, lead, and mercury: Strategies and challenges. Front. Genet. 2014, 5, 1–26. [Google Scholar] [CrossRef]
- Sengupta, K.; Kolla, J.N.; Bagchi, D.; Bagchi, M. Applications of toxicogenomics in reproductive and developmental toxicology. In Reproductive and Developmental Toxicology; Academic Press: Cambridge, MA, USA, 2011; pp. 793–799. ISBN 978-0-12-804239-7. [Google Scholar]
- Ho, S.-M.; Johnson, A.; Tarapore, P.; Janakiram, V.; Zhang, X.; Leung, Y.-K. Environmental Epigenetics and Its Implication on Disease Risk and Health Outcomes. Ilar J. 2012, 53, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Zhang, S.; Hao, J.; Xie, F.; Hu, X.; Liu, C.; Tong, J.; Zhou, J.; Wu, J.; Shao, C. Downregulation of miR-132 by promoter methylation contributes to pancreatic cancer development. Carcinogenesis 2011, 32, 1183–1189. [Google Scholar] [CrossRef]
- Ramírez-Moya, J.; Wert-Lamas, L.; Riesco-Eizaguirre, G.; Santisteban, P. Impaired microRNA processing by DICER1 downregulation endows thyroid cancer with increased aggressiveness. Oncogene 2019, 38, 5486–5499. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Shriram, V.; Kumar, V.; Devarumath, R.M.; Khare, T.S.; Wani, S.H. MicroRNAs As Potential Targets for Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2016, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Qin, C.; Gao, J.; Liu, M.; Luo, X.; Zhang, W.; Liu, H.; Liao, X.; Shen, Y.; Mao, L.; et al. Genome-wide identification and analysis of drought-responsive genes and microRNAs in tobacco. Int. J. Mol. Sci. 2015, 16, 5714–5740. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.F.; Zhu, C. The role of microRNAs in copper and cadmium homeostasis. Biochem. Biophys. Res. Commun. 2009, 386, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.S.; Huang, S.Q.; Yang, Z.M. Bioinformatic identification and expression analysis of new microRNAs from Medicago truncatula. Biochem. Biophys. Res. Commun. 2008, 374, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Pani, A.; Mahapatra, R.K. Computational identification of microRNAs and their targets in Catharanthus roseus expressed sequence tags. Genom. Data 2013, 1, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.A.; Mesnage, R.; Tsatsakis, A.M.; Golokhvast, K.S.; Yang, S.H.; Antoniou, M.N.; Chung, G. Addressing concerns over the fate of DNA derived from genetically modified food in the human body: A review. Food Chem. Toxicol. 2019, 124, 423–430. [Google Scholar] [CrossRef]
- Hou, L.; Wang, D.; Baccarelli, A. Environmental chemicals and microRNAs. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2011, 714, 105–112. [Google Scholar] [CrossRef]
- Palanichamy, J.K.; Rao, D.S. miRNA dysregulation in cancer: Towards a mechanistic understanding. Front. Genet. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fromm, B.; Billipp, T.; Peck, L.E.; Johansen, M.; Tarver, J.E.; King, B.L.; Newcomb, J.M.; Sempere, L.F.; Flatmark, K.; Hovig, E.; et al. A Uniform System for the Annotation of Vertebrate microRNA Genes and the Evolution of the Human microRNAome. Annu. Rev. Genet. 2015, 49, 213–242. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Pulliero, A. The effects of environmental chemical carcinogens on the microRNA machinery. Int. J. Hyg. Environ. Health 2014, 217, 601–627. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fang, F.; Zhang, J.; Josson, S.; Clair, W.H.S.; Clair, D.K.S. Mir-17* suppresses tumorigenicity of prostate cancer by inhibiting mitochondrial antioxidant enzymes. PLoS ONE 2010, 5, e14356. [Google Scholar] [CrossRef]
- Yuan, W.; Yang, N.; Li, X. Advances in Understanding How Heavy Metal Pollution Triggers Gastric Cancer. BioMed Res. Int. 2016, 2016, 7825432. [Google Scholar] [CrossRef]
- Li, Q.; Kappil, M.A.; Li, A.; Dassanayake, P.S.; Darrah, T.H.; Friedman, A.E.; Friedman, M.; Lambertini, L.; Landrigan, P.; Stodgell, C.J.; et al. Exploring the associations between microRNA expression profiles and environmental pollutants in human placenta from the National Children’s Study (NCS). Epigenetics 2015, 10, 793–802. [Google Scholar] [CrossRef]
- Kong, A.P.S.; Xiao, K.; Choi, K.C.; Wang, G.; Chan, M.H.M.; Ho, C.S.; Chan, I.; Wong, C.K.; Chan, J.C.N.; Szeto, C.C. Associations between microRNA (miR-21, 126, 155 and 221), albuminuria and heavy metals in Hong Kong Chinese adolescents. Clin. Chim. Acta 2012, 413, 1053–1057. [Google Scholar] [CrossRef]
- He, B.S.; Qu, J.; Zhao, Q. Identifying and exploiting potential miRNA-disease associations with neighborhood regularized logistic matrix factorization. Front. Genet. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Schulte, C.; Molz, S.; Appelbaum, S.; Karakas, M.; Ojeda, F.; Lau, D.M.; Hartmann, T.; Lackner, K.J.; Westermann, D.; Schnabel, R.B.; et al. MiRNA-197 and miRNA-223 predict cardiovascular death in a cohort of patients with symptomatic coronary artery disease. PLoS ONE 2015, 10, e0145930. [Google Scholar] [CrossRef]
- Dardiotis, E.; Aloizou, A.M.; Siokas, V.; Patrinos, G.P.; Deretzi, G.; Mitsias, P.; Aschner, M.; Tsatsakis, A. The Role of MicroRNAs in Patients with Amyotrophic Lateral Sclerosis. J. Mol. Neurosci. 2018, 66, 617–628. [Google Scholar] [CrossRef]
- Jian, H.; Yang, B.O.; Zhang, A.; Ma, J.; Ding, Y.; Chen, Z.; Li, J.; Xu, X.; Liu, L. Genome-wide identification of microRNAs in response to cadmium stress in oilseed rape (Brassica napus L.) using high-throughput sequencing. Int. J. Mol. Sci. 2018, 19, 1431. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, Y.; Jiang, Z.; Wang, F.; Jiang, Q.; Sun, J.; Chen, Z.; Zhu, C. MicroRNA268 Overexpression Affects Rice Seedling Growth under Cadmium Stress. J. Agric. Food Chem. 2017, 65, 5860–5867. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Q.; Peng, J.; Qiu, C.X.; Yang, Z.M. Heavy metal-regulated new microRNAs from rice. J. Inorg. Biochem. 2009, 103, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Soto, A.B.; Sánchez, F.; Hernández, G. MicroRNAs as regulators in plant metal toxicity response. Front. Plant Sci. 2012, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Qiang, J.; Tao, Y.F.; He, J.; Xu, P.; Bao, J.W.; Sun, Y.L. miR-122 promotes hepatic antioxidant defense of genetically improved farmed tilapia (GIFT, Oreochromis niloticus) exposed to cadmium by directly targeting a metallothionein gene. Aquat. Toxicol. 2017, 182, 39–48. [Google Scholar] [CrossRef]
- Renieri, E.A.; Sfakianakis, D.G.; Alegakis, A.A.; Safenkova, I.V.; Buha, A.; Matović, V.; Tzardi, M.; Dzantiev, B.B.; Divanach, P.; Kentouri, M.; et al. Nonlinear responses to waterborne cadmium exposure in zebrafish. An in vivo study. Environ. Res. 2017, 157, 173–181. [Google Scholar] [CrossRef]
- Chen, S.; McKinney, G.J.; Nichols, K.M.; Colbourne, J.K.; Sepúlveda, M.S. Novel Cadmium Responsive MicroRNAs in Daphnia pulex. Environ. Sci. Technol. 2015, 49, 14605–14613. [Google Scholar] [CrossRef]
- Suwazono, Y.; Kido, T.; Nakagawa, H.; Nishijo, M.; Honda, R.; Kobayashi, E.; Dochi, M.; Nogawa, K. Biological half-life of cadmium in the urine of inhabitants after cessation of cadmium exposure. Biomarkers 2009, 14, 77–81. [Google Scholar] [CrossRef]
- Fransson, M.N.; Barregard, L.; Sallsten, G.; Akerstrom, M.; Johanson, G. Physiologically-based toxicokinetic model for cadmium using markov-chain monte carlo analysis of concentrations in blood, urine, and kidney cortex from living kidney donors. Toxicol. Sci. 2014, 141, 365–376. [Google Scholar] [CrossRef]
- Varga, B.; Zsolnai, B.; Paksy, K.; Náray, M.; Ungváry, G. Age dependent accumulation of cadmium in the human ovary. Reprod. Toxicol. 1993, 7, 225–228. [Google Scholar] [CrossRef]
- Wang, W.; Chen, J.; Luo, L.; Li, Y.; Liu, J.; Zhang, W. Effect of cadmium on kitl pre-mRNA alternative splicing in murine ovarian granulosa cells and its associated regulation by miRNAs. J. Appl. Toxicol. 2018, 38, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.A.; Martin, E.; Smeester, L.; Grace, M.R.; Boggess, K.; Fry, R.C. miRNAs as common regulators of the transforming growth factor (TGF)-β pathway in the preeclamptic placenta and cadmium-treated trophoblasts: Links between the environment, the epigenome and preeclampsia. Food Chem. Toxicol. 2016, 98, 50–57. [Google Scholar] [CrossRef]
- Fay, M.; Alt, L.; Ryba, D.; Salamah, R.; Peach, R.; Papaeliou, A.; Zawadzka, S.; Weiss, A.; Patel, N.; Rahman, A.; et al. Cadmium Nephrotoxicity Is Associated with Altered MicroRNA Expression in the Rat Renal Cortex. Toxics 2018, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, K.L.; Gerlach, C.V.; Craciun, F.L.; Ramachandran, K.; Bijol, V.; Kissick, H.T.; Vaidya, V.S. Application of small RNA sequencing to identify microRNAs in acute kidney injury and fibrosis. Toxicol. Appl. Pharmacol. 2016, 312, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pacheco, M.; Hidalgo-Miranda, A.; Romero-Córdoba, S.; Valverde, M.; Rojas, E. MRNA and miRNA expression patterns associated to pathways linked to metal mixture health effects. Gene 2014, 533, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Silantyev; Falzone; Libra; Gurina; Kardashova; Nikolouzakis; Nosyrev; Sutton; Panayioti; Tsatsakis Current and Future Trends on Diagnosis and Prognosis of Glioblastoma: From Molecular Biology to Proteomics. Cells 2019, 8, 863. [CrossRef]
- International Agency for Research on Cancer (IARC). IARC Cadmium and Cadmium Compounds, 100C ed.; WHO Press: Lyon, France, 2012; Volume 100C. [Google Scholar]
- Buha, A.; Wallace, D.; Matovic, V.; Schweitzer, A.; Oluic, B.; Micic, D.; Djordjevic, V. Cadmium exposure as a putative risk factor for the development of pancreatic cancer: Three different lines of evidence. BioMed Res. Int. 2017, 2017, 1981837. [Google Scholar] [CrossRef]
- Djordjevic, V.R.; Wallace, D.R.; Schweitzer, A.; Boricic, N.; Knezevic, D.; Matic, S.; Grubor, N.; Kerkez, M.; Radenkovic, D.; Bulat, Z.; et al. Environmental cadmium exposure and pancreatic cancer: Evidence from case control, animal and in vitro studies. Environ. Int. 2019, 128, 353–361. [Google Scholar] [CrossRef]
- Wallace, D.; Spandidos, D.; Tsatsakis, A.; Schweitzer, A.; Djordjevic, V.; Djordjevic, A. Potential interaction of cadmium chloride with pancreatic mitochondria: Implications for pancreatic cancer. Int. J. Mol. Med. 2019, 44, 145–156. [Google Scholar] [CrossRef]
- Buha, A.; Matovic, V.; Antonijevic, B.; Bulat, Z.; Curcic, M.; Renieri, E.A.E.A.; Tsatsakis, A.M.A.M.; Schweitzer, A.; Wallace, D. Overview of cadmium thyroid disrupting effects and mechanisms. Int. J. Mol. Sci. 2018, 19, 1501. [Google Scholar] [CrossRef]
- Humphries, B.; Wang, Z.; Yang, C. The role of microRNAs in metal carcinogen-induced cell malignant transformation and tumorigenesis. Food Chem. Toxicol. 2016, 98, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Ngalame, N.N.O.; Waalkes, M.P.; Tokar, E.J. Silencing KRAS Overexpression in Cadmium-Transformed Prostate Epithelial Cells Mitigates Malignant Phenotype. Chem. Res. Toxicol. 2016, 29, 1458–1467. [Google Scholar] [CrossRef] [PubMed]
- Urani, C.; Melchioretto, P.; Fabbri, M.; Bowe, G.; Maserati, E.; Gribaldo, L. Cadmium Impairs p53 Activity in HepG2 Cells. ISRN Toxicol. 2014, 2014, 976428. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zheng, C.; Shen, H.; Zhou, Z.; Lei, Y. MicroRNAs-mRNAs Expression Profile and Their Potential Role in Malignant Transformation of Human Bronchial Epithelial Cells Induced by Cadmium. BioMed Res. Int. 2015, 2015, 902025. [Google Scholar] [CrossRef]
- Marsit, C.J. Influence of environmental exposure on human epigenetic regulation. J. Exp. Biol. 2015, 218, 71–79. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Drinking-Water Quality; World Health Organization: Geneva, Switzerland, 2011; ISBN 9789241548151. [Google Scholar]
- Yu, L.; Luo, Y.; Liao, B.; Xie, L.; Chen, L.; Xiao, S.; Li, J.; Hu, S.; Shu, W. Comparative transcriptome analysis of transporters, phytohormone and lipid metabolism pathways in response to arsenic stress in rice (Oryza sativa). New Phytol. 2012, 195, 97–112. [Google Scholar] [CrossRef]
- Awasthi, S.; Chauhan, R.; Srivastava, S.; Tripathi, R.D. The Journey of Arsenic from Soil to Grain in Rice. Front. Plant Sci. 2017, 8, 1007. [Google Scholar] [CrossRef]
- Ghosh, S.; Singh, K.; Shaw, A.K.; Azahar, I.; Adhikari, S.; Ghosh, U.; Basu, U.; Roy, S.; Saha, S.; Sherpa, A.R.; et al. Insights into the miRNA-mediated response of maize leaf to arsenate stress. Environ. Exp. Bot. 2017, 137, 96–109. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, H. Molecular Identification and Analysis of Arsenite Stress-Responsive miRNAs in Rice. J. Agric. Food Chem. 2012, 60, 6524–6536. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K.; Suprasanna, P.; D’Souza, S.F. Identification and profiling of arsenic stress-induced microRNAs in Brassica juncea. J. Exp. Bot. 2013, 64, 303–315. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. Arsenic and arsenic compounds. In Overall Evaluations of Carcinogenicity: An Updating of IARC Monographs Volumes 1 to 42; World Health Organization: Lyon, France, 1987; pp. 68–70. [Google Scholar]
- Naujokas, M.F.; Anderson, B.; Ahsan, H.; Aposhian, H.V.; Graziano, J.H.; Thompson, C.; Suk, W.A. The Broad Scope of Health Effects from Chronic Arsenic Exposure: Update on a Worldwide Public Health Problem. Environ. Health Perspect. 2013, 121, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.V.; Pal, S.; Mohammed, A.; Farooqui, M.; Doescher, M.P.; Asch, A.S.; Yamada, H.Y. Biological effects and epidemiological consequences of arsenic exposure, and reagents that can ameliorate arsenic damage in vivo. Oncotarget 2017, 8, 57605–57621. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.S. Arsenic-induced oxidative stress and its reversibility. Free Radic. Biol. Med. 2011, 51, 257–281. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; McHale, C.M.; Skibola, C.F.; Smith, A.H.; Smith, M.T.; Zhang, L. An Emerging Role for Epigenetic Dysregulation in Arsenic Toxicity and Carcinogenesis. Environ. Health Perspect. 2011, 119, 11–19. [Google Scholar] [CrossRef]
- Bailey, K.A.; Fry, R.C. Arsenic-Induced Changes to the Epigenome. In Toxicology and Epigenetics; John Wiley & Sons, Ltd.: Chichester, UK, 2012; pp. 149–190. [Google Scholar]
- Cardoso, A.P.F.; Al-Eryani, L.; States, J.C. Arsenic-Induced Carcinogenesis: The Impact of miRNA Dysregulation. Toxicol. Sci. 2018, 165, 284–290. [Google Scholar] [CrossRef]
- Bjørklund, G.; Aaseth, J.; Chirumbolo, S.; Urbina, M.A.; Uddin, R. Effects of arsenic toxicity beyond epigenetic modifications. Environ. Geochem. Health 2018, 44, 955–965. [Google Scholar] [CrossRef]
- Fang, X.; Sun, R.; Hu, Y.; Wang, H.; Guo, Y.; Yang, B.; Pi, J.; Xu, Y. miRNA-182-5p, via HIF2α, contributes to arsenic carcinogenesis: Evidence from human renal epithelial cells. Metallomics 2018, 10, 1607–1617. [Google Scholar] [CrossRef]
- Michailidi, C.; Hayashi, M.; Datta, S.; Sen, T.; Zenner, K.; Oladeru, O.; Brait, M.; Izumchenko, E.; Baras, A.; VandenBussche, C.; et al. Involvement of epigenetics and EMT-related miRNA in arsenic-induced neoplastic transformation and their potential clinical use. Cancer Prev. Res. 2015, 8, 208–221. [Google Scholar] [CrossRef]
- Gao, J.; Wang, G.; Wu, J.; Zuo, Y.; Zhang, J.; Jin, X. Skp2 Expression Is Inhibited by Arsenic Trioxide through the Upregulation of miRNA-330-5p in Pancreatic Cancer Cells. Mol. Ther. Oncolytics 2019, 12, 214–223. [Google Scholar] [CrossRef]
- Ghaffari, S.H.; Bashash, D.; Dizaji, M.Z.; Ghavamzadeh, A.; Alimoghaddam, K. Alteration in miRNA gene expression pattern in acute promyelocytic leukemia cell induced by arsenic trioxide: A possible mechanism to explain arsenic multi-target action. Tumor Biol. 2012, 33, 157–172. [Google Scholar] [CrossRef]
- Nurchi; Djordjevic; Crisponi; Alexander; Bjørklund; Aaseth Arsenic Toxicity: Molecular Targets and Therapeutic Agents. Biomolecules 2020, 10, 235. [CrossRef]
- Abernathy, C.O.; Thomas, D.J.; Calderon, R.L. Health Effects and Risk Assessment of Arsenic. J. Nutr. 2003, 133, 1536S–1538S. [Google Scholar] [CrossRef]
- Tyler, C.R.; Allan, A.M. The Effects of Arsenic Exposure on Neurological and Cognitive Dysfunction in Human and Rodent Studies: A Review. Curr. Environ. Health Rep. 2014, 1, 132–147. [Google Scholar] [CrossRef]
- Ruíz-Vera, T.; Ochoa-Martínez, Á.C.; Zarazúa, S.; Carrizales-Yáñez, L.; Pérez-Maldonado, I.N. Circulating miRNA-126, -145 and -155 levels in Mexican women exposed to inorganic arsenic via drinking water. Environ. Toxicol. Pharmacol. 2019, 67, 79–86. [Google Scholar] [CrossRef]
- Beck, R.; Bommarito, P.; Douillet, C.; Kanke, M.; Del Razo, L.M.; García-Vargas, G.; Fry, R.C.; Sethupathy, P.; Stýblo, M. Circulating miRNAs Associated with Arsenic Exposure. Environ. Sci. Technol. 2018, 52, 14487–14495. [Google Scholar] [CrossRef]
- Rager, J.E.; Bailey, K.A.; Smeester, L.; Miller, S.K.; Parker, J.S.; Laine, J.E.; Drobná, Z.; Currier, J.; Douillet, C.; Olshan, A.F.; et al. Prenatal arsenic exposure and the epigenome: Altered microRNAs associated with innate and adaptive immune signaling in newborn cord blood. Environ. Mol. Mutagenesis 2014, 55, 196–208. [Google Scholar] [CrossRef]
- Martin, E.M.; Stýblo, M.; Fry, R.C. Genetic and epigenetic mechanisms underlying arsenic-associated diabetes mellitus: A perspective of the current evidence. Epigenomics 2017, 9, 701–710. [Google Scholar] [CrossRef]
- Banerjee, N.; Bandyopadhyay, A.K.; Dutta, S.; Das, J.K.; Roy Chowdhury, T.; Bandyopadhyay, A.; Giri, A.K. Increased microRNA 21 expression contributes to arsenic induced skin lesions, skin cancers and respiratory distress in chronically exposed individuals. Toxicology 2017, 378, 10–16. [Google Scholar] [CrossRef]
- Liu, J.Q.; Niu, Q.; Hu, J.H.; Li, Y.; Wang, H.X.; Xu, S.Z.; Ding, Y.S.; Li, S.G.; Ma, R.L. The Bidirectional Effects of Arsenic on miRNA-21: A Systematic Review and Meta-analysis. Biomed. Environ. Sci. 2018, 31, 654–666. [Google Scholar]
- Al-Eryani, L.; Jenkins, S.F.; States, V.A.; Pan, J.; Malone, J.C.; Rai, S.N.; Galandiuk, S.; Giri, A.K.; States, J.C. miRNA expression profiles of premalignant and malignant arsenic-induced skin lesions. PLoS ONE 2018, 13, e0202579. [Google Scholar] [CrossRef]
- Schaefer, A.; O’Carroll, D.; Chan, L.T.; Hillman, D.; Sugimori, M.; Llinas, R.; Greengard, P. Cerebellar neurodegeneration in the absence of microRNAs. J. Exp. Med. 2007, 204, 1553–1558. [Google Scholar] [CrossRef]
- Lukiw, W.J. Micro-RNA speciation in fetal, adult and Alzheimer’s disease hippocampus. NeuroReport 2007, 18, 297–300. [Google Scholar] [CrossRef]
- Lukiw, J.W.; Pogue, I.A. Induction of specific micro RNA (miRNA) species by ROS- generating metal sulfates in primary human brain cells. J. Inorg. Biochem. 2007, 101, 1265–1269. [Google Scholar] [CrossRef]
- Xu, M.; Yu, Z.; Hu, F.; Zhang, H.; Zhong, L.; Han, L.; An, Y.; Zhu, B.; Zhang, H. Identification of differential plasma miRNA profiles in Chinese workers with occupational lead exposure. Biosci. Rep. 2017, 37, BSR20171111. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Z.; Deng, M.; Liu, R.; Niu, S.; Fan, G. Identification and functional analysis of MicroRNAs and their targets in Platanus acerifolia under lead (Pb) stress. Int. J. Mol. Sci. 2015, 16, 7098–7111. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Shen, H.; Zhu, X.; Zhai, L.; Xu, L.; Wang, R.; Gong, Y.; Limera, C.; Liu, L. Identification of Radish (Raphanus sativus L.) miRNAs and Their Target Genes to Explore miRNA-Mediated Regulatory Networks in Lead (Pb) Stress Responses by High-Throughput Sequencing and Degradome Analysis. Plant Mol. Biol. Report. 2015, 33, 358–376. [Google Scholar]
- He, Q.; Zhu, S.; Zhang, B. MicroRNA-target gene responses to lead-induced stress in cotton (Gossypium hirsutum L.). Funct. Integr. Genom. 2014, 14, 507–515. [Google Scholar] [CrossRef]
- Masoud, A.M.; Bihaqi, S.W.; Machan, J.T.; Zawia, N.H.; Renehan, W.E. Early-life exposure to lead (Pb) alters the expression of microrna that target proteins associated with Alzheimer’s disease. J. Alzheimer’s Dis. 2016, 51, 1257–1264. [Google Scholar] [CrossRef]
- An, J.; Cai, T.; Che, H.; Yu, T.; Cao, Z.; Liu, X.; Zhao, F.; Jing, J.; Shen, X.; Liu, M.; et al. The changes of miRNA expression in rat hippocampus following chronic lead exposure. Toxicol. Lett. 2014, 229, 158–166. [Google Scholar] [CrossRef]
- Su, P.; Zhao, F.; Cao, Z.; Zhang, J.; Aschner, M.; Luo, W. Mir-203-mediated tricellulin mediates lead-induced in vitro loss of blood-cerebrospinal fluid barrier (BCB) function. Toxicol. In Vitro 2015, 29, 1185–1194. [Google Scholar] [CrossRef]
- Siddeek, B.; Inoubli, L.; Lakhdari, N.; Rachel, P.B.; Fussell, K.C.; Schneider, S.; Mauduit, C.; Benahmed, M. MicroRNAs as potential biomarkers in diseases and toxicology. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2014, 764–765, 46–57. [Google Scholar] [CrossRef]
- Feng, Y.-H.; Tsao, C.-J. Emerging role of microRNA-21 in cancer. Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef]
- Bollati, V.; Marinelli, B.; Apostoli, P.; Bonzini, M.; Nordio, F.; Hoxha, M.; Pegoraro, V.; Motta, V.; Tarantini, L.; Cantone, L.; et al. Exposure to metal-rich particulate matter modifies the expression of candidate MicroRNAs in peripheral blood leukocytes. Environ. Health Perspect. 2010, 118, 763–768. [Google Scholar] [CrossRef]
- Guo, X.; Yang, Q.; Zhang, W.; Chen, Y.; Ren, J.; Gao, A. Associations of blood levels of trace elements and heavy metals with metabolic syndrome in Chinese male adults with microRNA as mediators involved. Environ. Pollut. 2019, 66–73. [Google Scholar] [CrossRef]
- Ding, E.; Guo, J.; Bai, Y.; Zhang, H.; Liu, X.; Cai, W.; Zhong, L.; Zhu, B. MiR-92a and miR-486 are potential diagnostic biomarkers for mercury poisoning and jointly sustain NF-κB activity in mercury toxicity. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Faro, L.R.F.; Do Nascimento, J.L.M.; Alfonso, M.; Durán, R. In vivo effects of inorganic mercury (HgCl2) on striatal dopaminergic system. Ecotoxicol. Environ. Saf. 2001, 48, 263–267. [Google Scholar] [CrossRef]
- Belyaeva, E.A.; Korotkov, S.M.; Saris, N.E. In vitro modulation of heavy metal-induced rat liver mitochondria dysfunction: A comparison of copper and mercury with cadmium. J. Trace Elem. Med. Biol. 2011, 25, S63–S73. [Google Scholar] [CrossRef]
- Belyaeva, E.A.; Sokolova, T.V.; Emelyanova, L.V.; Zakharova, I.O. Mitochondrial Electron Transport Chain in Heavy Metal-Induced Neurotoxicity: Effects of Cadmium, Mercury, and Copper. Sci. World J. 2012, 2012, 1–14. [Google Scholar] [CrossRef]
- Wallace, D.R.; Lienemann, E.; Hood, A.N. Clinical aspects of mercury neurotoxicity. In Clinical Neurotoxicology: Syndromes, Substances, Environments; Dobbs, M., Ed.; Saunders-Elsevier: Philadelphia, PA, USA, 2009; pp. 251–258. ISBN 9780323052603. [Google Scholar]
- Wallace, D.R. Current State of Developmental Neurotoxicology Research. Toxics 2015, 3, 370–372. [Google Scholar] [CrossRef]
- Belyaeva, E.A.; Dymkowska, D.; Wieckowski, M.R.; Wojtczak, L. Mitochondria as an important target in heavy metal toxicity in rat hepatoma AS-30D cells. Toxicol. Appl. Pharmacol. 2008, 231, 34–42. [Google Scholar] [CrossRef]
- Stohs, S.J.J.; Bagchi, D. Oxidative mechanisms in the toxicity of metal ions. Free Radic. Biol. Med. 1995, 18, 321–336. [Google Scholar] [CrossRef]
- Cao, C.; Young, M.; Wong, J.; Mahran, L.; Eldefrawi, M. Putative cocaine receptor in striatum is a glycoprotein with active thiol function. Membr. Biochem. 1989, 8, 207–220. [Google Scholar] [CrossRef]
- Franco, J.L.; Posser, T.; Dunkley, P.R.; Dickson, P.W.; Mattos, J.J.; Martins, R.; Bainy, A.C.D.; Marques, M.R.; Dafre, A.L.; Farina, M. Methylmercury neurotoxicity is associated with inhibition of the antioxidant enzyme glutathione peroxidase. Free Radic. Biol. Med. 2009, 47, 449–457. [Google Scholar] [CrossRef]
- Franco, R.; Sanchez-Olea, R.; Reyes-Reyes, E.M.; Panayiotidis, M.I. Environmental toxicity, oxidative stress and apoptosis: Menage e Trois. Mutat. Res. 2009, 674, 3–22. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Zeng, H.Q.; Liu, Z.P.; Yang, Z.M. Genome-wide identification of Medicago truncatula microRNAs and their targets reveals their differential regulation by heavy metal. Plant Cell Environ. 2012, 35, 86–99. [Google Scholar] [CrossRef]
- Ding, E.; Zhao, Q.; Bai, Y.; Xu, M.; Pan, L.; Liu, Q.; Wang, B.; Song, X.; Wang, J.; Chen, L.; et al. Plasma microRNAs expression profile in female workers occupationally exposed to mercury. J. Thorac. Dis. 2016, 8, 833–841. [Google Scholar] [CrossRef][Green Version]
- Sriram, K.; Lin, G.X.; Jefferson, A.M.; Roberts, J.R.; Chapman, R.S.; Chen, B.T.; Soukup, J.M.; Ghio, A.J.; Antonini, J.M. Dopaminergic neurotoxicity following pulmonary exposure to manganese-containing welding fumes. Arch. Toxicol. 2010, 84, 521–540. [Google Scholar] [CrossRef]
- Brown, S.; Taylor, N.L. Could mitochondrial dysfunction play a role in manganese toxicity? Environ. Toxicol. Pharmacol. 1999, 7, 49–57. [Google Scholar] [CrossRef]
- Martin, S.; Van Veen, S.; Holemans, T.; Demirsoy, S.; Van Den Haute, C.; Baekelandt, V.; Agostinis, P.; Eggermont, J.; Vangheluwe, P. Protection against Mitochondrial and Metal Toxicity Depends on Functional Lipid Binding Sites in ATP13A2. Parkinson’s Dis. 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Valdes-Lopez, O.; Yang, S.S.; Aparicio-Fabre, R.; Graham, P.H.; Reyes, J.L.; Vance, C.P.; Hernandez, G. MicroRNA expression profile in common bean (Phaseolus vulgaris) under nutrient deficiency stresses and manganese toxicity. New Phytol. 2010, 187, 805–818. [Google Scholar] [CrossRef]
- Grogg, M.W.; Braydich-Stolle, L.K.; Maurer-Gardner, E.I.; Hill, N.T.; Sakaram, S.; Kadakia, M.P.; Hussain, S.M. Modulation of miRNA-155 alters manganese nanoparticle-induced inflammatory response. Toxicol. Res. 2016, 5, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.M.Y.; Chan, B.K.K.; Park, J.S.; Hill, K.J.; Aitken, J.B.; Cottle, L.; Farghaian, H.; Cole, A.R.; Lay, P.A.; Sue, C.M.; et al. Parkinson’s disease-linked human PARK9/ATP13A2 maintains zinc homeostasis and promotes α-Synuclein externalization via exosomes. Hum. Mol. Genet. 2014, 23, 2816–2833. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Xie, X.; Lv, L.; Huang, Y.; Xia, X.; Chen, X.; Zhang, L. Comprehensive investigation of aberrant microRNAs expression in cells culture model of MnCl2-induced neurodegenerative disease. Biochem. Biophys. Res. Commun. 2017, 486, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Tarale, P.; Daiwile, A.P.; Sivanesan, S.; Stöger, R.; Bafana, A.; Naoghare, P.K.; Parmar, D.; Chakrabarti, T.; Krishnamurthi, K. Manganese exposure: Linking down-regulation of miRNA-7 and miRNA-433 with α-synuclein overexpression and risk of idiopathic Parkinson’s disease. Toxicol. In Vitro 2018, 46, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Harischandra, D.S.; Ghaisas, S.; Rokad, D.; Zamanian, M.; Jin, H.; Anantharam, V.; Kimber, M.; Kanthasamy, A.; Kanthasamy, A.G. Environmental neurotoxicant manganese regulates exosome-mediated extracellular miRNAs in cell culture model of Parkinson’s disease: Relevance to α-synuclein misfolding in metal neurotoxicity. NeuroToxicology 2018, 64, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Wei, Z.; Shaikh, Z.A. Requirement of ERα and basal activities of EGFR and Src kinase in Cd-induced activation of MAPK/ERK pathway in human breast cancer MCF-7 cells. Toxicol. Appl. Pharmacol. 2015, 287, 26–34. [Google Scholar] [CrossRef]
- Koedrith, P.; Kim, H.L.; Weon, J.I.; Seo, Y.R. Toxicogenomic approaches for understanding molecular mechanisms of heavy metal mutagenicity and carcinogenicity. Int. J. Hyg. Environ. Health 2013, 216, 587–598. [Google Scholar] [CrossRef]
- Caffo, M.; Caruso, G.; Fata, G.L.; Barresi, V.; Visalli, M.; Venza, M.; Venza, I. Heavy Metals and Epigenetic Alterations in Brain Tumors. Curr. Genom. 2014, 15, 457–463. [Google Scholar] [CrossRef]
- Fiati Kenston, S.S.; Su, H.; Li, Z.; Kong, L.; Wang, Y.; Song, X.; Gu, Y.; Barber, T.; Aldinger, J.; Hua, Q.; et al. The systemic toxicity of heavy metal mixtures in rats. Toxicol. Res. 2018, 7, 396–407. [Google Scholar] [CrossRef]
- Walayat, A.; Yang, M.; Xiao, D. Therapeutic Implication of miRNA in Human Disease. In Antisense Therapy; Sharad, S., Kapur, S., Eds.; IntechOpen: London, UK, 2019; ISBN 978-1-78984-532-7. [Google Scholar]
- Garzon, R.; Marcucci, G.; Croce, C.M. Targeting microRNAs in cancer: Rationale, strategies and challenges. Nat. Rev. Drug Discov. 2010, 9, 775–789. [Google Scholar] [CrossRef]
- Beg, M.S.; Brenner, A.J.; Sachdev, J.; Borad, M.; Kang, Y.-K.; Stoudemire, J.; Smith, S.; Bader, A.G.; Kim, S.; Hong, D.S. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Acunzo, M.; Romano, G.; Palmieri, D.; Lagana, A.; Garofalo, M.; Balatti, V.; Drusco, A.; Chiariello, M.; Nana-Sinkam, P.; Croce, C.M. Cross-talk between MET and EGFR in non-small cell lung cancer involves miR-27a and Sprouty2. Proc. Natl. Acad. Sci. USA 2013, 110, 8573–8578. [Google Scholar] [CrossRef] [PubMed]
- Van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.J.; Bailey, D.L.; et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: A first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef]
- Noguchi, S.; Iwasaki, J.; Kumazaki, M.; Mori, T.; Maruo, K.; Sakai, H.; Yamada, N.; Shimada, K.; Naoe, T.; Kitade, Y.; et al. Chemically Modified Synthetic microRNA-205 Inhibits the Growth of Melanoma Cells In Vitro and In Vivo. Mol. Ther. 2013, 21, 1204–1211. [Google Scholar] [CrossRef]
- Yu, W.; Liang, X.; Li, X.; Zhang, Y.; Sun, Z.; Liu, Y.; Wang, J. MicroRNA-195: A review of its role in cancers. OTT 2018, 11, 7109–7123. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type of Study | Cell Culture/Species | Treatment Doses and Duration | Effects on miRNAs | Ref. |
|---|---|---|---|---|
| In vitro | Murine ovarian granulosa cells | 10, 20 and 40 µM 2, 4, 6, and 8 h |
| [56] |
| In vitro | Primary human proximal tubular epithelial cells (HPTECs) | 25 µM CdCl2 6 and 24 h |
| [58,59] |
| In vitro | Cd-transformed prostate epithelial cells (CTPE) developed from immortalized nontumorigenic human prostate epithelial cells (RWPE-1) | 10 µM 8 weeks |
| [68] |
| In vitro | Hepatoma cell line (HepG2) | 0.1–10 µM CdCl2 24, 48, and 72 h |
| [69] |
| In vivo | Male Sprague-Dawley rats | s.c. 0.6 mg/kg CdCl2 12 weeks |
| [58] |
| Type of Study | Cell Culture/Species | Treatment Doses and Duration | Effects on miRNAs | Ref. |
|---|---|---|---|---|
| In vitro | Human renal epithelial cells (HK-2) | 2.0–5.0 µM NaAsO2 30 weeks |
| [87] |
| In vitro | Pancreatic cancer cells Panc-1 and Patu8988 cells | 3 µmol/L As2O3 |
| [89] |
| In vitro | Normal human urothelial cells (HUC1) | 1 µM As2O3 2, 4, 6, 8, and 10 months |
| [88] |
| In vivo | Patients with urothelial carcinoma (urine samples) |
| ||
| In vitro | HaCaT cells (immortalized human keratinocytes) | 0.5 ppm 15 passages 60 days |
| [98] |
| In vivo | Peripheral blood mononuclear cells Chronically exposed humans with skin cancers (SCC and BCC) |
| ||
| In vivo | Keratinocytes Chronically exposed humans with skin cancers (SCC and BCC) |
| [100] | |
| Toxic Metal | Type of Study | Cell Culture/Species | Treatment Doses and Duration | Effects on miRNAs | Ref. |
|---|---|---|---|---|---|
| Pb | In vivo | Male pups from C57BL/6J strain mice | 0.2% Pb acetate via mothers’ milk Postnatal days (PND) 1–20 days |
| [108] |
| In vivo | Male Sprague-Dawley rats | 100, 200, and 300 ppm Pb acetate 8 weeks |
| [109] | |
| In vivo | Battery factories workers (blood samples) |
| [104] | ||
| Hg | In vivo | Mercury thermometer factory, female workers (blood samples) |
| [127] | |
| Mn | In vitro | Human neuroblastoma cells SH-SY5Y | 2 mM MnCl2 24 h |
| [134] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wallace, D.R.; Taalab, Y.M.; Heinze, S.; Tariba Lovaković, B.; Pizent, A.; Renieri, E.; Tsatsakis, A.; Farooqi, A.A.; Javorac, D.; Andjelkovic, M.; et al. Toxic-Metal-Induced Alteration in miRNA Expression Profile as a Proposed Mechanism for Disease Development. Cells 2020, 9, 901. https://doi.org/10.3390/cells9040901
Wallace DR, Taalab YM, Heinze S, Tariba Lovaković B, Pizent A, Renieri E, Tsatsakis A, Farooqi AA, Javorac D, Andjelkovic M, et al. Toxic-Metal-Induced Alteration in miRNA Expression Profile as a Proposed Mechanism for Disease Development. Cells. 2020; 9(4):901. https://doi.org/10.3390/cells9040901
Chicago/Turabian StyleWallace, David R., Yasmeen M. Taalab, Sarah Heinze, Blanka Tariba Lovaković, Alica Pizent, Elisavet Renieri, Aristidis Tsatsakis, Ammad Ahmad Farooqi, Dragana Javorac, Milena Andjelkovic, and et al. 2020. "Toxic-Metal-Induced Alteration in miRNA Expression Profile as a Proposed Mechanism for Disease Development" Cells 9, no. 4: 901. https://doi.org/10.3390/cells9040901
APA StyleWallace, D. R., Taalab, Y. M., Heinze, S., Tariba Lovaković, B., Pizent, A., Renieri, E., Tsatsakis, A., Farooqi, A. A., Javorac, D., Andjelkovic, M., Bulat, Z., Antonijević, B., & Buha Djordjevic, A. (2020). Toxic-Metal-Induced Alteration in miRNA Expression Profile as a Proposed Mechanism for Disease Development. Cells, 9(4), 901. https://doi.org/10.3390/cells9040901