Association of Proteins Modulating Immune Response and Insulin Clearance during Gestation with Antenatal Complications in Patients with Gestational or Type 2 Diabetes Mellitus

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Considerations
2.2. Population
2.2.1. Inclusion Criteria for Patients with GDM or T2DM
2.2.2. Diagnostic Criteria for Diabetic Fetopathy
2.3. Reagents
2.4. Blood Samples Collection and Handling
2.5. Liquid Chromatography and Surveying Mass Spectrometry
2.6. Accurate Quantitative Mass Spectrometry Analysis
2.7. LC-MS Data Analysis and Proteins Identification
2.8. Statistical Data Analysis and Functional Clustering Analysis.
3. Results
4. Discussion
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ramos-Leví, A.M.; Pérez-Ferre, N.; Fernández, M.D.; del Valle, L.; Bordiu, E.; Bedia, A.R. Risk Factors for Gestational Diabetes Mellitus in a Large Population of Women Living in Spain: Implications for Preventative Strategies. Int. J. Endocrinol. 2012, 2012, 312529. [Google Scholar] [CrossRef] [PubMed]
- Assaf-Balut, C.; García de la Torre, N.; Fuentes, M.; Durán, A.; Bordiú, E.; del Valle, L. A High Adherence to Six Food Targets of the Mediterranean Diet in the Late First Trimester is Associated with a Reduction in the Risk of Materno-Foetal Outcomes: The St. Carlos Gestational Diabetes Mellitus Prevention Study. Nutrients 2018, 11, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athukorala, C.; Rumbold, A.R.; Willson, K.J.; Crowther, C.A. The risk of adverse pregnancy outcomes in women who are overweight or obese. BMC Pregnancy Childbirth 2010, 10, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desoye, G.; Nolan, C.J. The fetal glucose steal: An underappreciated phenomenon in diabetic pregnancy. Diabetologia 2016, 59, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Grohmann, B.; Brazeau-Gravelle, P.; Momoli, F.; Moreau, K.; Zhang, T.; J Keely, E. Obstetric healthcare providers’ perceptions of communicating gestational weight gain recommendations to overweight/obese pregnant women. Obstet. Med. 2012, 5, 161–165. [Google Scholar] [CrossRef]
- Filardi, T.; Panimolle, F.; Crescioli, C.; Lenzi, A.; Morano, S. Gestational Diabetes Mellitus: The Impact of Carbohydrate Quality in Diet. Nutrients 2019, 11, 1549. [Google Scholar] [CrossRef] [Green Version]
- The “Global Report on Diabetes” of the World Health Organization Library; WHO Press: Geneva, Switzerland; Available online: https://www.who.int/diabetes/global-report/en/ (accessed on 10 January 2020).
- Jiwani, A.; Marseille, E.; Lohse, N.; Damm, P.; Hod, M.; Kahn, J.G. Gestational diabetes mellitus: Results from a survey of country prevalence and practices. J. Matern. Neonatal Med. 2012, 25, 600–610. [Google Scholar] [CrossRef]
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Dedov, I.I.; Shestakova, M.V.; Vikulova, O.K.; Zheleznyakova, A.V.; Isakov, M.A. Diabetes mellitus in Russian Federation: Prevalence, morbidity, mortality, parameters of glycaemic control and structure of glucose lowering therapy according to the federal diabetes register, status 2017. Diabetes Mellit. 2018, 21, 144–159. [Google Scholar] [CrossRef] [Green Version]
- Leipold, H.; Worda, C.; Schwindt, J.; Kautzky-Willer, A.; Bancher-Todesca, D.; Husslein, P.W. Severe diabetic fetopathy despite strict metabolic control. Wien. Klin. Wochenschr. 2005, 117, 561–564. [Google Scholar] [CrossRef]
- Bellamy, L.; Casas, J.-P.; Hingorani, A.D.; Williams, D. Type 2 diabetes mellitus after gestational diabetes: A systematic review and meta-analysis. Lancet 2009, 373, 1773–1779. [Google Scholar] [CrossRef]
- Filardi, T.; Tavaglione, F.; Di Stasio, M.; Fazio, V.; Lenzi, A.; Morano, S. Impact of risk factors for gestational diabetes (GDM) on pregnancy outcomes in women with GDM. J. Endocrinol. Investig. 2018, 41, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Jawerbaum, A.; Capobianco, E. Review: Effects of PPAR activation in the placenta and the fetus: Implications in maternal diabetes. Placenta 2011, 32, S212–S217. [Google Scholar] [CrossRef] [PubMed]
- Schaefer-Graf, U.M.; Meitzner, K.; Ortega-Senovilla, H. Differences in the implications of maternal lipids on fetal metabolism and growth between gestational diabetes mellitus and control pregnancies. Diabet. Med. 2011, 28, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.C.; Moore, M.P.; Gullam, J.E.; Mohamed, K.; Rowan, J. An early pregnancy HbA1c ≥ 5.9% (41 mmol/mol) is optimal for detecting diabetes and identifies women at increased risk of adverse pregnancy outcomes. Diabetes Care 2014, 37, 2953–2959. [Google Scholar] [CrossRef] [Green Version]
- Diagnostic criteria and classification of hyperglycaemia first detected in pregnancy: A World Health Organization Guideline. Diabetes Res. Clin. Pract. 2014, 103, 341–363. [CrossRef]
- Malin, G.L.; Bugg, G.J.; Takwoingi, Y.; Thornton, J.G.; Jones, N.W. Antenatal magnetic resonance imaging versus ultrasound for predicting neonatal macrosomia: A systematic review and meta-analysis. BJOG 2016, 123, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Flores, J.; Cruceyra, M.; Cañamares, M.; Garicano, A.; Espada, M.; Nieto, O. Sonographic Evaluation of Fetal Adrenal Gland in Gestational Diabetes. J. Ultrasound Med. 2017, 36, 999–1007. [Google Scholar] [CrossRef]
- Metzger, B.E. International Association of Diabetes and Pregnancy Study Groups Consensus Panel. International association of diabetes and pregnancy study groups recommendations on the diagnosis and classification of hyperglycemia in pregnancy. Diabetes Care 2010, 33, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Dedov, I.I.; Krasnopolskiy, V.I.; Sukhikh, G.T. Russian National Consensus Statement on gestational diabetes: Diagnostics, treatment and postnatal care. Diabetes Mellit. 2012, 15, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Ilgisonis, E.V.; Kopylov, A.T.; Ponomarenko, E.A.; Poverennaya, E.V.; Tikhonova, O.V.; Farafonova, T.E. Increased Sensitivity of Mass Spectrometry by Alkaline Two-Dimensional Liquid Chromatography: Deep Cover of the Human Proteome in Gene-Centric Mode. J. Proteome Res. 2018, 17, 4258–4266. [Google Scholar] [CrossRef] [PubMed]
- Beck, S.; Michalski, A.; Raether, O. The Impact II, a Very High-Resolution Quadrupole Time-of-Flight Instrument (QTOF) for Deep Shotgun Proteomics. Mol. Cell. Proteom. 2015, 14, 2014–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaudel, M.; Barsnes, H.; Berven, F.S.; Sickmann, A.; Martens, L. SearchGUI: An open-source graphical user interface for simultaneous OMSSA and X!Tandem searches. Proteomics 2011, 11, 996–999. [Google Scholar] [CrossRef] [PubMed]
- Vaudel, M.; Burkhart, J.M.; Zahedi, R.P.; Oveland, E.; Berven, F.S.; Sickmann, A. PeptideShaker enables reanalysis of MS-derived proteomics data sets. Nat. Biotechnol. 2015, 33, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Elias, J.E.; Gygi, S.P. Target-decoy search strategy for increased confidence in large-scale protein identifications by mass spectrometry. Nat. Methods 2007, 4, 207–214. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 2009, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, A.; Jupe, S.; Matthews, L. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef]
- Ravnsborg, T.; Svaneklink, S.; Andersen, L.L.T.; Larsen, M.R.; Jensen, D.M.; Overgaard, M. First-trimester proteomic profiling identifies novel predictors of gestational diabetes mellitus. PLoS ONE 2019, 14, e0214457. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Shen, L.; Wei, Y. Identification of candidate biomarkers for the prediction of gestational diabetes mellitus in the early stages of pregnancy using iTRAQ quantitative proteomics. Proteom. Clin. Appl. 2017, 11, 1600152. [Google Scholar] [CrossRef] [PubMed]
- Lappas, M. Lower circulating levels of complement split proteins C3a and C4a in maternal plasma of women with gestational diabetes mellitus. Diabet. Med. 2011, 28, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Qu, T.; He, X. Screening of differentially expressed proteins from syncytiotrophoblast for severe early-onset preeclampsia in women with gestational diabetes mellitus using tandem mass tag quantitative proteomics. BMC Egnancy Childbirth 2018, 18, 437. [Google Scholar] [CrossRef]
- Shen, Y.; Yang, J.; Li, J.; Shi, X.; Ouyang, L.; Tian, Y. Carnosine Inhibits the Proliferation of Human Gastric Cancer SGC-7901 Cells through Both of the Mitochondrial Respiration and Glycolysis Pathways. PLoS ONE 2014, 9, e104632. [Google Scholar] [CrossRef] [Green Version]
- Teufel, M.; Saudek, V.; Ledig, J.P.; Bernhardt, A.; Boularand, S.; Carreau, A.; Cairns, N.J.; Carter, C.; Cowley, D.J.; Duverger, D.; et al. Sequence identification and characterization of human carnosinase and a closely related non-specific dipeptidase. J. Biol. Chem. 2003, 278, 6521–6531. [Google Scholar] [CrossRef] [Green Version]
- Sauerhofer, S.; Yuan, G.; Braun, G.S.; Deinzer, M.; Neumaier, M.; Gretz, N.; Floege, J.; Kriz, W.; van der Woude, F.; Moeller, M.J. L-carnosine, a substrate of carnosinase-1, influences glucose metabolism. Diabetes 2007, 56, 2425–2432. [Google Scholar] [CrossRef] [Green Version]
- Vionnet, N.; Tregouet, D.; Kazeem, G.; Gut, I.; Groop, P.H.; Tarnow, L.; Parving, H.H.; Hadjadj, S.; Forsblom, C.; Farrall, M.; et al. Analysis of 14 candidate genes for diabetic nephropathy on chromosome 3q in European populations: Strongest evidence for association with a variant in the promoter region of the adiponectin gene. Diabetes 2006, 55, 3166–3174. [Google Scholar] [CrossRef]
- Wanic, K.; Placha, G.; Dunn, J.; Smiles, A.; Warram, J.H.; Krolewski, A.S. Exclusion of polymorphisms in carnosinase genes (CNDP1 and CNDP2) as a cause of diabetic nephropathy in type 1 diabetes: Results of large case-control and follow-up studies. Diabetes 2008, 57, 2547–2551. [Google Scholar] [CrossRef]
- Sleddering, M.A.; Markvoort, A.J.; Dharuri, H.K. Proteomic analysis in type 2 diabetes patients before and after a very low calorie diet reveals potential disease state and intervention specific biomarkers. PLoS ONE 2014, 9, e112835. [Google Scholar] [CrossRef] [Green Version]
- Pfister, F.; Riedl, E.; Wang, Q.; vom Hagen, F.; Deinzer, M.; Harmsen, M.C.; Molema, G.; Yard, B.; Feng, Y.; Hammes, H.P. Oral carnosine supplementation prevents vascular damage in experimental diabetic retinopathy. Cell. Physiol. Biochem. 2011, 28, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedl, E.; Pfister, F.; Braunagel, M.; Brinkkotter, P.; Sternik, P.; Deinzer, M.; Bakker, S.J.; Henning, R.H.; van den Born, J.; Kramer, B.K.; et al. Carnosine prevents apoptosis of glomerular cells and podocyte loss in STZ diabetic rats. Cell. Physiol. Biochem. 2011, 28, 279–288. [Google Scholar] [CrossRef]
- Mong, M.C.; Chao, C.Y.; Yin, M.C. Histidine and carnosine alleviated hepatic steatosis in mice consumed high saturated fat diet. Eur. J. Pharmacol. 2011, 653, 82–88. [Google Scholar] [CrossRef]
- Aldini, G.; Orioli, M.; Rossoni, G.; Savi, F.; Braidotti, P.; Vistoli, G.; Yeum, K.J.; Negrisoli, G.; Carini, M. The carbonyl scavenger carnosine ameliorates dyslipidaemia and renal function in Zucker obese rats. J. Cell. Mol. Med. 2011, 15, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Köninger, A.; Mathan, A.; Mach, P.; Frank, M.; Schmidt, B.; Schleussner, E. Is Afamin a novel biomarker for gestational diabetes mellitus? A pilot study. Reiprod. Biol. Endocrinol. 2018, 16, 30. [Google Scholar] [CrossRef]
- Scholze, A.; Bladbjerg, E.-M.; Sidelmann, J.J.; Diederichsen, A.C.P.; Mickley, H.; Nybo, M. Plasma concentrations of extracellular matrix protein fibulin-1 are related to cardiovascular risk markers in chronic kidney disease and diabetes. Cardiovasc. Diabetol. 2013, 12, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Gao, J.; Pu, C.; Feng, G.; Wang, L.; Huang, L. ApoM/HDL-C and apoM/apoA-I ratios are indicators of diabetic nephropathy in healthy controls and type 2 diabetes mellitus. Clin. Chim. Acta 2017, 466, 31–37. [Google Scholar] [CrossRef]
- Mach, P.; Gellhaus, A.; Prager, S. Soluble CEACAM1 and CEACAM6 are differently expressed in blood serum of pregnant women during normal pregnancy. Am. J. Reprod. Immunol. 2017, 78, e12700. [Google Scholar] [CrossRef]
- Najjar, S.M.; Philippe, N.; Suzuki, Y.; Ignacio, G.A.; Formisano, P.; Accili, D.; Taylor, S.I. Insulin-stimulated phosphorylation of recombinant pp120/HA4, an endogenous substrate of the insulin receptor tyrosine kinase. Biochemistry 1995, 34, 9341–9349. [Google Scholar] [CrossRef]
- Dubois, M.-J.; Bergeron, S.; Kim, H.-J.; Dombrowski, L.; Perreault, M.; Fournès, B. The SHP-1 protein tyrosine phosphatase negatively modulates glucose homeostasis. Nat. Med. 2006, 12, 549–556. [Google Scholar] [CrossRef]
- Najjar, S.M.; Yang, Y.; Fernström, M.A.; Lee, S.J.; Deangelis, A.M.; Rjaily, G.A. Insulin acutely decreases hepatic fatty acid synthase activity. Cell Metab. 2005, 2, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dentin, R.; Liu, Y.; Koo, S.-H.; Hedrick, S.; Vargas, T.; Heredia, J. Insulin modulates gluconeogenesis by inhibition of the coactivator TORC2. Nature 2007, 449, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Najjar, S.M.; Russo, L. CEACAM1 loss links inflammation to insulin resistance in obesity and non-alcoholic steatohepatitis (NASH). Semin. Immunopathol. 2014, 36, 55–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, S.O.; Hoogeveen, R.C.; Brancati, F.L. Association of blood lactate with type 2 diabetes: The Atherosclerosis Risk in Communities Carotid MRI Study. Int. J. Epidemiol. 2010, 39, 1647–1655. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Dong, Y.; Atefi, M.; Liu, Y.; Elshimali, Y.; Vadgama, J.V. Lactate, a Neglected Factor for Diabetes and Cancer Interaction. Mediat. Inflamm. 2016, 2016, 6456018. [Google Scholar] [CrossRef] [Green Version]
- Cerychova, R.; Pavlinkova, G. HIF-1, Metabolism, and Diabetes in the Embryonic and Adult Heart. Front. Endocrinol. 2018, 9, 460. [Google Scholar] [CrossRef] [Green Version]
- Peters, V.; Zschocke, J.; Schmitt, C.P. Carnosinase, diabetes mellitus and the potential relevance of carnosinase deficiency. J. Inherit. Metab. Dis. 2018, 41, 39–47. [Google Scholar] [CrossRef]
- Puglianiello, A.; Cianfarani, S. Central Control of Glucose Homeostasis. Rev. Diabet. Stud. 2006, 3, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Reaven, G.M.; Hollenbeck, C.; Jeng, C.-Y.; Wu, M.S.; Chen, Y.D.-I. Measurement of Plasma Glucose, Free Fatty Acid, Lactate, and Insulin for 24 h in Patients with NIDDM. Diabetes 1988, 37, 1020–1024. [Google Scholar] [CrossRef]
- Ahluwalia, T.S.; Lindholm, E.; Groop, L.C. Common variants in CNDP1 and CNDP2, and risk of nephropathy in type 2 diabetes. Diabetologia 2011, 54, 2295–2302. [Google Scholar] [CrossRef] [Green Version]
- Najjar, S.M. Regulation of insulin action by CEACAM1. Trends Endocrinol. Metab. 2002, 13, 240–245. [Google Scholar] [CrossRef]
- Muller, M.M. Transmembrane CEACAM1 affects integrin-dependent signaling and regulates extracellular matrix protein-specific morphology and migration of endothelial cells. Blood 2005, 105, 3925–3934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray-Owen, S.D.; Blumberg, R.S. CEACAM1: Contact-dependent control of immunity. Nat. Rev. Immunol. 2006, 6, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.; Dubois, M.-J.; Leung, N.; Charbonneau, A.; Turbide, C.; Avramoglu, R.K. Targeted Disruption of Carcinoembryonic Antigen-Related Cell Adhesion Molecule 1 Promotes Diet-Induced Hepatic Steatosis and Insulin Resistance. Endocrinology 2009, 150, 3503–3512. [Google Scholar] [CrossRef] [PubMed]
- Horst, A.K.; Najjar, S.M.; Wagener, C.; Tiegs, G. CEACAM1 in Liver Injury, Metabolic and Immune Regulation. Int. J. Mol. Sci. 2018, 19, 3110. [Google Scholar] [CrossRef] [Green Version]
- Bouché, C.; Serdy, S.; Kahn, C.R.; Goldfine, A.B. The Cellular Fate of Glucose and Its Relevance in Type 2 Diabetes. Endocr. Rev. 2004, 25, 807–830. [Google Scholar] [CrossRef]
- Boden, G.; Chen, X. Effects of fat on glucose uptake and utilization in patients with non-insulin-dependent diabetes. J. Clin. Investig. 1995, 96, 1261–1268. [Google Scholar] [CrossRef] [Green Version]
- Haffner, S.M. The Metabolic Syndrome: Inflammation, Diabetes Mellitus, and Cardiovascular Disease. Am. J. Cardiol. 2006, 97, 3–11. [Google Scholar] [CrossRef]
- Calle, M.C.; Fernandez, M.L. Inflammation and type 2 diabetes. Diabetes Metab. 2012, 38, 183–191. [Google Scholar] [CrossRef]
- Freeman, D.J.; Norrie, J.; Caslake, M.J.; Gaw, A.; Ford, I.; Lowe, G.D.O. C-Reactive Protein Is an Independent Predictor of Risk for the Development of Diabetes in the West of Scotland Coronary Prevention Study. Diabetes 2002, 51, 1596–1600. [Google Scholar] [CrossRef] [Green Version]
- Preventing Chronic Diseases. A Vital Investment: WHO Global Report; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Santangelo, C.; Filardi, T.; Perrone, G.; Mariani, M.; Mari, E.; Scazzocchio, B.; Masella, R.; Brunelli, R.; Lenzi, A.; Zicari, A.; et al. Cross-talk between fetal membranes and visceral adipose tissue involves HMGB1-RAGE and VIP-VPAC2 pathways in human gestational diabetes mellitus. Acta Diabetol. 2019, 56, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A.D. C-Reactive Protein, Interleukin 6, and Risk of Developing Type 2 Diabetes Mellitus. JAMA 2001, 286, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Gelaye, B.; Revilla, L.; Lopez, T.; Suarez, L.; Sanchez, S.E.; Hevner, K. Association between insulin resistance and c-reactive protein among Peruvian adults. Diabetol. Metab. Syndr. 2010, 2, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geerlings, S. Immune dysfunction in patients with diabetes mellitus (DM). FEMS Immunol. Med. Microbiol. 1999, 26, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Pickup, J.C.; Crook, M.A. Is Type II diabetes mellitus a disease of the innate immune system? Diabetologia 1998, 41, 1241–1248. [Google Scholar] [CrossRef] [Green Version]
- Ardawi, M.; Nasrat, H.A.N.; Bahnassy, A.A. Serum Immunoglobulin Concentrations in Diabetic Patients. Diabet. Med. 1994, 11, 384–387. [Google Scholar] [CrossRef]
- Mazer, N.A.; Welbourn, D.; Bernstein, R.K.; Rubinstein, A. Immunoglobulin Deficiency in Patients with Diabetes Mellitus. J. Allergy Clin. Immunol. 2011, 127, AB12. [Google Scholar] [CrossRef]
- Ayres, J.S.; Schneider, D.S. Tolerance of Infections. Ann. Rev. Immunol. 2012, 30, 271–294. [Google Scholar] [CrossRef]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef]

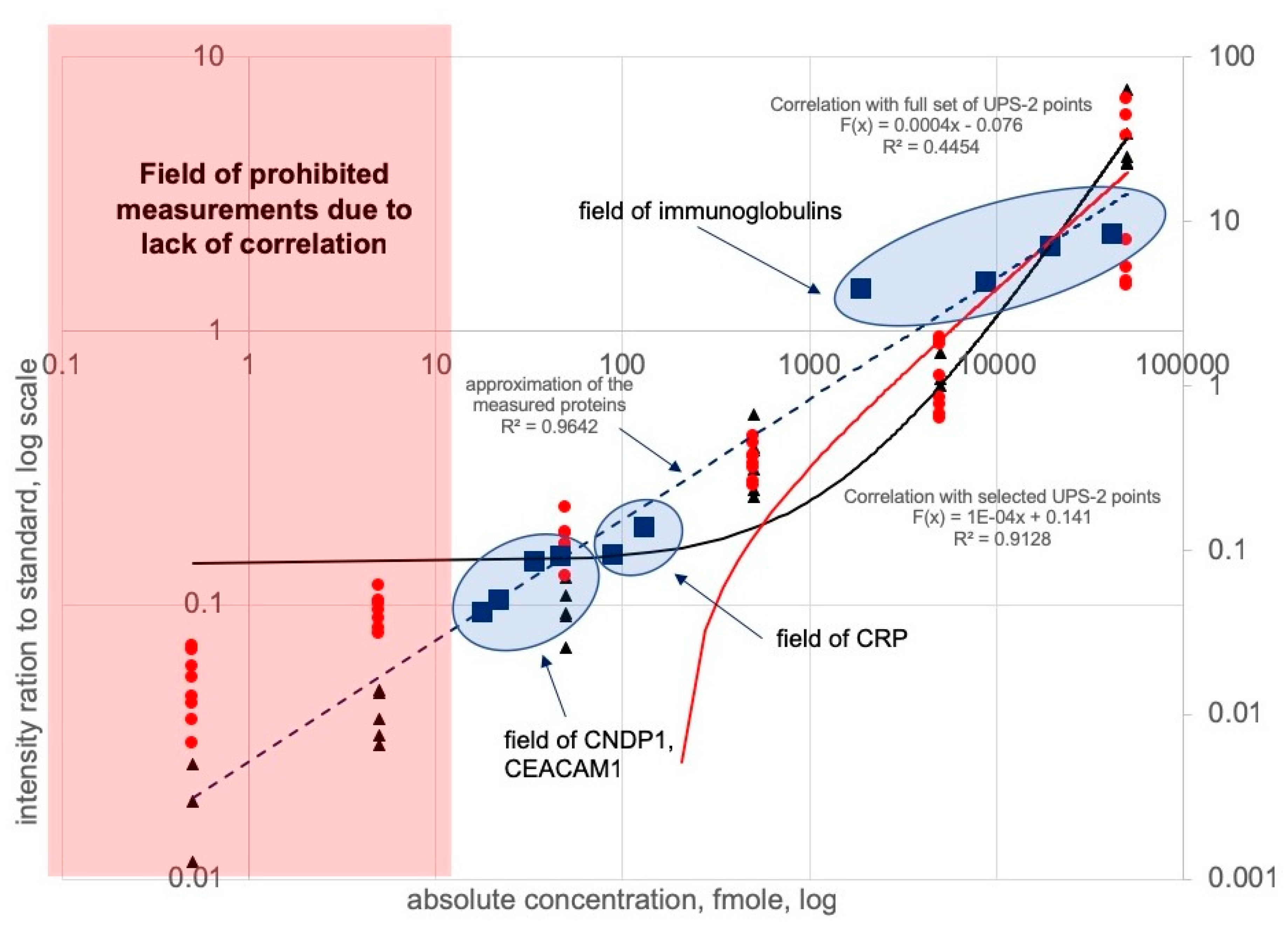
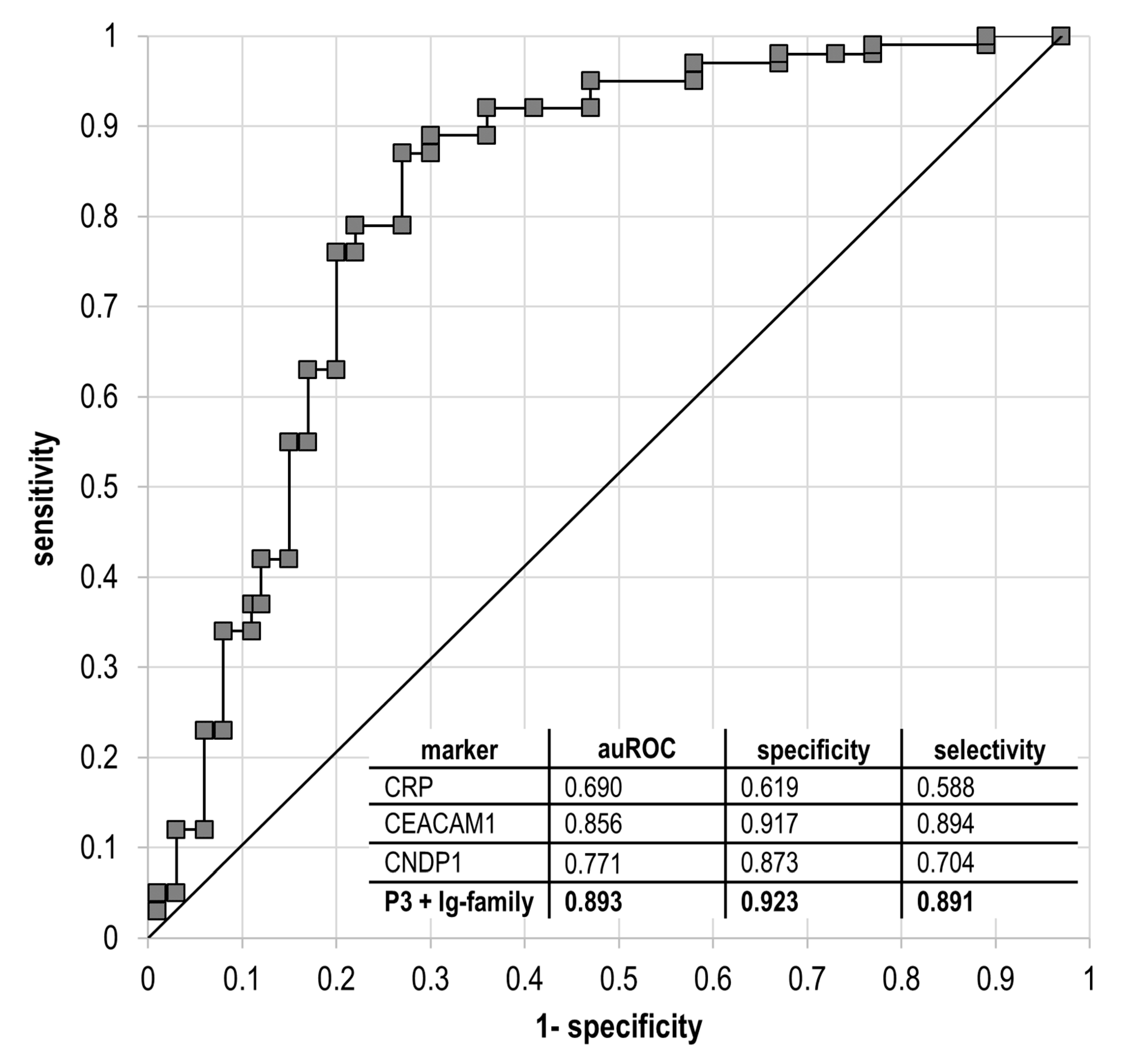


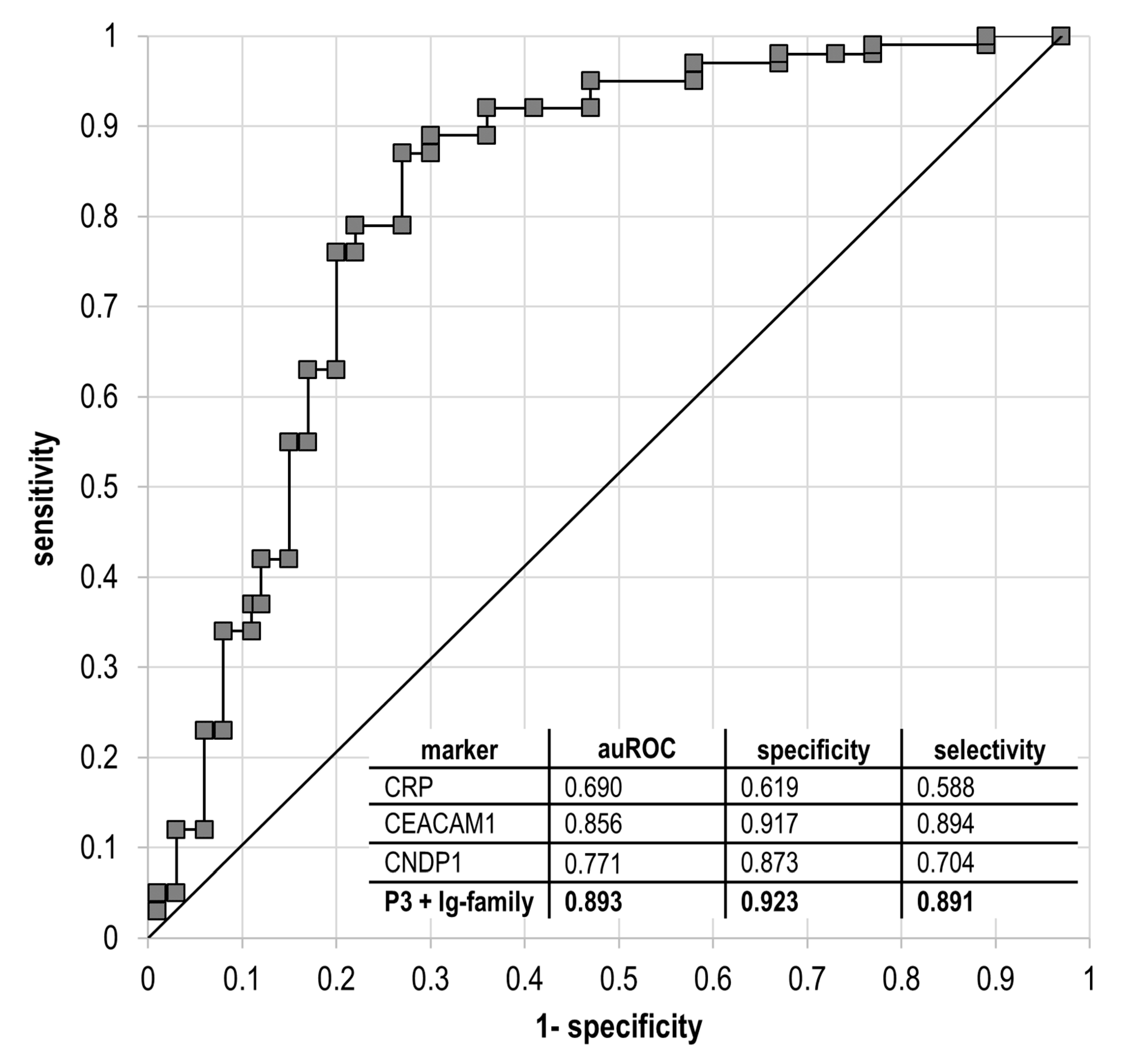
{kind=link}
{kind=link}
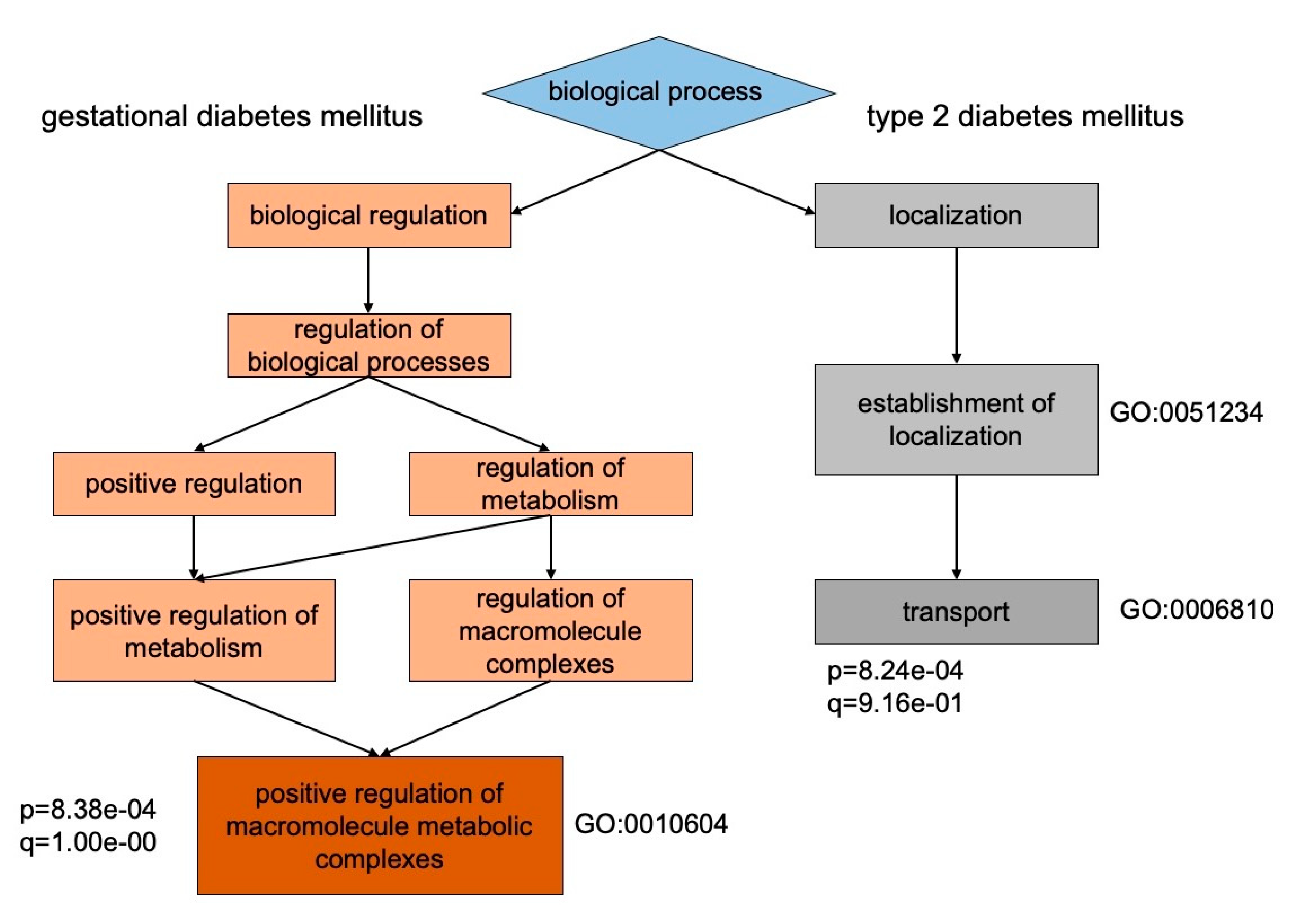
{kind=link}
{kind=link}
| Group Number | G01 | G02 | G03 | G04 | G05 | p-Value (Cut-Off p < 0.05) | |
|---|---|---|---|---|---|---|---|
| Groups Description | GDM (27% Insulin-Treated; 78% Dietary Intervention) | T2DM | Uncomplicated Pregnancy | ||||
| Ultrasound Examination | Normal Course | Diabetic Fetopathy | Normal Course | Diabetic Fetopathy | Normal Course | ||
| Group size (n) | 43 | 37 | 34 | 29 | 36 | 0.174 | |
| BMI ± SD, kg/m2 (between 16–19 weeks of gestation) # | 26.48 ± 5.41 (p = 0.007) & | 26.77 ± 4.99 (p = 0.003) & | 30.99 ± 3.29 (p = 0.02) & | 28.86 ± 6.20 (p = 0.008) & | 21.68 ± 4.25 | 0.517 | |
| Age ± SD, years | 27.2 ± 4.8 | 26.7 ± 5.4 | 27.7 ± 5.9 | 25.2 ± 5.1 | 26.6 ± 5.2 | 0.889 | |
| OGTT (75 g), mean ± SD, mmol/L | Fasting (>5.8 mmol/L) † | 6.1 ± 0.4 (p = 0.023) & | 6.0 ± 0.3 (p = 0.018) & | 9.7 ± 1.2 (p = 0.004) & | 9.9 ± 1.1 (p = 0.003) & | 3.8 ± 0.5 | 0.037 |
| 1 h (>9.8 mmol/L) ‡ | 9.9 ± 1.1 | 10.6 ± 0.7 | N/A | N/A | N/A | 0.641 | |
| 2 h (>8.5 mmol/L) ‡ | 8.8 ± 0.4 | 9.2 ± 0.4 | N/A | N/A | N/A | 0.820 | |
| HbA1c, mean ± SD, % | I trimester | 4.9 ± 0.3 | 5.6 ± 0.4 | 5.6 ± 0.5 | 5.5 ± 0.8 | 3.1 ± 1.5 | 0.917 |
| II trimester | 6.7 ± 1.1 | 6.2 ± 2.0 | 8.2 ± 0.7 | 8.2 ± 2.5 | 3.5 ± 1.4 | 0.602 | |
| III trimester | 6.3 ± 0.7 | 5.9 ± 0.6 | 6.4 ± 1.0 | 6.8 ± 0.9 | 3.9 ± 0.9 | 0.873 | |
| Caesar delivery, % | emergency | 2.33 | 8.11 | 0.00 | 0.00 | 5.56 | N/A |
| planned | 27.91 | 29.73 | 29.41 | 34.48 | 36.11 | N/A | |
| Gestational age at delivery, day; median (range) | 270 (259–278) | 265 (258–270) | 273 (268–289) | 270 (269–284) | 272 (265–288) | 0.902 | |
| Maternal weight gain, kg | 9.4 ± 4 | 11.7 ± 6 | 10.8 ± 5 | 12.1 ± 5 | 10.3 ± 6 | 0.673 | |
| Fetal weight, kg | 3157 ± 254 | 4189 ± 212 | 3186 ± 198 | 4402 ± 278 | 3207 ± 112 | 0.044 | |
| Family history of diabetes | 5 | 4 | 4 | 5 | 0 | N/A | |
| Previous GDM history | 6 | 3 | 0 | 0 | 0 | N/A | |
| Type of Diabetes Mellitus | GO Identifier | Annotation of Cellular Localization | p-Value | FDR q-Value | Enrichment Rate | Gene and Recommended Protein Names |
|---|---|---|---|---|---|---|
| T2DM | GO:0044424 | intracellular part | 8.2 × 10−4 | 1.7 × 10−1 | 1.14 | ARL17B; CNGA2; CST3; IFI16; PPIA; GLTPD2; RIMS1; RPL13; GOLGA4; TEX26; GOLT1A; U2AF1L4; PACS2; CAPRIN2; COL6A3; EPB42; SEMA6D; CAT; HBM; DGKH; SARS; SPTB; SLC4A1; DENND1C; BLVRB; CDC40; ARHGAP18; ZNF883; ANK1; HSPA8 |
| GDM | GO:0070062 | extracellular and exosome | 3.7 × 10−6 | 3.2 × 10−4 | 2.37 | PDE8A; MASP2; SAA2; TUBB; EZR; QSOX1; IGF2R; LBP; CD14; SEPP1; RRAS2; SOD1; AGRN; TNXB; ABCB1; SERPINA5; LTF; TXN |
| Acc. Number | Gene Name | Protein Name | G01, GDM | G02, GDM + DF | G03, T2DM | G04, T2DM + DF | G05, Control | p−Value † |
|---|---|---|---|---|---|---|---|---|
| P02741 | CRP | C-reactive protein, mcg/mL | 2.04 ± 1.32 | 5.29 ± 1.82 | 2.84 ± 0.67 | 4.21 ± 1.37 | 1.97 ± 0.71 | 3.12 × 10−4 |
| P13688 | CEACAM1 | Carcinoembryonic antigen-related cell adhesion molecule 1, ng/mL | 291.62 ± 34.55 | 113.18 ± 16.23 | 311.17 ± 42.16 | 81.09 ± 10.54 | 515.6 ± 72.14 | 5.33 × 10−3 |
| Q96KN2 | CNDP1 | Beta-Ala-His dipeptidase, ng/mL | 32.4 ± 5.23 | 49.3 ± 5.18 | 27.4 ± 2.63 | 37.7 ± 3.34 | 17.1 ± 4.31 | 2.17 × 10−4 |
| Acc. Number | Gene Name | Protein Name | G01, GDM | G02, GDM + DF | G03, T2DM | G04, T2DM + DF | G05, Control | p-Value † |
|---|---|---|---|---|---|---|---|---|
| P01857 | IGHG1 | Immunoglobulin IgG1 | 8.84 ± 1.22 | 7.81 ± 1.69 | 8.26 ± 2.09 | 7.65 ± 2.13 | 7.07 ± 1.99 | 1.032 × 10−3 |
| P01859 | IGHG2 | Immunoglobulin IgG2 | 2.27 ± 0.41 | 2.43 ± 0.37 | 2.54 ± 0.31 | 2.37 ± 0.22 | 2.11 ± 0.14 | 0.774 × 10−3 |
| P01860 | IGHG3 | Immunoglobulin IgG3 | 2.06 ± 0.49 | 2.12 ± 0.38 | 1.87 ± 0.24 | 2.05 ± 0.38 | 1.93 ± 0.65 | 0.591 × 10−2 |
| P01861 | IGHG4 | Immunoglobulin IgG4 | 2.12 ± 0.42 | 1.85 ± 0.36 | 2.08 ± 0.49 | 1.81 ± 0.41 | 2.64 ± 0.98 | 1.065 × 10−3 |
| P01871 | IGHM | Immunoglobulin IgM | 1.87 ± 0.31 | 1.67 ± 0.20 | 1.93 ± 0.26 | 1.71 ± 0.17 | 1.34 ± 0.12 | 4.912 × 10−3 |
| P01876 | IGHA1 | Immunoglobulin IgA1 | 1.42 ± 0.26 | 1.98 ± 0.14 | 1.39 ± 0.19 | 1.55 ± 0.28 | 1.31 ± 0.23 | 2.817 × 10−4 |
| P01877 | IGHA2 | Immunoglobulin IgA2 | 0.41 ± 0.08 | 0.52 ± 0.04 | 0.38 ± 0.07 | 0.46 ± 0.07 | 0.23 ± 0.09 | 1.296 × 10−3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopylov, A.T.; Kaysheva, A.L.; Papysheva, O.; Gribova, I.; Kotaysch, G.; Kharitonova, L.; Mayatskaya, T.; Krasheninnikova, A.; Morozov, S.G. Association of Proteins Modulating Immune Response and Insulin Clearance during Gestation with Antenatal Complications in Patients with Gestational or Type 2 Diabetes Mellitus. Cells 2020, 9, 1032. https://doi.org/10.3390/cells9041032
Kopylov AT, Kaysheva AL, Papysheva O, Gribova I, Kotaysch G, Kharitonova L, Mayatskaya T, Krasheninnikova A, Morozov SG. Association of Proteins Modulating Immune Response and Insulin Clearance during Gestation with Antenatal Complications in Patients with Gestational or Type 2 Diabetes Mellitus. Cells. 2020; 9(4):1032. https://doi.org/10.3390/cells9041032
Chicago/Turabian StyleKopylov, Arthur T., Anna L. Kaysheva, Olga Papysheva, Iveta Gribova, Galina Kotaysch, Lubov Kharitonova, Tatiana Mayatskaya, Anna Krasheninnikova, and Sergey G. Morozov. 2020. "Association of Proteins Modulating Immune Response and Insulin Clearance during Gestation with Antenatal Complications in Patients with Gestational or Type 2 Diabetes Mellitus" Cells 9, no. 4: 1032. https://doi.org/10.3390/cells9041032