(R)-Salbutamol Improves Imiquimod-Induced Psoriasis-Like Skin Dermatitis by Regulating the Th17/Tregs Balance and Glycerophospholipid Metabolism

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Mice Treatments
2.3. Evaluating the Severity of Skin Inflammation
2.4. Histopathological Examination
2.5. Hematological Analysis
2.6. IL-17 ELISA
2.7. Intracellular Staining and Flow Cytometry
2.8. Metabolomic Analysis
2.9. Statistical Analysis
3. Results
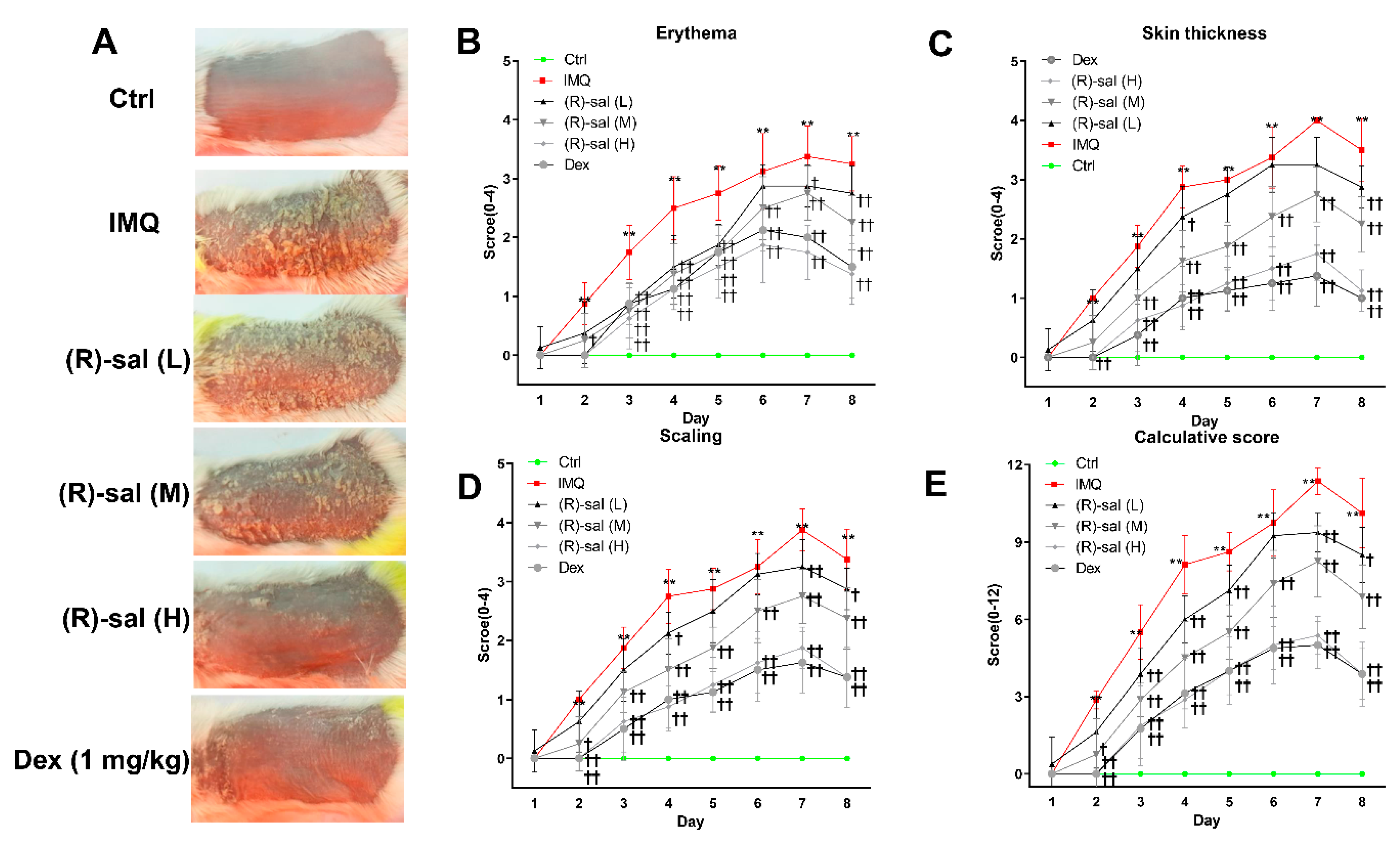
3.1. (R)-Salbutamol Alleviates Psoriatic Dermatiti
3.2. (R)-Salbutamol Alleviates the Pathology Changes Alterations Caused by IMQ on Mice Skin
3.3. Effect of (R)-Salbutamol on Haematological Parameters of IMQ-Induced Mouse Psoriasis
3.4. (R)-Salbutamol Reduced IL-17 Secretion in Mice Plasma
3.5. Effect of (R)-Salbutamol Treatment on the Ratio of Spleen Weight to Body Weight
3.6. (R)-Salbutamol Immune-Regulates the Number of CD3+CD4+ T Cells in Mice Treated with IMQ
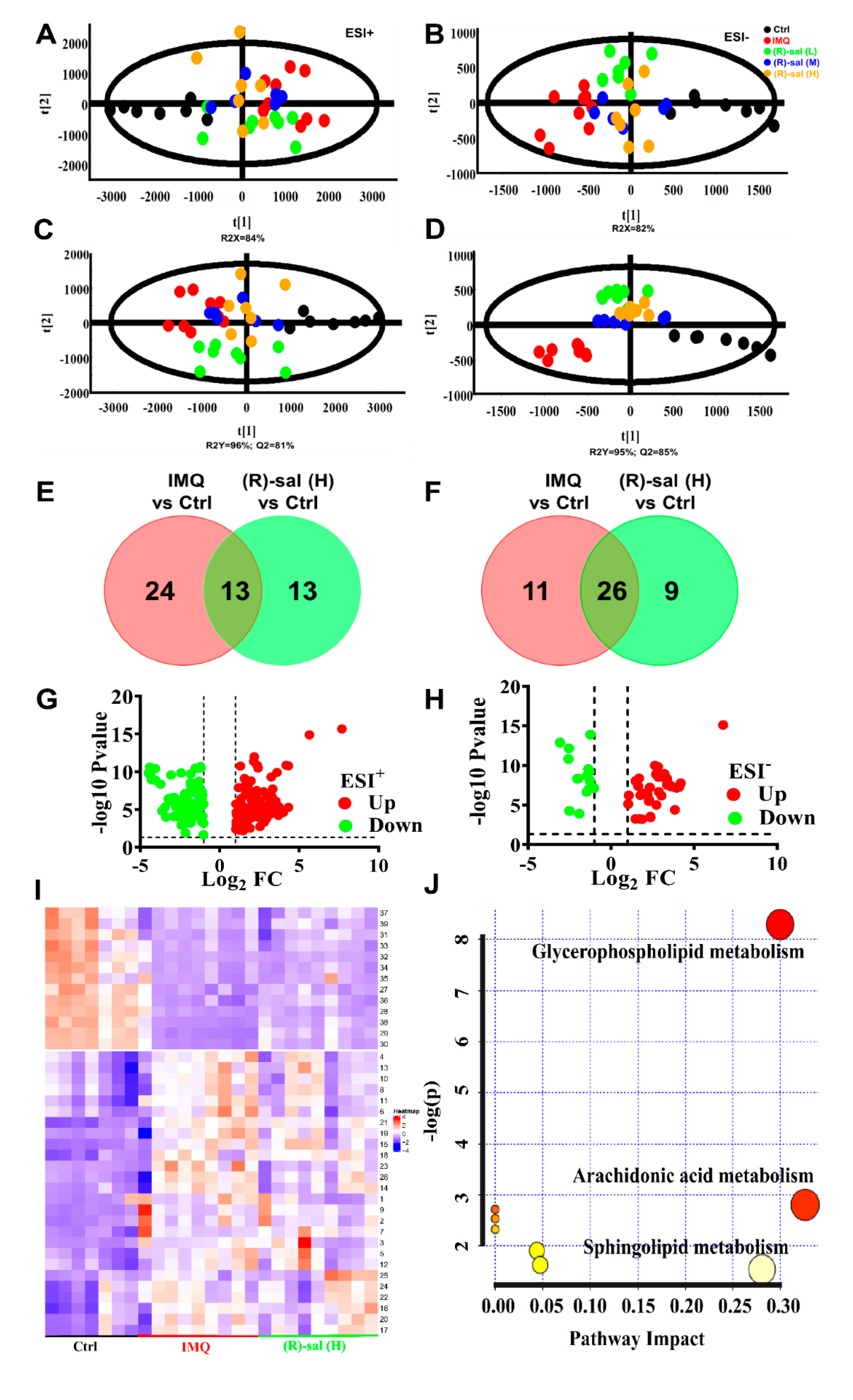
3.7. The Influence of (R)-Salbutamol on Metabolic Effects of IMQ Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chiang, C.C.; Cheng, W.J.; Korinek, M.; Lin, C.Y.; Hwang, T.L. Neutrophils in Psoriasis. Front. Immunol. 2019, 10, 2376. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.S.; Diamond, A.; Russell, A.; Jameson, J.M. Human αβ and γδ T Cells in Skin Immunity and Disease. Front. Immunol. 2018, 9, 1304. [Google Scholar] [CrossRef] [PubMed]
- Traidl, C.; Jugert, F.; Krieg, T.; Merk, H.; Hunzelmann, N. Inhibition of allergic contact dermatitis to DNCB but not to oxazolone in interleukin-4-deficient mice. J. Investig. Dermatol. 1999, 112, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Paschoal, R.S.; Silva, D.A.; Cardili, R.N.; Souza, C.D.S. Metabolic syndrome, C-reactive protein and cardiovascular risk in psoriasis patients: A cross-sectional study. An. Bras. Dermatol. 2018, 93, 222–228. [Google Scholar] [CrossRef]
- Semenov, Y.R.; Herbosa, C.M.; Rogers, A.T.; Huang, A.; Kwatra, S.G.; Cohen, B.; Anadkat, M.J.; Silverberg, J.I. Psoriasis and Mortality in the US: Data from the National Health and Nutrition Examination Survey. J. Am. Acad. Dermatol. 2019. [Google Scholar] [CrossRef]
- Peters, E.M.J. Stressed skin-a molecular psychosomatic update on stress-causes and effects in dermatologic diseases. JDDG J. Dtsch. Dermatol. Ges. 2016, 14, 233–252. [Google Scholar] [CrossRef]
- Mease, P.J.; Menter, M.A. Quality-of-life issues in psoriasis and psoriatic arthritis: Outcome measures and therapies from a dermatological perspective. J. Am. Acad. Dermatol. 2006, 54, 685–704. [Google Scholar] [CrossRef]
- Wang, C.; Yuan, J.; Wu, H.X.; Chang, Y.; Wang, Q.T.; Wu, Y.J.; Zhou, P.; Yang, X.D.; Yu, J.; Wei, W. Total glucosides of paeony inhibit the inflammatory responses of mice with allergic contact dermatitis by restoring the balanced secretion of pro-/anti-inflammatory cytokines. Int. Immunopharmacol. 2015, 24, 325–334. [Google Scholar] [CrossRef]
- Prinz, I.; Sandrock, I.; Mrowietz, U. Interleukin-17 cytokines: Effectors and targets in psoriasis-A breakthrough in understanding and treatment. J. Exp. Med. 2019. [Google Scholar] [CrossRef]
- Kalekar, L.A.; Rosenblum, M.D. Regulatory T cells in inflammatory skin disease: From mice to humans. Int. Immunol. 2019, 31, 457–463. [Google Scholar] [CrossRef]
- Chesne, J.; Braza, F.; Mahay, G.; Brouard, S.; Aronica, M.; Magnan, A. IL-17 in severe asthma. Where do we stand? Am. J. Respir. Crit. Care Med. 2014, 190, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Rinschen, M.M.; Ivanisevic, J.; Giera, M.; Siuzdak, G. Identification of bioactive metabolites using activity metabolomics. Nat. Rev. Mol. Cell Biol. 2019, 20, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Wheelock, C.E.; Wheelock, A.M.; Kawashima, S.; Diez, D.; Kanehisa, M.; van Erk, M.; Kleemann, R.; Haeggstrom, J.Z.; Goto, S. Systems biology approaches and pathway tools for investigating cardiovascular disease. Mol. Biosyst. 2009, 5, 588–602. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.; Wilson, I.D.; Nicholson, J.K. Metabolic phenotyping in health and disease. Cell 2008, 134, 714–717. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, A.-H.; Miao, J.-H.; Sun, H.; Han, Y.; Yan, G.-L.; Wu, F.-F.; Wang, X.-J. Metabolomics biotechnology, applications, and future trends: A systematic review. RSC Adv. 2019, 9, 37245–37257. [Google Scholar] [CrossRef]
- Lindon, J.C.; Holmes, E.; Nicholson, J.K. Metabonomics and its role in drug development and disease diagnosis. Expert Rev. Mol. Diagn. 2004, 4, 189–199. [Google Scholar] [CrossRef]
- Ueharaguchi, Y.; Honda, T.; Kusuba, N.; Hanakawa, S.; Adachi, A.; Sawada, Y.; Otsuka, A.; Kitoh, A.; Dainichi, T.; Egawa, G.; et al. Thromboxane A2 facilitates IL-17A production from Vgamma4(+) gammadelta T cells and promotes psoriatic dermatitis in mice. J. Allergy Clin. Immunol. 2018, 142, 680–683.e2. [Google Scholar] [CrossRef]
- Yan, J. Identifying biomarkers in human psoriasis: Revealed by a systems metabolomics approach. Br. J. Dermatol. 2017, 176, 555–557. [Google Scholar] [CrossRef]
- Bazargani, Y.T.; de Boer, A.; Leufkens, H.G.; Mantel-Teeuwisse, A.K. Essential medicines for COPD and asthma in low and middle-income countries. Thorax 2014, 69, 1149–1151. [Google Scholar] [CrossRef]
- Tanaka, S.; Tsutsui, M.; Kishida, T.; Souma, S.; Kuroda1, J.; Yoshida, T. Salbutamol inhibits lipopolysaccharide-induced inflammatory responses in rat peritoneal macrophages. J. Toxicol. Sci. 2010, 35, 327–334. [Google Scholar] [CrossRef]
- Jemec, G.B.; Ullman, S.; Goodfield, M.; Bygum, A.; Olesen, A.B.; Berth-Jones, J.; Nyberg, F.; Cramers, M.; Faergemann, J.; Andersen, P.; et al. A randomized controlled trial of R-salbutamol for topical treatment of discoid lupus erythematosus. Br. J. Dermatol. 2009, 161, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Gruber, W.; Frischauf, A.M.; Aberger, F. An old friend with new skills: Imiquimod as novel inhibitor of Hedgehog signaling in basal cell carcinoma. Oncoscience 2014, 1, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Van der Fits, L.; Mourits, S.; Voerman, J.S.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.M.; Florencia, E.; Prens, E.P.; et al. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J. Immunol. 2009, 182, 5836–5845. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Garrett, T.J.; Bowden, J.A. Optimization of Folch, Bligh-Dyer, and Matyash Sample-to Extraction Solvent Ratios for Human Plasma-Based Lipidomics Studies. Anal. Chim. Acta 2018, 1037. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Q.; Hu, C.; Yu, D.; Wang, L.; Zhou, Y.; Zhao, X.; Xu, G. Development of a High Coverage Pseudotargeted Lipidomics Method Based on Ultra-High Performance Liquid Chromatography–Mass Spectrometry. Anal. Chem. 2018. [Google Scholar] [CrossRef]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef]
- Gika, H.G.; Macpherson, E.; Theodoridis, G.A.; Wilson, I.D. Evaluation of the repeatability of ultra-performance liquid chromatography-TOF-MS for global metabolic profiling of human urine samples. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 871, 299–305. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Prenni, J.E. Stacked Injections of Biphasic Extractions for Improved Metabolomic Coverage and Sample Throughput. Anal. Chem. 2018, 90, 1147–1153. [Google Scholar] [CrossRef]
- Mattozzi, C.; Salvi, M.; D’Epiro, S.; Giancristoforo, S.; Macaluso, L.; Luci, C.; Lal, K.; Calvieri, S.; Richetta, A.G. Importance of regulatory T cells in the pathogenesis of psoriasis: Review of the literature. Dermatology 2013, 227, 134–145. [Google Scholar] [CrossRef]
- Vida, G.; Pena, G.; Kanashiro, A.; Thompson-Bonilla Mdel, R.; Palange, D.; Deitch, E.A.; Ulloa, L. beta2-Adrenoreceptors of regulatory lymphocytes are essential for vagal neuromodulation of the innate immune system. FASEB J. 2011, 25, 4476–4485. [Google Scholar] [CrossRef]
- Beng, H.; Su, H.; Wang, S.; Kuai, Y.; Hu, J.; Zhang, R.; Liu, F.; Tan, W. Differential effects of inhaled R- and S-terbutaline in ovalbumin-induced asthmatic mice. Int. Immunopharmacol. 2019, 73, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, F.; Tan, K.S.; Ser, H.L.; Tan, L.T.; Lee, L.H.; Tan, W. Effect of (R)-salbutamol on the switch of phenotype and metabolic pattern in LPS-induced macrophage cells. J. Cell. Mol. Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Uzkeser, H.; Cadirci, E.; Halici, Z.; Odabasoglu, F.; Polat, B.; Yuksel, T.N.; Ozaltin, S.; Atalay, F. Anti-inflammatory and antinociceptive effects of salbutamol on acute and chronic models of inflammation in rats: Involvement of an antioxidant mechanism. Mediat. Inflamm. 2012, 2012, 438912. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, G.; Pallone, F.; MacDonald, T.T.; Chimenti, S.; Costanzo, A. Psoriasis: From pathogenesis to novel therapeutic approaches. Clin. Sci. 2011, 120, 1–11. [Google Scholar] [CrossRef]
- Prinz, J.C. Autoimmune aspects of psoriasis: Heritability and autoantigens. Autoimmun. Rev. 2017, 16, 970–979. [Google Scholar] [CrossRef]
- Hawkes, J.E.; Chan, T.C.; Krueger, J.G. Psoriasis pathogenesis and the development of novel targeted immune therapies. J. Allergy Clin. Immunol. 2017, 140, 645–653. [Google Scholar] [CrossRef]
- Agrawal, S.; Gupta, S. TLR1/2, TLR7, and TLR9 signals directly activate human peripheral blood naive and memory B cell subsets to produce cytokines, chemokines, and hematopoietic growth factors. J. Clin. Immunol. 2011, 31, 89–98. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Sokolowska-Wojdylo, M.; Ruckemann-Dziurdzinska, K.; Roszkiewicz, J.; Nowicki, R.J. Chemokines and cytokines network in the pathogenesis of the inflammatory skin diseases: Atopic dermatitis, psoriasis and skin mastocytosis. Adv. Dermatol. Allergol. 2014, 31, 84–91. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Q.; Liu, H.; Lu, C.; Liang, C.L.; Qiu, F.; Han, L.; Dai, Z. Esculetin Ameliorates Psoriasis-Like Skin Disease in Mice by Inducing CD4(+)Foxp3(+) Regulatory T Cells. Front. Immunol. 2018, 9, 2092. [Google Scholar] [CrossRef]
- Cai, Y.; Fleming, C.; Yan, J. New insights of T cells in the pathogenesis of psoriasis. Cell Mol. Immunol. 2012, 9, 302–309. [Google Scholar] [CrossRef]
- Noack, M.; Miossec, P. Th17 and regulatory T cell balance in autoimmune and inflammatory diseases. Autoimmun. Rev. 2014, 13, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Haak, S.G.G.; Becher, B. Th17 cells in autoimmune disease: Changing the verdict. Immunotherapy 2009, 1, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Littman, D.R.; Rudensky, A.Y. Th17 and regulatory T cells in mediating and restraining inflammation. Cell 2010, 140, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Barbi, J.; Pardoll, D.; Pan, F. Metabolic control of the Treg/Th17 axis. Immunol. Rev. 2013, 252, 52–77. [Google Scholar] [CrossRef]
- Chen, S.; Li, H.; Liu, Y.; Zhu, Z.; Wei, Q. Quercitrin extracted from Tartary buckwheat alleviates imiquimod-induced psoriasis-like dermatitis in mice by inhibiting the Th17 cell response. J. Funct. Foods 2017, 38, 9–19. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Soma, T.; Takaku, Y.; Nakagome, K.; Hagiwara, K.; Kanazawa, M.; Nagata, M. Salbutamol modulates the balance of Th1 and Th2 cytokines by mononuclear cells from allergic asthmatics. Int. Arch. Allergy Immunol. 2010, 152, 32–40. [Google Scholar] [CrossRef]
- Elmore, S.A. Enhanced histopathology of the spleen. Toxicol. Pathol. 2006, 34, 648–655. [Google Scholar] [CrossRef]
- Pietrzak, A.; Michalak-Stoma, A.; Chodorowska, G.; Szepietowski, J.C. Lipid disturbances in psoriasis: An update. Mediat. Inflamm. 2010, 2010. [Google Scholar] [CrossRef]
- Ottas, A.; Fishman, D.; Okas, T.L.; Kingo, K.; Soomets, U. The metabolic analysis of psoriasis identifies the associated metabolites while providing computational models for the monitoring of the disease. Arch. Dermatol. Res. 2017, 309, 519–528. [Google Scholar] [CrossRef]
- Hishikawa, D.; Hashidate, T.; Shimizu, T.; Shindou, H. Diversity and function of membrane glycerophospholipids generated by the remodeling pathway in mammalian cells. J. Lipid Res. 2014, 55, 799–807. [Google Scholar] [CrossRef]
- Schulze, T.; Golfier, S.; Tabeling, C.; Rabel, K.; Graler, M.H.; Witzenrath, M.; Lipp, M. Sphingosine-1-phospate receptor 4 (S1P(4)) deficiency profoundly affects dendritic cell function and TH17-cell differentiation in a murine model. FASEB J. 2011, 25, 4024–4036. [Google Scholar] [CrossRef] [PubMed]
- Bai, A.; Robson, S. Beyond ecto-nucleotidase: CD39 defines human Th17 cells with CD161. Purinergic Signal. 2015, 11, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.H.; Cho, K.A.; Hahn, S.; Lee, Y.; Kim, Y.H.; Woo, S.Y.; Ryu, K.H.; Park, W.J.; Park, J.W. Inhibiting Sphingosine Kinase 2 Derived-sphingosine-1-phosphate Ameliorates Psoriasis-like Skin Disease via Blocking Th17 Differentiation of Naive CD4 T Lymphocytes in Mice. Acta Derm. Venereol. 2019, 99, 594–601. [Google Scholar] [CrossRef] [PubMed]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Wang, S.; Liu, B.; Wang, Y.; Tan, W. (R)-Salbutamol Improves Imiquimod-Induced Psoriasis-Like Skin Dermatitis by Regulating the Th17/Tregs Balance and Glycerophospholipid Metabolism. Cells 2020, 9, 511. https://doi.org/10.3390/cells9020511
Liu F, Wang S, Liu B, Wang Y, Tan W. (R)-Salbutamol Improves Imiquimod-Induced Psoriasis-Like Skin Dermatitis by Regulating the Th17/Tregs Balance and Glycerophospholipid Metabolism. Cells. 2020; 9(2):511. https://doi.org/10.3390/cells9020511
Chicago/Turabian StyleLiu, Fei, Shanping Wang, Bo Liu, Yukun Wang, and Wen Tan. 2020. "(R)-Salbutamol Improves Imiquimod-Induced Psoriasis-Like Skin Dermatitis by Regulating the Th17/Tregs Balance and Glycerophospholipid Metabolism" Cells 9, no. 2: 511. https://doi.org/10.3390/cells9020511
APA StyleLiu, F., Wang, S., Liu, B., Wang, Y., & Tan, W. (2020). (R)-Salbutamol Improves Imiquimod-Induced Psoriasis-Like Skin Dermatitis by Regulating the Th17/Tregs Balance and Glycerophospholipid Metabolism. Cells, 9(2), 511. https://doi.org/10.3390/cells9020511