Analysis of Activity-Dependent Energy Metabolism in Mice Reveals Regulation of Mitochondrial Fission and Fusion mRNA by Voluntary Physical Exercise in Subcutaneous Fat from Male Marathon Mice (DUhTP)

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
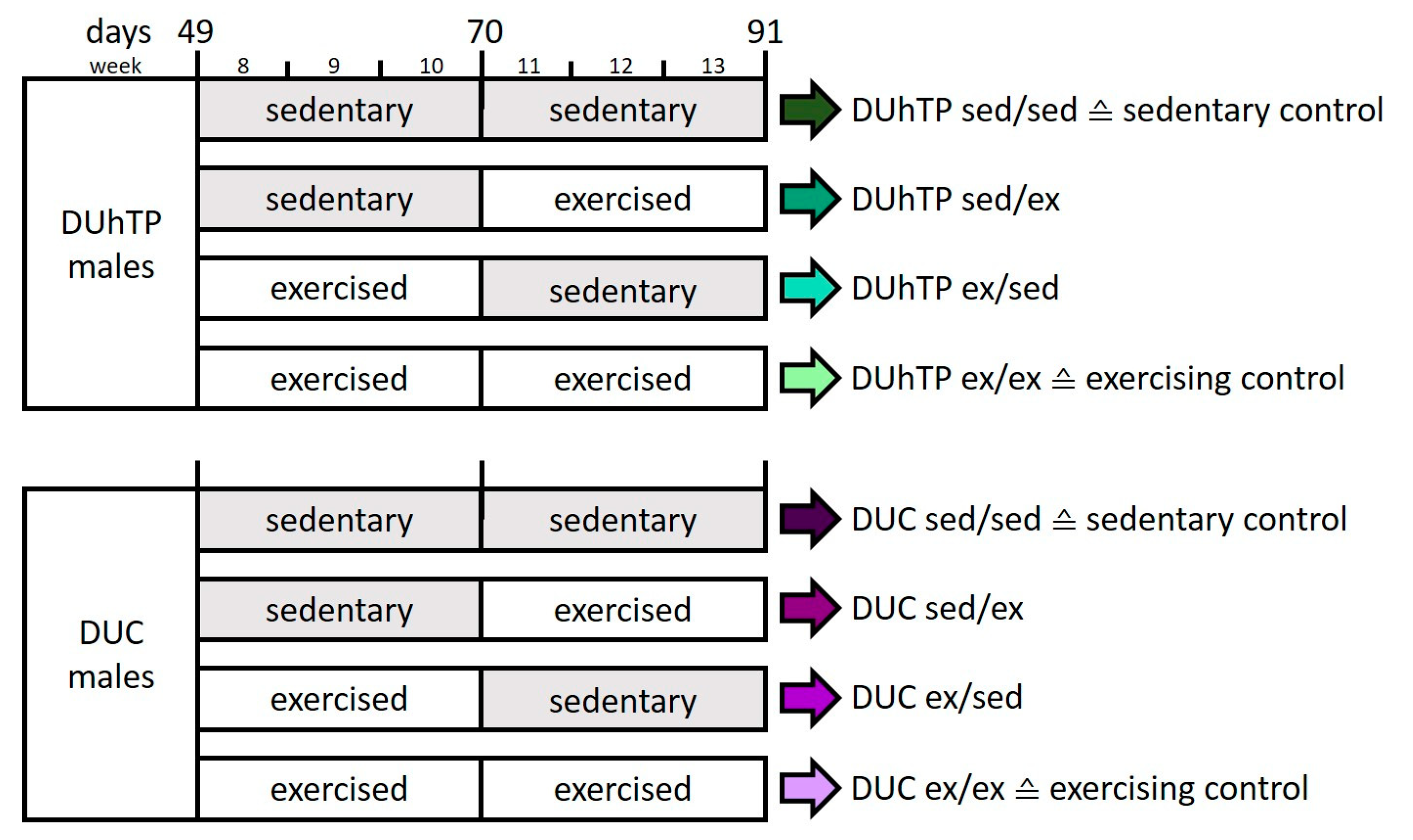
2.1. Animals and Cross-Over Study Design
2.2. Analysis of Triglycerides in Serum Samples
2.3. mRNA Expression in Subcutaneous Fat
2.4. Mitochondrial Morphology in Adipose Tissue
2.5. Statistical Analysis
3. Results
3.1. Running Wheel Performance and Food Consumption in Response to Differential Exercise Programs
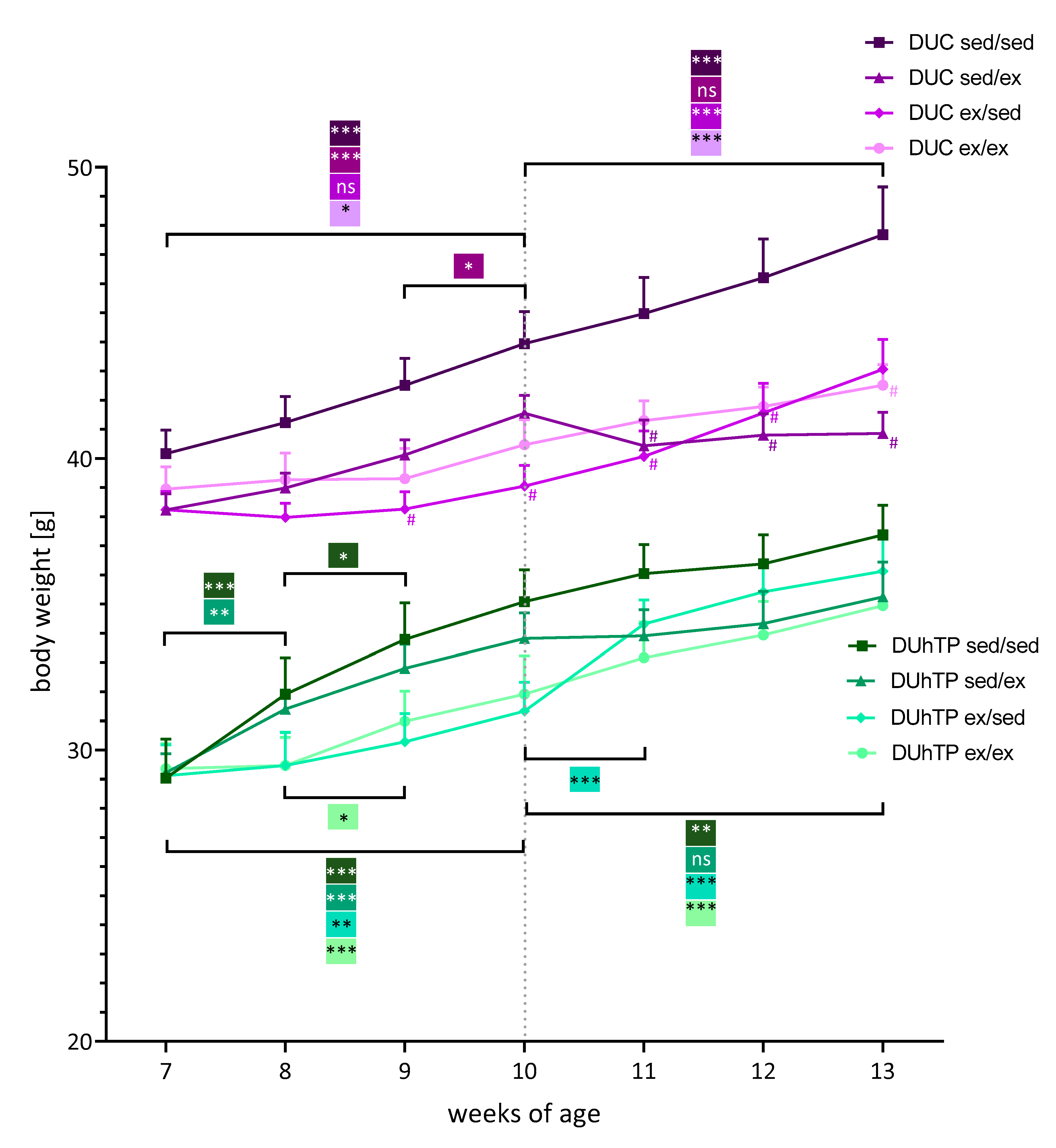
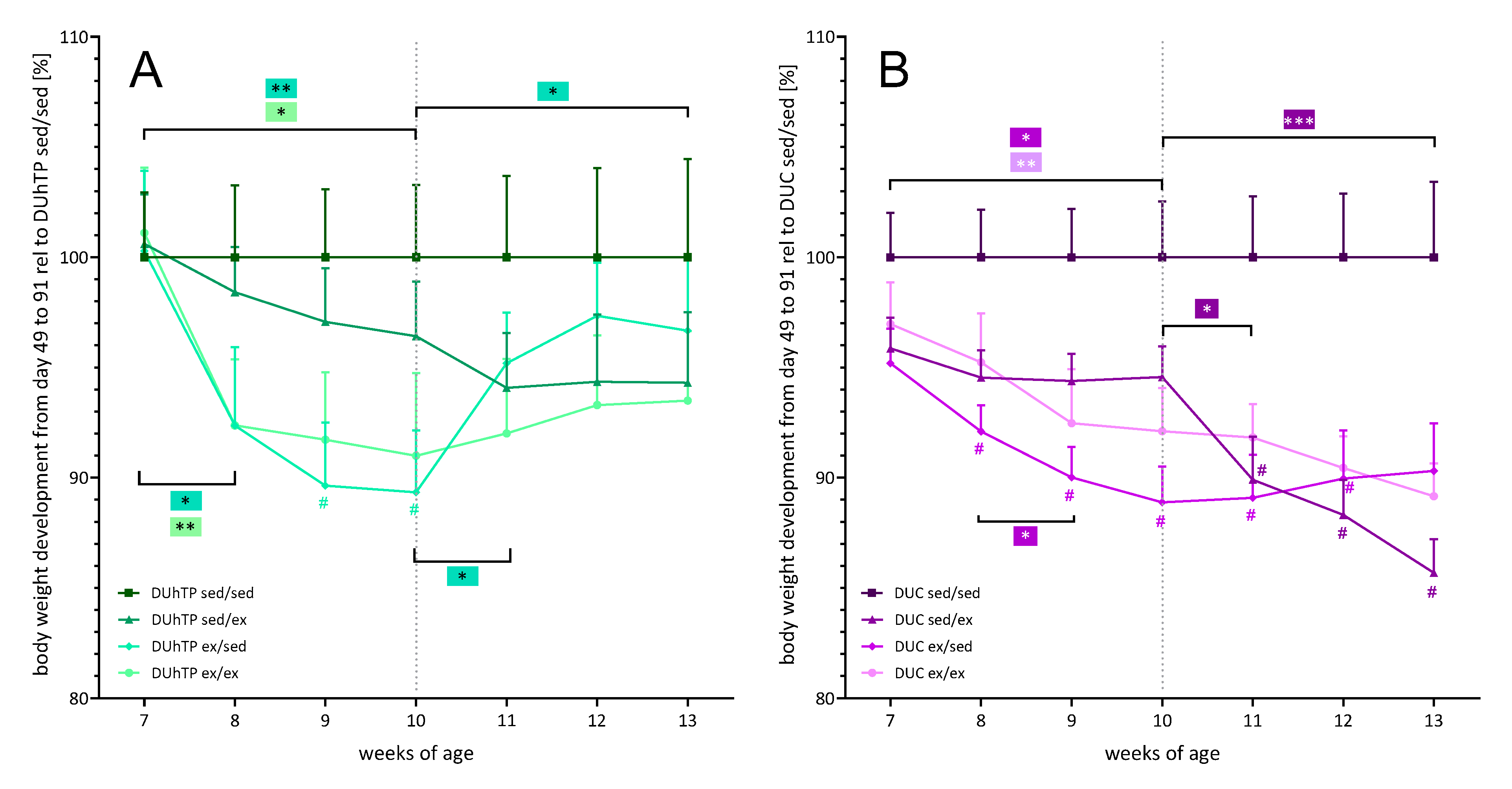
3.2. Effects of Three Different Exercise Programs on Body Weight, Serum Triglyceride Levels, and Fat Deposition
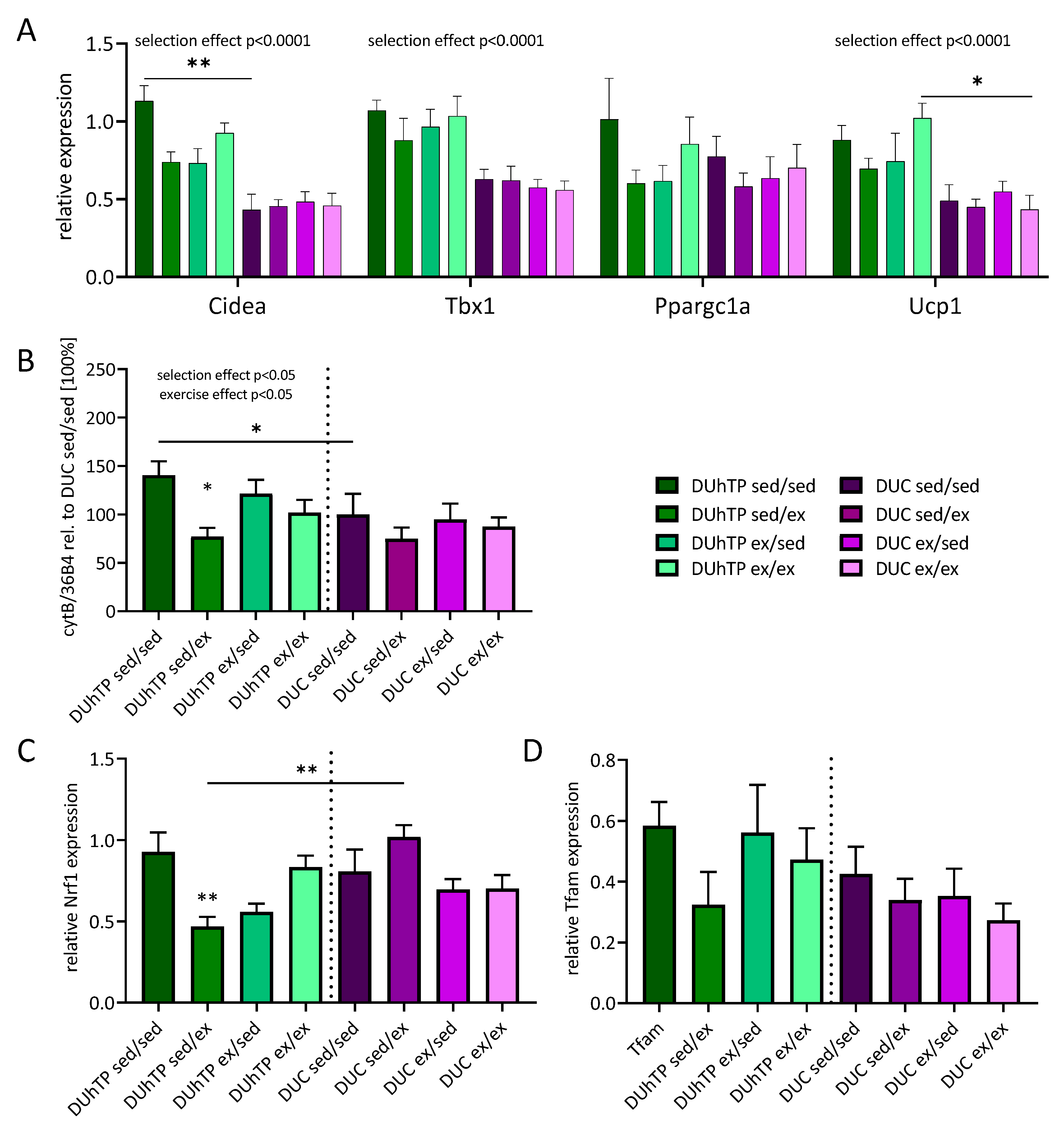
3.3. Effects of Three Different Exercise Programs on the Expression of Browning Markers and Mitochondrial Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cao, L.; Choi, E.Y.; Liu, X.; Martin, A.; Wang, C.; Xu, X.; During, M.J. White to brown fat phenotypic switch induced by genetic and environmental activation of a hypothalamic-adipocyte axis. Cell Metab. 2011, 14, 324–338. [Google Scholar] [CrossRef] [PubMed]
- Bostrom, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Bostrom, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.I.; Middelbeek, R.J.; Goodyear, L.J. Exercise Effects on White Adipose Tissue: Beiging and Metabolic Adaptations. Diabetes 2015, 64, 2361–2368. [Google Scholar] [CrossRef]
- Puigserver, P.; Spiegelman, B.M. Peroxisome proliferator-activated receptor-gamma coactivator 1 alpha (PGC-1 alpha): Transcriptional coactivator and metabolic regulator. Endocr. Rev. 2003, 24, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Little, J.P.; Safdar, A.; Bishop, D.; Tarnopolsky, M.A.; Gibala, M.J. An acute bout of high-intensity interval training increases the nuclear abundance of PGC-1α and activates mitochondrial biogenesis in human skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, 1303–1310. [Google Scholar] [CrossRef]
- Trevellin, E.; Scorzeto, M.; Olivieri, M.; Granzotto, M.; Valerio, A.; Tedesco, L.; Fabris, R.; Serra, R.; Quarta, M.; Reggiani, C.; et al. Exercise training induces mitochondrial biogenesis and glucose uptake in subcutaneous adipose tissue through eNOS-dependent mechanisms. Diabetes 2014, 63, 2800–2811. [Google Scholar] [CrossRef]
- Sutherland, L.N.; Bomhof, M.R.; Capozzi, L.C.; Basaraba, S.A.; Wright, D.C. Exercise and adrenaline increase PGC-1{alpha} mRNA expression in rat adipose tissue. J. Physiol. 2009, 587, 1607–1617. [Google Scholar] [CrossRef]
- Brenmoehl, J.; Ohde, D.; Walz, C.; Schultz, J.; Tuchscherer, A.; Rieder, F.; Renne, U.; Hoeflich, A. Dynamics of Fat Mass in DUhTP Mice Selected for Running Performance—Fat Mobilization in a Walk. Obes. Facts 2015, 8, 373–385. [Google Scholar] [CrossRef]
- Lehnig, A.C.; Dewal, R.S.; Baer, L.A.; Kitching, K.M.; Munoz, V.R.; Arts, P.J.; Sindeldecker, D.A.; May, F.J.; Lauritzen, H.; Goodyear, L.J.; et al. Exercise Training Induces Depot-Specific Adaptations to White and Brown Adipose Tissue. iScience 2019, 11, 425–439. [Google Scholar] [CrossRef]
- Tanaka, T.; Nishimura, A.; Nishiyama, K.; Goto, T.; Numaga-Tomita, T.; Nishida, M. Mitochondrial dynamics in exercise physiology. Pflug. Arch. 2019. [Google Scholar] [CrossRef]
- Hyde, B.B.; Twig, G.; Shirihai, O.S. Organellar vs cellular control of mitochondrial dynamics. Semin. Cell Dev. Biol. 2010, 21, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Van der Bliek, A.M.; Shen, Q.; Kawajiri, S. Mechanisms of mitochondrial fission and fusion. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Gan, Z.; Fu, T.; Kelly, D.P.; Vega, R.B. Skeletal muscle mitochondrial remodeling in exercise and diseases. Cell Res. 2018, 28, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C. Mitochondrial fusion and fission in mammals. Annu Rev. Cell Dev. Biol. 2006, 22, 79–99. [Google Scholar] [CrossRef]
- Chen, H.; Chan, D.C. Emerging functions of mammalian mitochondrial fusion and fission. Hum. Mol. Genet. 2005, 14, R283–R289. [Google Scholar] [CrossRef]
- Martin, O.J.; Lai, L.; Soundarapandian, M.M.; Leone, T.C.; Zorzano, A.; Keller, M.P.; Attie, A.D.; Muoio, D.M.; Kelly, D.P. A Role for PGC-1 Coactivators in the Control of Mitochondrial Dynamics during Postnatal Cardiac Growth. Circ. Res. 2014, 114, 626–636. [Google Scholar] [CrossRef]
- Vainshtein, A.; Tryon, L.D.; Pauly, M.; Hood, D.A. Role of PGC-1α during acute exercise-induced autophagy and mitophagy in skeletal muscle. Am. J. Physiol. Cell Physiol. 2015, 308, C710–C719. [Google Scholar] [CrossRef]
- Drake, J.C.; Wilson, R.J.; Yan, Z. Molecular mechanisms for mitochondrial adaptation to exercise training in skeletal muscle. FASEB J. 2016, 30, 13–22. [Google Scholar] [CrossRef]
- Ding, H.; Jiang, N.; Liu, H.; Liu, X.; Liu, D.; Zhao, F.; Wen, L.; Liu, S.; Ji, L.L.; Zhang, Y. Response of mitochondrial fusion and fission protein gene expression to exercise in rat skeletal muscle. Biochim. Biophys. Acta 2010, 1800, 250–256. [Google Scholar] [CrossRef]
- Marton, O.; Koltai, E.; Takeda, M.; Koch, L.G.; Britton, S.L.; Davies, K.J.A.; Boldogh, I.; Radak, Z. Mitochondrial biogenesis-associated factors underlie the magnitude of response to aerobic endurance training in rats. Pflug. Arch. Eur. J. Physiol. 2015, 467, 779–788. [Google Scholar] [CrossRef]
- Pagano, A.F.; Py, G.; Bernardi, H.; Candau, R.B.; Sanchez, A.M. Autophagy and protein turnover signaling in slow-twitch muscle during exercise. Med. Sci. Sports Exerc. 2014, 46, 1314–1325. [Google Scholar] [CrossRef] [PubMed]
- Trewin, A.J.; Berry, B.J.; Wojtovich, A.P. Exercise and Mitochondrial Dynamics: Keeping in Shape with ROS and AMPK. Antioxidant (Basel) 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.M.; Zhou, Z.; Cohn, W.; Norheim, F.; Lin, A.J.; Kalajian, N.; Strumwasser, A.R.; Cory, K.; Whitney, K.; Ho, T.; et al. The impact of exercise on mitochondrial dynamics and the role of Drp1 in exercise performance and training adaptations in skeletal muscle. Mol. Metab. 2019, 21, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Brenmoehl, J.; Walz, C.; Renne, U.; Ponsuksili, S.; Wolf, C.; Langhammer, M.; Schwerin, M.; Hoeflich, A. Metabolic adaptations in the liver of born long-distance running mice. Med. Sci. Sports Exerc. 2013, 45, 841–850. [Google Scholar] [CrossRef]
- Falkenberg, H.; Langhammer, M.; Renne, U. Comparison of biochemical blood traits after long-term selection on high or low locomotory activity in mice. Arch. Tierz. 2000, 43, 513–522. [Google Scholar] [CrossRef]
- Brenmoehl, J.; Walz, C.; Spitschak, M.; Wirthgen, E.; Walz, M.; Langhammer, M.; Tuchscherer, A.; Naumann, R.; Hoeflich, A. Partial phenotype conversion and differential trait response to conditions of husbandry in mice. J. Comp. Physiol. B 2018, 188, 527–539. [Google Scholar] [CrossRef]
- Brenmoehl, J.; Ohde, D.; Albrecht, E.; Walz, C.; Tuchscherer, A.; Hoeflich, A. Browning of subcutaneous fat and higher surface temperature in response to phenotype selection for advanced endurance exercise performance in male DUhTP mice. J. Comp. Physiol. B 2017, 187, 361–373. [Google Scholar] [CrossRef]
- Ohde, D.; Brenmoehl, J.; Walz, C.; Tuchscherer, A.; Wirthgen, E.; Hoeflich, A. Comparative analysis of hepatic miRNA levels in male marathon mice reveals a link between obesity and endurance exercise capacities. J. Comp. Physiol. B 2016, 186, 1067–1078. [Google Scholar] [CrossRef]
- Ballester, M.; Cordón, R.; Folch, J.M. DAG expression: High-throughput gene expression analysis of real-time PCR data using standard curves for relative quantification. PLoS ONE 2013, 8, e80385. [Google Scholar] [CrossRef]
- Hofmeister-Brix, A.; Kollmann, K.; Langer, S.; Schultz, J.; Lenzen, S.; Baltrusch, S. Identification of the ubiquitin-like domain of midnolin as a new glucokinase interaction partner. J. Biol. Chem. 2013, 288, 35824–35839. [Google Scholar] [CrossRef]
- Barbatelli, G.; Murano, I.; Madsen, L.; Hao, Q.; Jimenez, M.; Kristiansen, K.; Giacobino, J.P.; De Matteis, R.; Cinti, S. The emergence of cold-induced brown adipocytes in mouse white fat depots is determined predominantly by white to brown adipocyte transdifferentiation. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1244–E1253. [Google Scholar] [CrossRef] [PubMed]
- Dietl, G.; Langhammer, M.; Renne, U. Model simulations for genetic random drift in the outbred strain Fzt:DU. Arch. Tierz. 2004, 47, 595–604. [Google Scholar] [CrossRef]
- Davidson, S.R.; Burnett, M.; Hoffman-Goetz, L. Training effects in mice after long-term voluntary exercise. Med. Sci. Sports Exerc. 2006, 38, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Sertie, R.A.L.; Curi, R.; Oliveira, A.C.; Andreotti, S.; Caminhotto, R.O.; de Lima, T.M.; Proenca, A.R.G.; Reis, G.B.; Lima, F.B. The mechanisms involved in the increased adiposity induced by interruption of regular physical exercise practice. Life Sci. 2019, 222, 103–111. [Google Scholar] [CrossRef]
- Tokuyama, K.; Saito, M.; Okuda, H. Effects of wheel running on food intake and weight gain of male and female rats. Physiol. Behav. 1982, 28, 899–903. [Google Scholar] [CrossRef]
- Ziegler, A.K.; Damgaard, A.; Mackey, A.L.; Schjerling, P.; Magnusson, P.; Olesen, A.T.; Kjaer, M.; Scheele, C. An anti-inflammatory phenotype in visceral adipose tissue of old lean mice, augmented by exercise. Sci Rep. 2019, 9, 12069. [Google Scholar] [CrossRef]
- Ruegsegger, G.N.; Company, J.M.; Toedebusch, R.G.; Roberts, C.K.; Roberts, M.D.; Booth, F.W. Rapid Alterations in Perirenal Adipose Tissue Transcriptomic Networks with Cessation of Voluntary Running. PLoS ONE 2015, 10, e0145229. [Google Scholar] [CrossRef]
- Ruegsegger, G.N.; Sevage, J.A.; Childs, T.E.; Grigsby, K.B.; Booth, F.W. 5-Aminoimidazole-4-carboxamide ribonucleotide prevents fat gain following the cessation of voluntary physical activity. Exp. Physiol. 2017, 102, 1474–1485. [Google Scholar] [CrossRef]
- Wu, M.V.; Bikopoulos, G.; Hung, S.; Ceddia, R.B. Thermogenic capacity is antagonistically regulated in classical brown and white subcutaneous fat depots by high fat diet and endurance training in rats: Impact on whole-body energy expenditure. J. Biol. Chem. 2014, 289, 34129–34140. [Google Scholar] [CrossRef]
- Aldiss, P.; Betts, J.; Sale, C.; Pope, M.; Budge, H.; Symonds, M.E. Exercise-induced ‘browning’ of adipose tissues. Metabolism 2018, 81, 63–70. [Google Scholar] [CrossRef]
- Templeman, N.M.; Schutz, H.; Garland, T., Jr.; McClelland, G.B. Do mice bred selectively for high locomotor activity have a greater reliance on lipids to power submaximal aerobic exercise? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R101–R111. [Google Scholar] [CrossRef][Green Version]
- Wu, J.; Bostrom, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef]
- Jash, S.; Banerjee, S.; Lee, M.-J.; Farmer, S.R.; Puri, V. CIDEA Transcriptionally Regulates UCP1 for Britening and Thermogenesis in Human Fat Cells. iScience 2019, 20, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Garnier, A.; Fortin, D.; Zoll, J.; N’Guessan, B.; Mettauer, B.; Lampert, E.; Veksler, V.; Ventura-Clapier, R. Coordinated changes in mitochondrial function and biogenesis in healthy and diseased human skeletal muscle. FASEB J. 2005, 19, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef]
- Virbasius, J.V.; Scarpulla, R.C. Activation of the human mitochondrial transcription factor A gene by nuclear respiratory factors: A potential regulatory link between nuclear and mitochondrial gene expression in organelle biogenesis. Proc. Natl. Acad. Sci. USA 1994, 91, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.C.; Han, D.H.; Garcia-Roves, P.M.; Geiger, P.C.; Jones, T.E.; Holloszy, J.O. Exercise-induced mitochondrial biogenesis begins before the increase in muscle PGC-1alpha expression. J. Biol. Chem. 2007, 282, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.R.; Blackstone, C. Dynamic regulation of mitochondrial fission through modification of the dynamin-related protein Drp1. Ann. N. Y. Acad. Sci. 2010, 1201, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.B.; Bush, Z.; McGinnis, G.R.; Rowe, G.C. Adult skeletal muscle deletion of Mitofusin 1 and 2 impedes exercise performance and training capacity. J. Appl. Physiol. (1985) 2019, 126, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Civiletto, G.; Varanita, T.; Cerutti, R.; Gorletta, T.; Barbaro, S.; Marchet, S.; Lamperti, C.; Viscomi, C.; Scorrano, L.; Zeviani, M. Opa1 overexpression ameliorates the phenotype of two mitochondrial disease mouse models. Cell Metab. 2015, 21, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Lally, J.S.; Herbst, E.A.; Matravadia, S.; Maher, A.C.; Perry, C.G.; Ventura-Clapier, R.; Holloway, G.P. Over-expressing mitofusin-2 in healthy mature mammalian skeletal muscle does not alter mitochondrial bioenergetics. PLoS ONE 2013, 8, e55660. [Google Scholar] [CrossRef] [PubMed]
- Touvier, T.; De Palma, C.; Rigamonti, E.; Scagliola, A.; Incerti, E.; Mazelin, L.; Thomas, J.L.; D’Antonio, M.; Politi, L.; Schaeffer, L.; et al. Muscle-specific Drp1 overexpression impairs skeletal muscle growth via translational attenuation. Cell Death Dis. 2015, 6, e1663. [Google Scholar] [CrossRef] [PubMed]
- Liesa, M.; Van der Bliek, A.; Shirihai, O.S. To Fis or not to Fuse? This is the question! EMBO J. 2019, 38. [Google Scholar] [CrossRef]
- Yu, R.; Jin, S.B.; Lendahl, U.; Nister, M.; Zhao, J. Human Fis1 regulates mitochondrial dynamics through inhibition of the fusion machinery. EMBO J. 2019, 38. [Google Scholar] [CrossRef]
- Cereghetti, G.M.; Stangherlin, A.; de Brito, O.M.; Chang, C.R.; Blackstone, C.; Bernardi, P.; Scorrano, L. Dephosphorylation by calcineurin regulates translocation of Drp1 to mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 15803–15808. [Google Scholar] [CrossRef]
- Cartoni, R.; Leger, B.; Hock, M.B.; Praz, M.; Crettenand, A.; Pich, S.; Ziltener, J.L.; Luthi, F.; Deriaz, O.; Zorzano, A.; et al. Mitofusins 1/2 and ERRalpha expression are increased in human skeletal muscle after physical exercise. J. Physiol. 2005, 567, 349–358. [Google Scholar] [CrossRef]
- Rodríguez, A.; Becerril, S.; Ezquerro, S.; Méndez-Giménez, L.; Frühbeck, G. Crosstalk between adipokines and myokines in fat browning. Acta Physiol. (Oxf.) 2017, 219, 362–381. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward 5′-3′ | Reverse 5′-3′ |
|---|---|---|
| Nrf1 | ACAGATAGTCCTGTCTGGGGAAA | TGGTACATGCTCACAGGGATCT |
| Tfam | AAGACCTCGTTCAGCATATAACATT | TTTTCCAAGCCTCATTTACAAGC |
| Atp5b | CGTGAGGGCAATGATTTATACCAT | TCCTGGTCTCTGAAGTATTCAGCAA |
| Cyc | ATAGGGGCATGTCACCTCAAAC | GTGGTTAGCCATGACCTGAAAG |
| Cox4i1 | TGGGACTATGACAAGAATGAGTGG | TTAGCATGGACCATTGGATACGG |
| 18S | CTGCCCTATCAACTTTCGATGGTAG | CCGTTTCTCAGGCTCCCTCTC |
| Cyb | CTTCGCTTTCCACTTCATCTTACC | TTGGGTTGTTTGATCCTGTTTCG |
| 36B4 | AGGATATGGGATTCGGTCTCTTC | TCATCCTGCTTAAGTGAACAAACT |
| Ucp1 | GGCCTCTACGACTCAGTCCA | TAAGCCGGCTGAGATCTTGT |
| Pgargc1a | CTGCATGAGTGTGTGCTGTG | GGAAGATCTGGGCAAAGAGG |
| Tbx1 | GGCAGGCAGACGAATGTTC | TTGTCATCTACGGGCACAAAG |
| Cidea | GTACTCGGTGTCCTACGACATC | TCATCTGTGCAGCATAGGACATA |
| Dnm1 | CGGTTAGACAGTGCACCAAG | GGATGTGGGTGGTCACAAT |
| Fis1 | GCCCCTGCTACTGGACCAT | CCCTGAAAGCCTCACACTAAGG |
| Mfn1 | AGCCAAGGAAGTTCTCAACTC | GCTCTGATAGTGTGCTGTTCA |
| Mfn2 | GCCAGCTTCCTTGAAGACAC | GCAGAACTTTGTCCCAGAGC |
| Opa1 | TGACAAACTTAAGGAGGCTGTG | CATTGTGCTGAATAACCCTCAA |
| Rpl19 | CAATGCCAACTCCCGTCAGC | TCTTGGATTCCCGGTATCTC |
| Pgk1 | CAGTCTAGAGCTCCTGGAAGGT | GCCACTAGCTGAATCTTGCG |
| Actb | TGACAGGATGCAGAAGGAGA | CGCTCAGGAGGAGCAATG |
| Rplp2 | GACGATGATCGGCTCAACAAG | ACCCTGAGCGATGACATCCT |
| Sdha | CAAATTCTCTCTTGGACCTTGTAGT | CCTTAATTGAAGGAACTTTATCTCCA |
| DUhTP | DUC | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Correlation | sed/sed | sed/ex | ex/sed | ex/ex | sed/sed | sed/ex | ex/sed | ex/ex | |
| Ucp1/Cidea | r | 0.754 | 0.785 | 0.935 | 0.48 | 0.968 | 0.664 | 0.865 | 0.793 |
| p | * | * | ** | ns | *** | ns | ** | * | |
| Ppargc1a/Nrf1 | r | 0.375 | 0.379 | −0.266 | 0.334 | 0.098 | −0.237 | −0.444 | 0.057 |
| p | ns | ns | ns | ns | ns | ns | ns | ns | |
| Ppargc1a/Tfam | r | −0.000 | 0.867 | 0.828 | 0.850 | 0.211 | 0.753 | 0.008 | 0.424 |
| p | ns | ** | * | ** | ns | * | ns | ns | |
| Ppargc1a/Cyc | r | −0.102 | 0.733 | 0.836 | 0.843 | 0.275 | 0.501 | 0.210 | 0.021 |
| p | ns | * | * | ** | ns | ns | ns | ns | |
| Ppargc1a/Cytb | r | 0.067 | 0.632 | 0.731 | 0.854 | 0.406 | 0.495 | 0.653 | 0.211 |
| p | ns | ns | * | ** | ns | ns | ns | ns | |
| Ppargc1a/Mfn1 | r | −0.401 | 0.866 | 0.925 | 0.945 | 0.285 | 0.626 | 0.745 | 0.315 |
| p | ns | ** | ** | *** | ns | ns | * | ns | |
| Ppargc1a/Opa1 | r | 0.363 | 0.943 | 0.788 | 0.899 | 0.196 | 0.694 | 0.414 | 0.310 |
| p | ns | *** | * | ** | ns | * | ns | ns | |
| Mfn1/Opa1 | r | 0.464 | 0.963 | 0.938 | 0.936 | 0.899 | 0.990 | 0.830 | 0.914 |
| p | ns | *** | ** | *** | *** | *** | * | ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brenmoehl, J.; Ohde, D.; Walz, C.; Langhammer, M.; Schultz, J.; Hoeflich, A. Analysis of Activity-Dependent Energy Metabolism in Mice Reveals Regulation of Mitochondrial Fission and Fusion mRNA by Voluntary Physical Exercise in Subcutaneous Fat from Male Marathon Mice (DUhTP). Cells 2020, 9, 2697. https://doi.org/10.3390/cells9122697
Brenmoehl J, Ohde D, Walz C, Langhammer M, Schultz J, Hoeflich A. Analysis of Activity-Dependent Energy Metabolism in Mice Reveals Regulation of Mitochondrial Fission and Fusion mRNA by Voluntary Physical Exercise in Subcutaneous Fat from Male Marathon Mice (DUhTP). Cells. 2020; 9(12):2697. https://doi.org/10.3390/cells9122697
Chicago/Turabian StyleBrenmoehl, Julia, Daniela Ohde, Christina Walz, Martina Langhammer, Julia Schultz, and Andreas Hoeflich. 2020. "Analysis of Activity-Dependent Energy Metabolism in Mice Reveals Regulation of Mitochondrial Fission and Fusion mRNA by Voluntary Physical Exercise in Subcutaneous Fat from Male Marathon Mice (DUhTP)" Cells 9, no. 12: 2697. https://doi.org/10.3390/cells9122697
APA StyleBrenmoehl, J., Ohde, D., Walz, C., Langhammer, M., Schultz, J., & Hoeflich, A. (2020). Analysis of Activity-Dependent Energy Metabolism in Mice Reveals Regulation of Mitochondrial Fission and Fusion mRNA by Voluntary Physical Exercise in Subcutaneous Fat from Male Marathon Mice (DUhTP). Cells, 9(12), 2697. https://doi.org/10.3390/cells9122697