Retinoic Acid and Its Derivatives in Skin

 , , , , ,
, , , , ,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Retinoic Acid Receptors and Molecular Mechanism of Their Action
3. Non-Genomic Effects of ATRA
4. Retinoic Acid in the Skin
5. Skin Differentiation
6. Epidermal Barrier
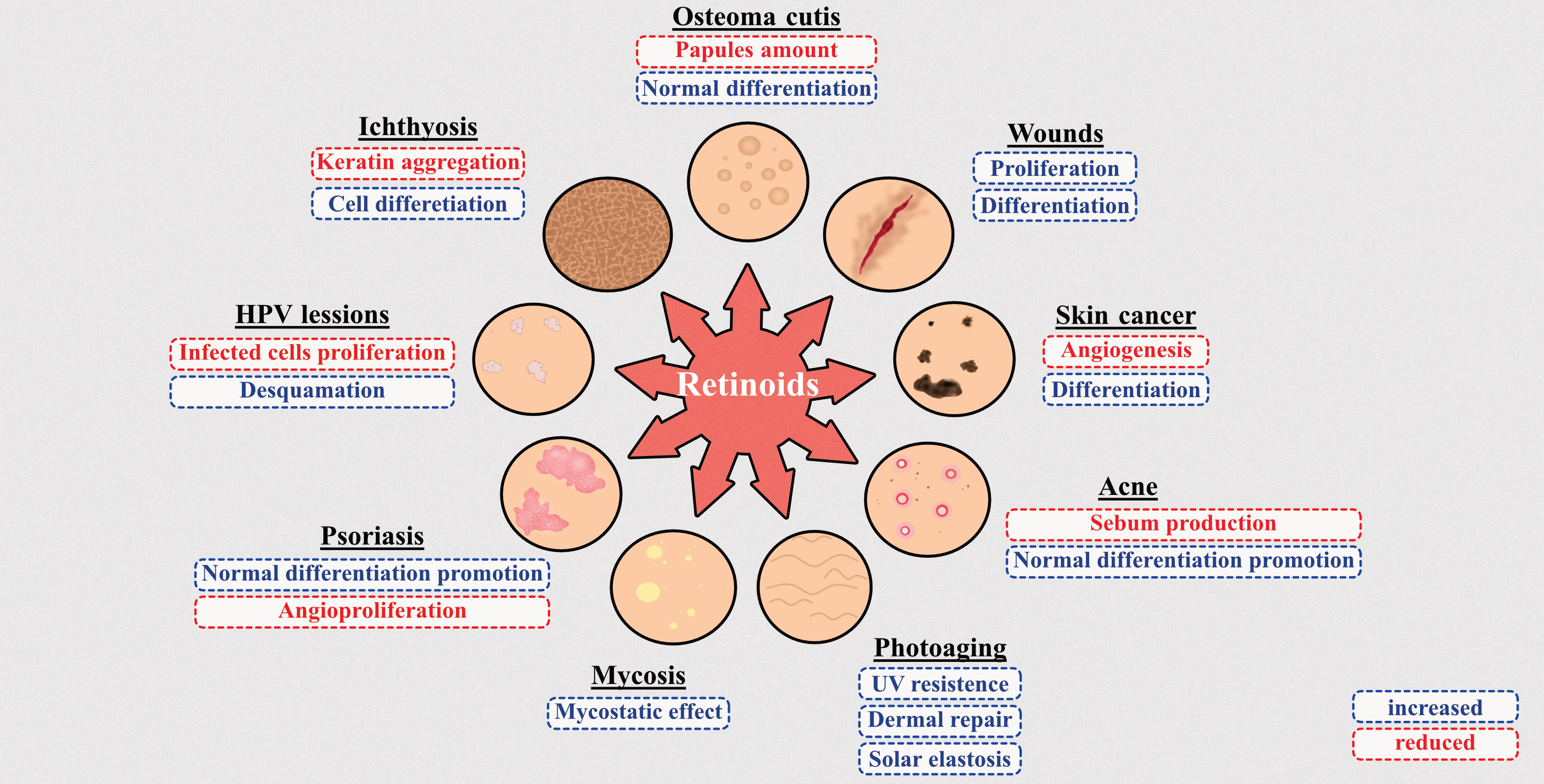
7. New Strategies in ATRA and Other Retinoids-Mediated Therapy of Skin Conditions
8. Limitations of Use
Author Contributions
Funding
Conflicts of Interest
References
- Niederreither, K.; Dollé, P. Retinoic acid in development: Towards an integrated view. Nat. Rev. Genet. 2008, 9, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019. [Google Scholar] [CrossRef] [PubMed]
- Khalil, S.; Bardawil, T.; Stephan, C.; Darwiche, N.; Abbas, O.; Kibbi, A.G.; Nemer, G.; Kurban, M. Retinoids: A Journey from the Molecular Structures and Mechanisms of Action to Clinical Uses in Dermatology and Adverse Effects. J. Dermatol. Treat. 2017, 28, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.C.; Zolfaghari, R. Cytochrome P450s in the Regulation of Cellular Retinoic Acid Metabolism. Annu. Rev. Nutr. 2011, 31, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Stevison, F.; Jing, J.; Tripathy, S.; Isoherranen, N. Role of retinoic acid metabolizing cytochrome P450s, CYP26, in inflammation and cancer. In Advances in Pharmacology; Academic Press: Cambridge, MA, USA, 2015; Volume 74, pp. 373–412. [Google Scholar] [CrossRef]
- Watson, P.J.; Fairall, L.; Schwabe, J.W.R. Nuclear hormone receptor co-repressors: Structure and function. Mol. Cell. Endocrinol. 2012, 348–135, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Chandra, V.; Rastinejad, F. Retinoic Acid Actions Through Mammalian Nuclear Receptors. Chem. Rev. 2014, 114, 233–254. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, O.; Welch, J.S. Retinoic Acid Receptors in Acute Myeloid Leukemia Therapy. Cancers 2019, 11, 1915. [Google Scholar] [CrossRef]
- Leid, M.; Kastner, P.; Chambon, P. Multiplicity generates diversity in the retinoic acid signalling pathways. Trends Biochem. Sci. 1992, 17, 427–433. [Google Scholar] [CrossRef]
- Schenk, T.; Stengel, S.; Zelent, A. Unlocking the potential of retinoic acid in anticancer therapy. Br. J. Cancer 2014, 111, 2039–2045. [Google Scholar] [CrossRef]
- Bushue, N.; Wan, Y.-J.Y. Retinoid pathway and cancer therapeutics. Adv. Drug Deliv. Rev. 2010, 62, 1285–1298. [Google Scholar] [CrossRef]
- Aging | Cellular Retinoic Acid Binding Protein-II Expression and Its Potential Role in Skin Aging. Available online: https://www.aging-us.com/article/101813 (accessed on 27 November 2020).
- Redfern, C.P.; Todd, C. Retinoic acid receptor expression in human skin keratinocytes and dermal fibroblasts in vitro. J. Cell Sci. 1992, 102, 113–121. [Google Scholar] [PubMed]
- Törmä, H. Regulation of keratin expression by retinoids. Dermatoendocrinology 2011, 3, 136–140. [Google Scholar] [CrossRef]
- Idres, N.; Marill, J.; Flexor, M.A.; Chabot, G.G. Activation of retinoic acid receptor-dependent transcription by all-trans-retinoic acid metabolites and isomers. J. Biol. Chem. 2002, 277, 31491–31498. [Google Scholar] [CrossRef] [PubMed]
- Di Masi, A.; Leboffe, L.; De Marinis, E.; Pagano, F.; Cicconi, L.; Rochette-Egly, C.; Lo-Coco, F.; Ascenzi, P.; Nervi, C. Retinoic acid receptors: From molecular mechanisms to cancer therapy. Mol. Asp. Med. 2015, 41, 1–115. [Google Scholar] [CrossRef] [PubMed]
- Das, B.C.; Thapa, P.; Karki, R.; Das, S.; Mahapatra, S.; Liu, T.-C.; Torregroza, I.; Wallace, D.P.; Kambhampati, S.; Van Veldhuizen, P.; et al. Retinoic acid signaling pathways in development and diseases. Bioorg. Med. Chem. 2014, 22, 673–683. [Google Scholar] [CrossRef]
- Piskunov, A.; Rochette-Egly, C. A retinoic acid receptor RARα pool present in membrane lipid rafts forms complexes with G protein αQ to activate p38MAPK. Oncogene 2012, 31, 3333–3345. [Google Scholar] [CrossRef]
- Masiá, S.; Alvarez, S.; de Lera, A.R.; Barettino, D. Rapid, nongenomic actions of retinoic acid on phosphatidylinositol-3-kinase signaling pathway mediated by the retinoic acid receptor. Mol. Endocrinol. Baltim. Md. 2007, 21, 2391–2402. [Google Scholar] [CrossRef]
- Kirchmeyer, M.; Koufany, M.; Sebillaud, S.; Netter, P.; Jouzeau, J.-Y.; Bianchi, A. All-trans retinoic acid suppresses interleukin-6 expression in interleukin-1-stimulated synovial fibroblasts by inhibition of ERK1/2 pathway independently of RAR activation. Arthritis Res. Ther. 2008, 10, R141. [Google Scholar] [CrossRef]
- Tkatch, L.S.; Rubin, K.A.; Ziegler, S.F.; Tweardy, D.J. Modulation of human G-CSF receptor mRNA and protein in normal and leukemic myeloid cells by G-CSF and retinoic acid. J. Leukoc. Biol. 1995, 57, 964–971. [Google Scholar] [CrossRef]
- De Gentile, A.; Toubert, M.E.; Dubois, C.; Krawice, I.; Schlageter, M.H.; Balitrand, N.; Castaigne, S.; Degos, L.; Rain, J.D.; Najean, Y. Induction of high-affinity GM-CSF receptors during all-trans retinoic acid treatment of acute promyelocytic leukemia. Leukemia 1994, 8, 1758–1762. [Google Scholar]
- Glasow, A.; Prodromou, N.; Xu, K.; von Lindern, M.; Zelent, A. Retinoids and myelomonocytic growth factors cooperatively activate RARA and induce human myeloid leukemia cell differentiation via MAP kinase pathways. Blood 2005, 105, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Coombs, C.C.; Tavakkoli, M.; Tallman, M.S. Acute promyelocytic leukemia: Where did we start, where are we now, and the future. Blood Cancer J. 2015, 5, e304. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-D.; Stojadinovic, O.; Krzyzanowska, A.; Vouthounis, C.; Blumenberg, M.; Tomic-Canic, M. Retinoid-Responsive Transcriptional Changes in Epidermal Keratinocytes. J. Cell. Physiol. 2009, 220, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Castleberry, S.A.; Hammond, P.T.; Quadir, M.A. Nano-Fibular Nanoparticle Polymer-Drug Conjugate for Sustained Dermal Delivery of Retinoids. U.S. Patent 0185513 A1, 5 July 2018. [Google Scholar]
- Orfanos, C.E.; Zouboulis, C.C.; Almond-Roesler, B.; Geilen, C.C. Current use and future potential role of retinoids in dermatology. Drugs 1997, 53, 358–388. [Google Scholar] [CrossRef]
- Njar, V.C.O.; Gediya, L.; Purushottamachar, P.; Chopra, P.; Vasaitis, T.S.; Khandelwal, A.; Mehta, J.; Huynh, C.; Belosay, A.; Patel, J. Retinoic acid metabolism blocking agents (RAMBAs) for treatment of cancer and dermatological diseases. Bioorg. Med. Chem. 2006, 14, 4323–4340. [Google Scholar] [CrossRef]
- Purushottamachar, P.; Patel, J.B.; Gediya, L.K.; Clement, O.O.; Njar, V.C.O. First chemical feature-based pharmacophore modeling of potent retinoidal retinoic acid metabolism blocking agents (RAMBAs): Identification of novel RAMBA scaffolds. Eur. J. Med. Chem. 2012, 47, 412–423. [Google Scholar] [CrossRef]
- Verfaille, C.J.; Borgers, M.; van Steensel, M.A.M. Retinoic acid metabolism blocking agents (RAMBAs): A new paradigm in the treatment of hyperkeratotic disorders. J. Dtsch. Dermatol. Ges. J. Ger. Soc. Dermatol. JDDG 2008, 6, 355–364. [Google Scholar] [CrossRef]
- Ketoconazole Inhibits the in Vitro and in Vivo Metabolism of All-Trans-Retinoic Acid. | Journal of Pharmacology and Experimental Therapeutics. Available online: https://jpet.aspetjournals.org/content/245/2/718 (accessed on 27 November 2020).
- Vijjan, V.; Dubey, D. New therapeutic targets in the treatment of prostate cancer. Indian J. Urol. IJU J. Urol. Soc. India 2007, 23, 61–66. [Google Scholar] [CrossRef]
- Vahlquist, A.; Blockhuys, S.; Steijlen, P.; van Rossem, K.; Didona, B.; Blanco, D.; Traupe, H. Oral liarozole in the treatment of patients with moderate/severe lamellar ichthyosis: Results of a randomized, double-blind, multinational, placebo-controlled phase II/III trial. Br. J. Dermatol. 2014, 170, 173–181. [Google Scholar] [CrossRef]
- Fuchs, E.; Green, H. Regulation of terminal differentiation of cultured human keratinocytes by vitamin A: Cell. Cell 1981, 25, 617–625. [Google Scholar] [CrossRef]
- Saitou, M.; Sugai, S.; Tanaka, T.; Shimouchi, K.; Fuchs, E.; Narumiya, S.; Kakizuka, A. Inhibition of skin development by targeted expression of a dominant-negative retinoic acid receptor. Nature 1995, 374, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Villarroya, F.; Giralt, M.; Iglesias, R. Retinoids and adipose tissues: Metabolism, cell differentiation and gene expression. Int. J. Obes. 1999, 23, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Fu, X.; Liang, X.; Deavila, J.M.; Wang, Z.; Zhao, L.; Tian, Q.; Zhao, J.; Gomez, N.A.; Trombetta, S.C.; et al. Retinoic acid induces white adipose tissue browning by increasing adipose vascularity and inducing beige adipogenesis of PDGFRα+ adipose progenitors. Cell Discov. 2017, 3, 17036. [Google Scholar] [CrossRef] [PubMed]
- Murphrey, M.B.; Miao, J.H.; Zito, P.M. Histology, Stratum Corneum. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Lee, E.-S.; Ahn, Y.; Bae, I.-H.; Min, D.; Park, N.H.; Jung, W.; Kim, S.-H.; Hong, Y.D.; Park, W.S.; Lee, C.S. Synthetic Retinoid Seletinoid G Improves Skin Barrier Function through Wound Healing and Collagen Realignment in Human Skin Equivalents. Int. J. Mol. Sci. 2020, 21, 3198. [Google Scholar] [CrossRef] [PubMed]
- Kong, R.; Cui, Y.; Fisher, G.J.; Wang, X.; Chen, Y.; Schneider, L.M.; Majmudar, G. A comparative study of the effects of retinol and retinoic acid on histological, molecular, and clinical properties of human skin. J. Cosmet. Dermatol. 2016, 15, 49–57. [Google Scholar] [CrossRef]
- Schurer, N.Y.; Elias, P.M. The biochemistry and function of stratum corneum lipids. Adv. Lipid Res. 1991, 24, 27–56. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Geng, S. All-trans retinoic acid alters the expression of the tight junction proteins Claudin-1 and -4 and epidermal barrier function-associated genes in the epidermis. Int. J. Mol. Med. 2019, 43, 1789–1805. [Google Scholar] [CrossRef]
- Cohen, A.D.; Chetov, T.; Cagnano, E.; Naimer, S.; Vardy, D.A. Treatment of multiple miliary osteoma cutis of the face with local application of tretinoin (all-trans-retinoic acid): A case report and review of the literature. J. Dermatol. Treat. 2001, 12, 171–173. [Google Scholar] [CrossRef]
- Kubeyinje, E. Evaluation of the efficacy and safety of 0.05% tretinoin cream in the treatment of plane warts in Arab children. J. Dermatol. Treat. 1996, 7, 21–22. [Google Scholar] [CrossRef]
- Gelmetti, C.; Cerri, D.; Schiuma, A.A.; Menni, S. Treatment of extensive warts with etretinate: A clinical trial in 20 children. Pediatr. Dermatol. 1987, 4, 254–258. [Google Scholar] [CrossRef]
- Eckert, R.L.; Agarwal, C.; Hembree, J.R.; Choo, C.K.; Sizemore, N.; Andreatta-van Leyen, S.; Rorke, E.A. Human cervical cancer. Retinoids, interferon and human papillomavirus. Adv. Exp. Med. Biol. 1995, 375, 31–44. [Google Scholar] [PubMed]
- Handley, J.; Dinsmore, W. Topical Tretinoin in the Treatment of Anogenital Warts. Sex. Transm. Dis. 1992, 19, 181. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Qian, W.; Chen, F.; Jin, Y.; Wang, F.; Lu, X.; Lee, S.R.; Su, D.; Chen, B. ATRA protects skin fibroblasts against UV-induced oxidative damage through inhibition of E3 ligase Hrd1. Mol. Med. Rep. 2019, 20, 2294–2302. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.P.; Kosuge, K.; Yang, M.; Ito, S. NRF2 as a determinant of cellular resistance in retinoic acid cytotoxicity. Free Radic. Biol. Med. 2008, 45, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Niu, X.; Xiao, S.; Ma, H. Retinoic acid ameliorates photoaged skin through RAR-mediated pathway in mice. Mol. Med. Rep. 2017, 16, 6240–6247. [Google Scholar] [CrossRef][Green Version]
- Sumita, J.M.; Miot, H.A.; Soares, J.L.M.; Raminelli, A.C.P.; Pereira, S.M.; Ogawa, M.M.; Picosse, F.R.; Guadanhim, L.R.S.; Enokihara, M.M.S.S.; Leonardi, G.R.; et al. Tretinoin (0.05% cream vs. 5% peel) for photoaging and field cancerization of the forearms: Randomized, evaluator-blinded, clinical trial. J. Eur. Acad. Dermatol. Venereol. JEADV 2018, 32, 1819–1826. [Google Scholar] [CrossRef]
- Campione, E.; Cosio, T.; Lanna, C.; Mazzilli, S.; Ventura, A.; Dika, E.; Gaziano, R.; Dattola, A.; Candi, E.; Bianchi, L. Predictive role of vitamin A serum concentration in psoriatic patients treated with IL-17 inhibitors to prevent skin and systemic fungal infections. J. Pharmacol. Sci. 2020, 144, 52–56. [Google Scholar] [CrossRef]
- Quan, T.; Fisher, G.J. Role of Age-Associated Alterations of the Dermal Extracellular Matrix Microenvironment in Human Skin Aging: A Mini-Review. Gerontology 2015, 61, 427–434. [Google Scholar] [CrossRef]
- Das, R.P.; Jain, A.K.; Ramesh, V. Current concepts in the pathogenesis of psoriasis. Indian J. Dermatol. 2009, 54, 7–12. [Google Scholar] [CrossRef]
- Wang, H.; Wu, C.; Wang, W.; Jing, H. Retinol and vitamin A metabolites accumulate through RBP4 and STRA6 changes in a psoriasis murine model | Nutrition & Metabolism | Full Text. Nutr. Metab. 2020, 17, 5. [Google Scholar] [CrossRef]
- Laursen, K.B.; Kashyap, V.; Scandura, J.; Gudas, L.J. An Alternative Retinoic Acid-responsive Stra6 Promoter Regulated in Response to Retinol Deficiency. J. Biol. Chem. 2015, 290, 4356–4366. [Google Scholar] [CrossRef] [PubMed]
- Laporte, M.; Galand, P.; Fokan, D.; de Graef, C.; Heenen, M. Apoptosis in established and healing psoriasis. Dermatology 2000, 200, 314–316. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-M.; Wang, K.-P.; Ma, J.; Guo Zheng, S. The role of all-trans retinoic acid in the biology of Foxp3+ regulatory T cells. Cell. Mol. Immunol. 2015, 12, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, L.; Orlandi, A.; Campione, E.; Angeloni, C.; Costanzo, A.; Spagnoli, L.G.; Chimenti, S. Topical treatment of basal cell carcinoma with tazarotene: A clinicopathological study on a large series of cases. Br. J. Dermatol. 2004, 151, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Orlandi, A.; Bianchi, L.; Costanzo, A.; Campione, E.; Spagnoli, L.G.; Chimenti, S. Evidence of Increased Apoptosis and Reduced Proliferation in Basal Cell Carcinomas Treated with Tazarotene. J. Investig. Dermatol. 2004, 122, 1037–1041. [Google Scholar] [CrossRef]
- Doldo, E.; Costanza, G.; Agostinelli, S.; Tarquini, C.; Ferlosio, A.; Arcuri, G.; Passeri, D.; Scioli, M.G.; Orlandi, A. Vitamin A, Cancer Treatment and Prevention: The New Role of Cellular Retinol Binding Proteins. BioMed Res. Int. 2015, 2015, 624627. [Google Scholar] [CrossRef]
- Husmann, M.; Hoffmann, B.; Stump, D.G.; Chytil, F.; Pfahl, M. A retinoic acid response element from the rat CRBPI promoter is activated by an RARRXR heterodimer. Biochem. Biophys. Res. Commun. 1992, 187, 1558–1564. [Google Scholar] [CrossRef]
- Kuppumbatti, Y.S.; Bleiweiss, I.J.; Mandeli, J.P.; Waxman, S.; Mira-Y-Lopez, R. Cellular retinol-binding protein expression and breast cancer. J. Natl. Cancer Inst. 2000, 92, 475–480. [Google Scholar] [CrossRef]
- Garattini, E.; Bolis, M.; Garattini, S.K.; Fratelli, M.; Centritto, F.; Paroni, G.; Gianni’, M.; Zanetti, A.; Pagani, A.; Fisher, J.N.; et al. Retinoids and breast cancer: From basic studies to the clinic and back again. Cancer Treat. Rev. 2014, 40, 739–749. [Google Scholar] [CrossRef]
- Kato, Y.; Salumbides, B.C.; Wang, X.-F.; Qian, D.Z.; Williams, S.; Wei, Y.; Sanni, T.B.; Atadja, P.; Pili, R. Antitumor effect of the histone deacetylase inhibitor LAQ824 in combination with 13-cis-retinoic acid in human malignant melanoma. Mol. Cancer Ther. 2007, 6, 70–81. [Google Scholar] [CrossRef]
- Zhao, X.; Graves, C.; Ames, S.J.; Fisher, D.E.; Spanjaard, R.A. Mechanism of Regulation and Suppression of Melanoma Invasiveness by Novel Retinoic Acid Receptor-γ Target Gene Carbohydrate Sulfotransferase 10. Cancer Res. 2009, 69, 5218–5225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Satyamoorthy, K.; Herlyn, M.; Rosdahl, I. All-trans retinoic acid (atRA) differentially induces apoptosis in matched primary and metastatic melanoma cells—A speculation on damage effect of atRA via mitochondrial dysfunction and cell cycle redistribution. Carcinogenesis 2003, 24, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Nagasawa, T.; Nakamura, N.; Takenaga, M.; Mizoguchi, M.; Kawai, S.-I.; Mizushima, Y.; Igarashi, R. Successful treatment of photo-damaged skin of nano-scale atRA particles using a novel transdermal delivery. J. Control. Release Off. J. Control. Release Soc. 2005, 104, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Arkin, M.; Cherkez, S.; Toledano, O. Method of Acne Treatment by Concomitant Topical Administration of Benzoyl Peroxide and Tretinoin. U.S. Patent 15/978,146, 17 January 2019. [Google Scholar]
- Sklar, J.L. Glycolic Acid and Tretinoin Formulation for the Treatment of Acne. U.S. Patent 5,861,432, 19 January 1999. [Google Scholar]
- Erlich, M.; Marco, D.; Neimann, K.; Toledano, O. Compositions Comprising Encapsulated Tretinoin. U.S. Patent 0015366 A1, 17 January 2019. [Google Scholar]
- Kligman, A.M.; Kligman, D.E. Method of Treating Skin Disorders with High-Strength Tretinoin. U.S. Patent 6,008,254, 28 December 1999. [Google Scholar]
- Bhowmick, S.B.; Dolai, S.K.; Panigrahi, L. Microparticles 2010. EU Patent EP2317989A2, 11 May 2011. [Google Scholar]
- Yamaguchi, Y.; Nakamura, N.; Nagasawa, T.; Kitagawa, A.; Matsumoto, K.; Soma, Y.; Matsuda, T.; Mizoguchi, M.; Igarashi, R. Enhanced skin regeneration by nanoegg formulation of all-trans retinoic acid. Die Pharm.-Int. J. Pharm. Sci. 2006, 61, 117–121. [Google Scholar]
- Marks, A.M. Tretinoin in a Gel Vehicle for Acne Treatment. U.S. Patent 4,247,547, 27 January 1981. [Google Scholar]
- Charoenputtakhun, P.; Opanasopit, P.; Rojanarata, T.; Ngawhirunpat, T. All-trans retinoic acid-loaded lipid nanoparticles as a transdermal drug delivery carrier. Pharm. Dev. Technol. 2014, 19, 164–172. [Google Scholar] [CrossRef]
- Yan, A.C. Current concepts in acne management. Adolesc. Med. Clin. 2006, 17, 613–637. [Google Scholar] [CrossRef]
- Pan, J.; Wang, Q.; Tu, P. A Topical Medication of All-Trans Retinoic Acid Reduces Sebum Excretion Rate in Patients With Forehead Acne. Am. J. Ther. 2017, 24, e207–e212. [Google Scholar] [CrossRef]
- Dogra, S.; Sumathy, T.K.; Nayak, C.; Ravichandran, G.; Vaidya, P.P.; Mehta, S.; Mittal, R.; Mane, A.; Charugulla, S.N. Efficacy and safety comparison of combination of 0.04% tretinoin microspheres plus 1% clindamycin versus their monotherapy in patients with acne vulgaris: A phase 3, randomized, double-blind study. J. Dermatol. Treat. 2020, 1–9. [Google Scholar] [CrossRef]
- Mukherjee, S.; Date, A.; Patravale, V.; Korting, H.C.; Roeder, A.; Weindl, G. Retinoids in the treatment of skin aging: An overview of clinical efficacy and safety. Clin. Interv. Aging 2006, 1, 327–348. [Google Scholar] [CrossRef]
- Kamm, J.J. Toxicology, carcinogenicity, and teratogenicity of some orally administered retinoids. J. Am. Acad. Dermatol. 1982, 6, 652–659. [Google Scholar] [CrossRef]
- Panchaud, A.; Csajka, C.; Merlob, P.; Schaefer, C.; Berlin, M.; De Santis, M.; Vial, T.; Ieri, A.; Malm, H.; Eleftheriou, G.; et al. Pregnancy outcome following exposure to topical retinoids: A multicenter prospective study. J. Clin. Pharmacol. 2012, 52, 1844–1851. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szymański, Ł.; Skopek, R.; Palusińska, M.; Schenk, T.; Stengel, S.; Lewicki, S.; Kraj, L.; Kamiński, P.; Zelent, A. Retinoic Acid and Its Derivatives in Skin. Cells 2020, 9, 2660. https://doi.org/10.3390/cells9122660
Szymański Ł, Skopek R, Palusińska M, Schenk T, Stengel S, Lewicki S, Kraj L, Kamiński P, Zelent A. Retinoic Acid and Its Derivatives in Skin. Cells. 2020; 9(12):2660. https://doi.org/10.3390/cells9122660
Chicago/Turabian StyleSzymański, Łukasz, Rafał Skopek, Małgorzata Palusińska, Tino Schenk, Sven Stengel, Sławomir Lewicki, Leszek Kraj, Paweł Kamiński, and Arthur Zelent. 2020. "Retinoic Acid and Its Derivatives in Skin" Cells 9, no. 12: 2660. https://doi.org/10.3390/cells9122660
APA StyleSzymański, Ł., Skopek, R., Palusińska, M., Schenk, T., Stengel, S., Lewicki, S., Kraj, L., Kamiński, P., & Zelent, A. (2020). Retinoic Acid and Its Derivatives in Skin. Cells, 9(12), 2660. https://doi.org/10.3390/cells9122660