MicroRNAs in Several Cutaneous Autoimmune Diseases: Psoriasis, Cutaneous Lupus Erythematosus and Atopic Dermatitis

 ,
,
Abstract
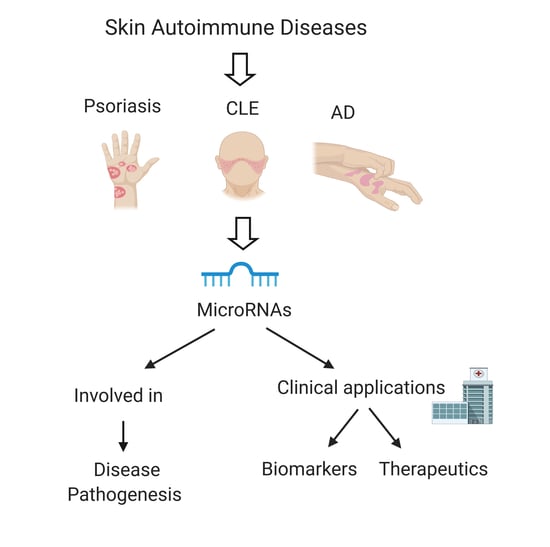
1. Introduction
2. Role of miRNAs in the Skin Pathogenesis of Cutaneous Immune Disorders
2.1. Psoriasis
2.2. Cutaneous Lupus Erythematosus (CLE)
2.3. Atopic Dermatitis (AD)
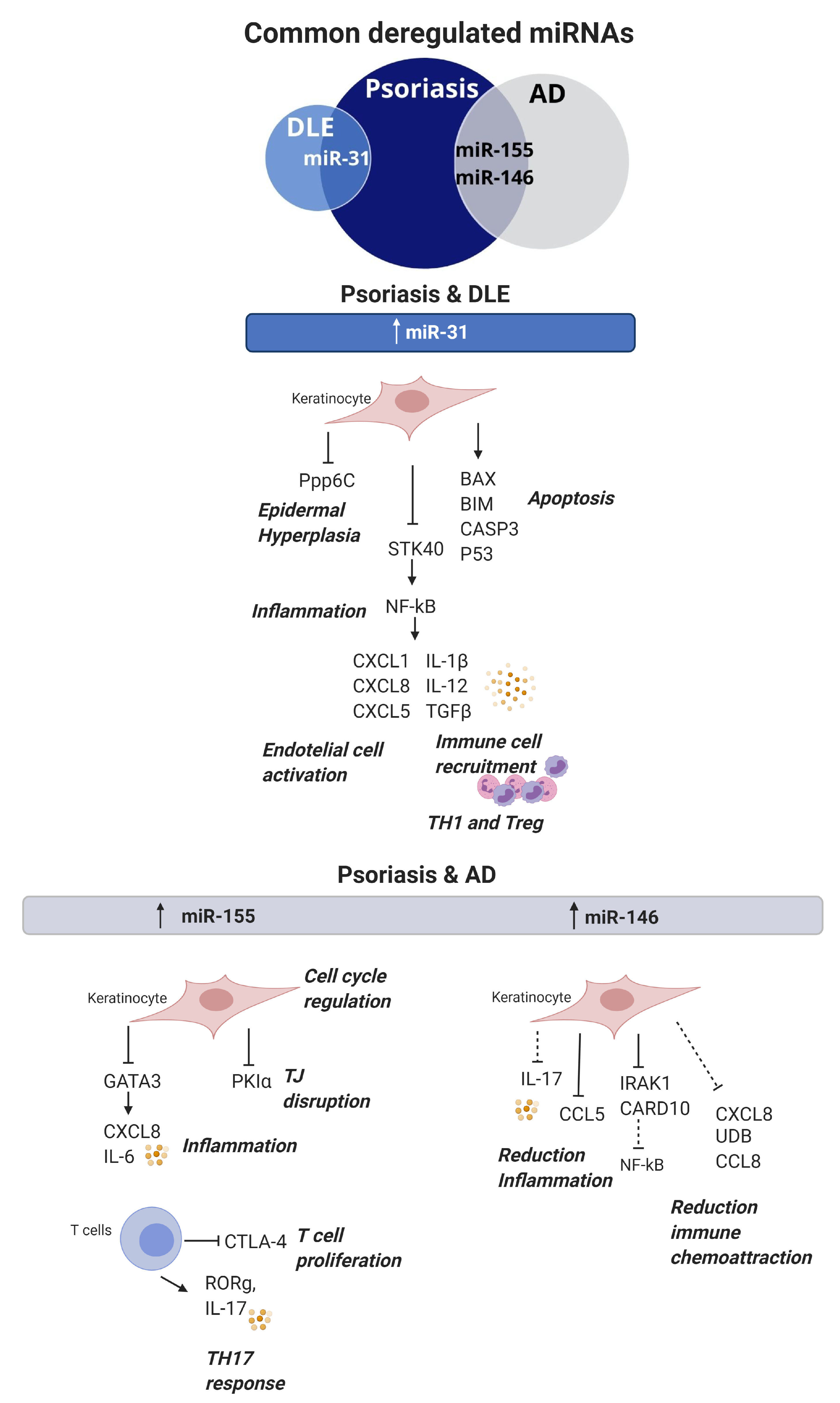
3. Common Deregulated miRNAs in Skin Autoimmune Conditions
4. miRNAs as Potential Biomarkers in Skin Inflammatory Diseases
4.1. miRNAs as Diagnostic Biomarkers
4.2. miRNA as Disease Activity and Severity Biomarkers
4.3. miRNA Levels to Monitor Therapeutic Effects
4.4. Associated Comorbidity Biomarkers
5. Targeting miRNAs to Treat Skin Autoimmune Diseases
5.1. In Vivo Approaches of miRNA Therapy for Skin Autoimmune Diseases
5.2. Clinical Trials Using miRNA as a Therapy for Treating Skin Diseases
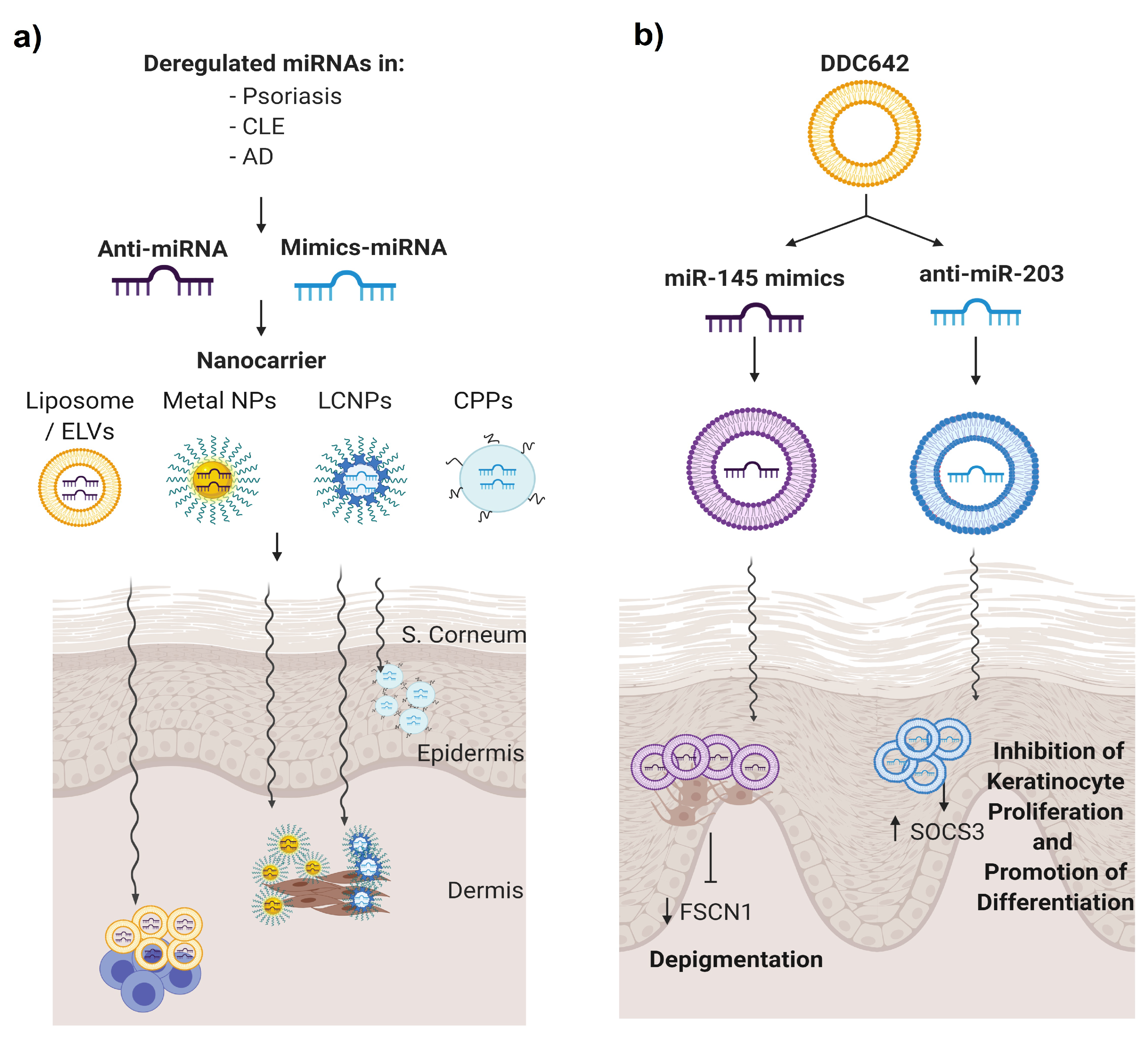
5.3. Topical Nanodelivery of miRNA
6. Conclusions, Limitations and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Baek, D.; Villén, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of microRNAs on protein output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human MicroRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Cell-cycle control of microRNA-mediated translation regulation. Cell Cycle 2008, 7, 1545–1549. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.L.; Sharp, P.A. Function and localization of microRNAs in mammalian cells. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2006; Volume 71, pp. 29–38. [Google Scholar]
- Soifer, H.S.; Rossi, J.J.; Saetrom, P. MicroRNAs in disease and potential therapeutic applications. Mol. Ther. 2007, 15, 2070–2079. [Google Scholar] [CrossRef]
- Andl, T.; Murchison, E.P.; Liu, F.; Zhang, Y.; Yunta-Gonzalez, M.; Tobias, J.W.; Andl, C.D.; Seykora, J.T.; Hannon, G.J.; Millar, S.E. The miRNA-processing enzyme dicer is essential for the morphogenesis and maintenance of hair follicles. Curr. Biol. 2006, 16, 1041–1049. [Google Scholar] [CrossRef]
- Yi, R.; O’Carroll, D.; Pasolli, H.A.; Zhang, Z.; Dietrich, F.S.; Tarakhovsky, A.; Fuchs, E. Morphogenesis in skin is governed by discrete sets of differentially expressed miRNAs. Nat. Genet. 2006, 38, 356–362. [Google Scholar] [CrossRef]
- Yi, R.; Fuchs, E. MicroRNA-mediated control in the skin. Cell Death Differ. 2010, 17, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Neagu, M.; Constantin, C.; Cretoiu, S.M.; Zurac, S. miRNAs in the Diagnosis and Prognosis of Skin Cancer. Front. Cell Dev. Biol. 2020, 8, 71. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Wei, T.; Janson, P.C.; Sääf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A.; et al. MicroRNAs: Novel regulators involved in the pathogenesis of psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef]
- Sonkoly, E.; Janson, P.; Majuri, M.L.; Savinko, T.; Fyhrquist, N.; Eidsmo, L.; Xu, N.; Meisgen, F.; Wei, T.; Bradley, M.; et al. MiR-155 is overexpressed in patients with atopic dermatitis and modulates T-cell proliferative responses by targeting cytotoxic T lymphocyte-associated antigen 4. J. Allergy Clin. Immunol. 2010, 126, 581–920. [Google Scholar] [CrossRef] [PubMed]
- Solé, C.; Domingo, S.; Ferrer, B.; Moliné, T.; Ordi-Ros, J.; Cortés-Hernández, J. MicroRNA Expression Profiling Identifies miR-31 and miR-485-3p as Regulators in the Pathogenesis of Discoid Cutaneous Lupus. J. Investig. Dermatol. 2019, 139, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Parisi, R.; Symmons, D.P.; Griffiths, C.E.; Ashcroft, D.M. Identification and Management of Psoriasis and Associated ComorbidiTy (IMPACT) project team. Global epidemiology of psoriasis: A systematic review of incidence and prevalence. J. Investig. Dermatol. 2013, 133, 377–385. [Google Scholar] [CrossRef]
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef]
- Xu, Y.; Ji, Y.; Lan, X.; Gao, X.; Chen, H.D.; Geng, L. miR-203 contributes to IL-17-induced VEGF secretion by targeting SOCS3 in keratinocytes. Mol. Med. Rep. 2017, 16, 8989–8996. [Google Scholar] [CrossRef]
- Primo, M.N.; Bak, R.O.; Schibler, B.; Mikkelsen, J.G. Regulation of pro-inflammatory cytokines TNFα and IL24 by microRNA-203 in primary keratinocytes. Cytokine 2012, 60, 741–748. [Google Scholar] [CrossRef]
- Wei, T.; Xu, N.; Meisgen, F.; Ståhle, M.; Sonkoly, E.; Pivarcsi, A. Interleukin-8 is regulated by miR-203 at the posttranscriptional level in primary human keratinocytes. Eur. J. Dermatol. 2013, 19, 1997. [Google Scholar]
- Xiao, Y.; Wang, H.; Wang, C.; Zeng, B.; Tang, X.; Zhang, Y.; Peng, Y.; Luo, M.; Huang, P.; Yang, Z. miR-203 promotes HaCaT cell overproliferation through targeting LXR-α and PPAR-γ. Cell Cycle 2020, 19, 1928–1940. [Google Scholar] [CrossRef] [PubMed]
- Luan, L.; Shi, J.; Yu, Z.; Andl, T. The major miR-31 target genes STK40 and LATS2 and their implications in the regulation of keratinocyte growth and hair differentiation. Exp. Dermatol. 2017, 26, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Borska, L.; Andrys, C.; Chmelarova, M.; Kovarikova, H.; Krejsek, J.; Hamakova, K.; Beranek, M.; Palicka, V.; Kremlacek, J.; Borsky, P.; et al. Roles of miR-31 and endothelin-1 in psoriasis vulgaris: Pathophysiological functions and potential biomarkers. Physiol. Res. 2017, 66, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Xu, Z.; Lou, F.; Zhang, L.; Ke, F.; Bai, J.; Liu, Z.; Liu, J.; Wang, H.; Zhu, H.; et al. NF-κB-induced microRNA-31 promotes epidermal hyperplasia by repressing protein phosphatase 6 in psoriasis. Nat. Commun. 2015, 6, 7652. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Meisgen, F.; Butler, L.M.; Han, G.; Wang, X.J.; Söderberg-Nauclér, C.; Ståhle, M.; Pivarcsi, A.; Sonkoly, E. MicroRNA-31 is overexpressed in psoriasis and modulates inflammatory cytokine and chemokine production in keratinocytes via targeting serine/threonine kinase 40. J. Immunol. 2013, 190, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chang, W.; Yang, X.; Cheng, Y.; Zhao, X.; Zhou, L.; Li, J.; Li, J.; Zhang, K. Levels of miR-31 and its target genes in dermal mesenchymal cells of patients with psoriasis. Int. J. Dermatol. 2019, 58, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Fang, X.; Zhang, Z.H.; Huang, Q.; Yan, K.X.; Kang, K.F.; Han, L.; Zheng, Z.Z. Dysregulation of miRNA146a versus IRAK1 induces IL-17 persistence in the psoriatic skin lesions. Immunol. Lett. 2012, 148, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Crone, S.G.; Jacobsen, A.; Federspiel, B.; Bardram, L.; Krogh, A.; Lund, A.H.; Friis-Hansen, L. microRNA-146a inhibits G protein-coupled receptor-mediated activation of NF-kappaB by targeting CARD10 and COPS8 in gastric cancer. Mol. Cancer 2012, 11, 71. [Google Scholar] [CrossRef]
- Hung, P.S.; Liu, C.J.; Chou, C.S.; Kao, S.Y.; Yang, C.C.; Chang, K.W.; Chiu, T.H.; Lin, S.C. miR-146a enhances the oncogenicity of oral carcinoma by concomitant targeting of the IRAK1, TRAF6 and NUMB genes. PLoS ONE 2013, 8, e79926. [Google Scholar] [CrossRef]
- Rebane, A.; Runnel, T.; Aab, A.; Maslovskaja, J.; Rückert, B.; Zimmermann, M.; Plaas, M.; Kärner, J.; Treis, A.; Pihlap, M.; et al. MicroRNA-146a alleviates chronic skin inflammation in atopic dermatitis through suppression of innate immune responses in keratinocytes. J. Allergy Clin. Immunol. 2014, 134, 836–847. [Google Scholar] [CrossRef]
- García-Rodríguez, S.; Arias-Santiago, S.; Blasco-Morente, G.; Orgaz-Molina, J.; Rosal-Vela, A.; Navarro, P.; Magro-Checa, C.; Martínez-López, A.; Ruiz, J.C.; Raya, E.; et al. Increased expression of microRNA-155 in peripheral blood mononuclear cells from psoriasis patients is related to disease activity. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 312–322. [Google Scholar] [CrossRef] [PubMed]
- El-Komy, M.; Amin, I.; El-Hawary, M.S.; Saadi, D.; Shaker, O. Upregulation of the miRNA-155, miRNA-210, and miRNA-20b in psoriasis patients and their relation to IL-17. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420933742. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Leng, H.; Shi, X.; Ji, J.; Fu, J.; Leng, H. MiR-155 promotes cell proliferation and inhibits apoptosis by PTEN signaling pathway in the psoriasis. Biomed. Pharmacother. 2017, 90, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Soonthornchai, W.; Tangtanatakul, P.; Meephansan, J.; Ruchusatsawat, K.; Reantragoon, R.; Hirankarn, N.; Wongpiyabovorn, J. Down-regulation of miR-155 after treatment with narrow-band UVB and methotrexate associates with apoptosis of keratinocytes in psoriasis. Asian Pac. J. Allergy Immunol. 2019. [Google Scholar] [CrossRef]
- Luo, Q.; Zeng, J.; Li, W.; Lin, L.; Zhou, X.; Tian, X.; Liu, W.; Zhang, L.; Zhang, X. Silencing of miR-155 suppresses inflammatory responses in psoriasis through inflammasome NLRP3 regulation. Int. J. Mol. Med. 2018, 42, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Luomei, J.; Huang, P.; Zhou, R.; Peng, Y. The miR-155/GATA3/IL37 axis modulates the production of proinflammatory cytokines upon TNF-α stimulation to affect psoriasis development. Exp. Dermatol. 2020. [Google Scholar] [CrossRef]
- Hou, R.X.; Liu, R.F.; Zhao, X.C.; Jia, Y.R.; An, P.; Hao, Z.P.; Li, J.Q.; Li, X.H.; Yin, G.H.; Zhang, K.M. Increased miR-155-5p expression in dermal mesenchymal stem cells of psoriatic patients: Comparing the microRNA expression profile by microarray. Genet Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Meisgen, F.; Xu, N.; Wei, T.; Janson, P.C.; Obad, S.; Broom, O.; Nagy, N.; Kauppinen, S.; Kemény, L.; Ståhle, M.; et al. MiR-21 is up-regulated in psoriasis and suppresses T cell apoptosis. Exp. Dermatol. 2012, 21, 312–314. [Google Scholar] [CrossRef]
- Jia, H.Y.; Zhang, K.; Lu, W.J.; Xu, G.W.; Zhang, J.F.; Tang, Z.L. LncRNA MEG3 influences the proliferation and apoptosis of psoriasis epidermal cells by targeting miR-21/caspase-8. BMC Mol. Cell. Biol. 2019, 20, 46. [Google Scholar] [CrossRef]
- Yang, C.; Luo, L.; Bai, X.; Shen, K.; Liu, K.; Wang, J.; Hu, D. Highly-expressed micoRNA-21 in adipose derived stem cell exosomes can enhance the migration and proliferation of the HaCaT cells by increasing the MMP-9 expression through the PI3K/AKT pathway. Arch. Biochem. Biophys. 2020, 681, 108259. [Google Scholar] [CrossRef]
- Degueurce, G.; D’Errico, I.; Pich, C.; Ibberson, M.; Schütz, F.; Montagner, A.; Sgandurra, M.; Mury, L.; Jafari, P.; Boda, A.; et al. Identification of a novel PPARβ/δ/miR-21-3p axis in UV-induced skin inflammation. EMBO Mol. Med. 2016, 8, 919–936. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Brodin, P.; Wei, T.; Meisgen, F.; Eidsmo, L.; Nagy, N.; Kemeny, L.; Stahle, M.; Sonkoly, E.; Pivarcsi, A. MiR-125b, a microRNA downregulated in psoriasis, modulates keratinocyte proliferation by targeting FGFR2. J. Investig. Dermatol. 2011, 131, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, A.; Jinnin, M.; Yamane, K.; Fujisawa, A.; Sakai, K.; Masuguchi, S.; Fukushima, S.; Maruo, K.; Ihn, H. microRNA-mediated keratinocyte hyperproliferatio in psoriasis vulgaris. Br. J. Dermatol. 2011, 165, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Sun, Z.; Dang, E.; Li, B.; Fang, H.; Li, J.; Gao, L.; Zhang, K.; Wang, G. TGFβ/SMAD/microRNA-486-3p Signaling Axis Mediates Keratin 17 Expression and Keratinocyte Hyperproliferation in Psoriasis. J. Investig. Dermatol. 2017, 137, 2177–2186. [Google Scholar] [CrossRef]
- Feng, S.J.; Chu, R.Q.; Ma, J.; Wang, Z.X.; Zhang, G.J.; Yang, X.F.; Song, Z.; Ma, Y.Y. MicroRNA138 regulates keratin 17 protein expression to affect HaCaT cell proliferation and apoptosis by targeting hTERT in psoriasis vulgaris. Biomed. Pharmacother. 2017, 85, 169–176. [Google Scholar] [CrossRef]
- Wang, C.; Zong, J.; Li, Y.; Wang, X.; Du, W.; Li, L. MiR-744-3p regulates keratinocyte proliferation and differentiation via targeting KLLN in psoriasis. Exp. Dermatol. 2019, 28, 283–291. [Google Scholar] [CrossRef]
- Li, Y.; Su, J.; Li, F.; Chen, X.; Zhang, G. MiR-150 regulates human keratinocyte proliferation in hypoxic conditions through targeting HIF-1α and VEGFA: Implications for psoriasis treatment. PLoS ONE 2017, 12, e0175459. [Google Scholar] [CrossRef]
- A, R.; Yu, P.; Hao, S.; Li, Y. MiR-876-5p suppresses cell proliferation by targeting Angiopoietin-1 in the psoriasis. Biomed. Pharmacother. 2018, 103, 1163–1169. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, X.; Wang, Y. miR-183-3p suppresses proliferation and migration of keratinocyte in psoriasis by inhibiting GAB1. Hereditas 2020, 157, 28. [Google Scholar] [CrossRef]
- Zhao, X.; Li, R.; Qiao, M.; Yan, J.; Sun, Q. MiR-548a-3p Promotes Keratinocyte Proliferation Targeting PPP3R1 after Being Induced by IL-22. Inflammation 2018, 41, 496–504. [Google Scholar] [CrossRef]
- Zhu, H.; Hou, L.; Liu, J.; Li, Z. MiR-217 is down-regulated in psoriasis and promotes keratinocyte differentiation via targeting GRHL2. Biochem. Biophys. Res. Commun. 2016, 471, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Chowdhari, S.; Sardana, K.; Saini, N. miR-4516, a microRNA downregulated in psoriasis inhibits keratinocyte motility by targeting fibronectin/integrin α9 signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 3142–3152. [Google Scholar] [CrossRef]
- Yu, X.; An, J.; Hua, Y.; Li, Z.; Yan, N.; Fan, W.; Su, C. MicroRNA-194 regulates keratinocyte proliferation and differentiation by targeting Grainyhead-like 2 in psoriasis. Pathol. Res. Pract. 2017, 213, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; He, S.; Zhu, Y.; Feng, B.; Su, Z.; Liu, B.; Xu, F.; Wang, X.; Liu, H.; Chutian, L.; et al. Downregulated miR-187 contributes to the keratinocytes hyperproliferation in psoriasis. J. Cell. Physiol. 2019, 234, 3661–3674. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Tian, Y.; Yao, X.; Liu, W.; Zhang, Y.; Yang, Z. MiR-99a inhibits keratinocyte proliferation by targeting Frizzled-5 (FZD5) / FZD8 through β-catenin signaling in psoriasis. Pharmazie 2017, 72, 461–467. [Google Scholar]
- Xiong, Y.; Chen, H.; Liu, L.; Lu, L.; Wang, Z.; Tian, F.; Zhao, Y. microRNA-130a Promotes Human Keratinocyte Viability and Migration and Inhibits Apoptosis Through Direct Regulation of STK40-Mediated NF-κB Pathway and Indirect Regulation of SOX9-Meditated JNK/MAPK Pathway: A Potential Role in Psoriasis. DNA Cell Biol. 2017, 36, 219–226. [Google Scholar] [CrossRef]
- Jiang, M.; Ma, W.; Gao, Y.; Jia, K.; Zhang, Y.; Liu, H.; Sun, Q. IL-22-induced miR-122-5p promotes keratinocyte proliferation by targeting Sprouty2. Exp. Dermatol. 2017, 26, 368–374. [Google Scholar] [CrossRef]
- Feng, S.; Wang, L.; Liu, W.; Zhong, Y.; Xu, S. MiR-126 correlates with increased disease severity and promotes keratinocytes proliferation and inflammation while suppresses cells’ apoptosis in psoriasis. J. Clin. Lab Anal. 2018, 32, e22588. [Google Scholar] [CrossRef]
- Yan, J.J.; Qiao, M.; Li, R.H.; Zhao, X.T.; Wang, X.Y.; Sun, Q. Downregulation of miR-145-5p contributes to hyperproliferation of keratinocytes and skin inflammation in psoriasis. Br. J. Dermatol. 2019, 180, 365–372. [Google Scholar] [CrossRef]
- Zhang, W.; Yi, X.; An, Y.; Guo, S.; Li, S.; Song, P.; Chang, Y.; Zhang, S.; Gao, T.; Wang, G.; et al. MicroRNA-17-92 cluster promotes the proliferation and the chemokine production of keratinocytes: Implication for the pathogenesis of psoriasis. Cell Death Dis. 2018, 9, 567. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, X.; Wang, L.; Ma, W.; Sun, Q. miR-320b Is Down-Regulated in Psoriasis and Modulates Keratinocyte Proliferation by Targeting AKT3. Inflammation 2018, 41, 2160–2170. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, C.; Zeng, B.; Tang, X.; Zhang, Y.; Xiang, L.; Mi, L.; Pan, Y.; Wang, H.; Yang, Z. miR124-3p/FGFR2 axis inhibits human keratinocyte proliferation and migration and improve the inflammatory microenvironment in psoriasis. Mol. Immunol. 2020, 122, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.C.; Warren, R.B.; Griffiths, C.E.; Ross, K. Expression of microRNA-184 in keratinocytes represses argonaute 2. J. Cell. Physiol. 2013, 228, 2314–2323. [Google Scholar] [CrossRef] [PubMed]
- Zibert, J.R.; Løvendorf, M.B.; Litman, T.; Olsen, J.; Kaczkowski, B.; Skov, L. MicroRNAs and potential target interactions in psoriasis. J. Dermatol. Sci. 2010, 58, 177–185. [Google Scholar] [CrossRef]
- Feng, C.; Bai, M.; Yu, N.Z.; Wang, X.J.; Liu, Z. MicroRNA-181b negatively regulates the proliferation of human epidermal keratinocytes in psoriasis through targeting TLR4. J. Cell. Mol. Med. 2017, 21, 278–285. [Google Scholar] [CrossRef]
- Srivastava, A.; Pasquali, L.; Pivarcsi, A.; Sonkoly, E. MiR-1307 is upregulated in psoriasis keratinocytes and promotes keratinocyte inflammatory response. In Proceedings of the 49th Annual ESDR Meeting, Bordeaux, France, 18 September 2019. [Google Scholar]
- Jiang, M.; Fang, H.; Dang, E.; Zhang, J.; Qiao, P.; Yu, C.; Yang, A.; Wang, G. Small extracellular vesicles containing miR-381-3p from keratinocytes promotes Th1/Th17 polarization in psoriasis. J. Investig. Dermatol. 2020, S0022-202X, 31938–31942. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, L.T.; Liang, G.P.; Zhan, P.; Deng, X.J.; Tang, Q.; Zhai, H.; Chang, C.C.; Su, Y.W.; Lu, Q.J. Up-regulation of microRNA-210 induces immune dysfunction via targeting FOXP3 in CD4(+) T cells of psoriasis vulgaris. Clin. Immunol. 2014, 150, 22–30. [Google Scholar] [CrossRef]
- Fu, D.; Yu, W.; Li, M.; Wang, H.; Liu, D.; Song, X.; Li, Z.; Tian, Z. MicroRNA-138 regulates the balance of Th1/Th2 via targeting RUNX3 in psoriasis. Immunol. Lett. 2015, 166, 55–62. [Google Scholar] [CrossRef]
- Lou, C.; Xiao, M.; Cheng, S.; Lu, X.; Jia, S.; Ren, Y.; Li, Z. MiR-485-3p and miR-485-5p suppress breast cancer cell metastasis by inhibiting PGC-1α expression. Cell Death Dis. 2016, 7, e2159. [Google Scholar] [CrossRef]
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic dermatitis. Nat. Rev. Dis. Primers 2018, 4, 1. [Google Scholar] [CrossRef]
- Nutten, S. Atopic dermatitis: Global epidemiology and risk factors. Ann. Nutr. Metab. 2015, 66, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.T.; Goodarzi, H.; Chen, H.Y. IgE, mast cells, and eosinophils in atopic dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Vaher, H.; Runnel, T.; Urgard, E.; Aab, A.; Carreras Badosa, G.; Maslovskaja, J.; Abram, K.; Raam, L.; Kaldvee, B.; Annilo, T.; et al. miR-10a-5p is increased in atopic dermatitis and has capacity to inhibit keratinocyte proliferation. Allergy 2019, 74, 2146–2156. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Li, Y.; Wu, J.; Xu, J. IFN-γ-induced microRNA-29b up-regulation contributes to keratinocyte apoptosis in atopic dermatitis through inhibiting Bcl2L2. Int. J. Clin. Exp. Pathol. 2017, 10, 10117–10126. [Google Scholar]
- Jia, H.Z.; Liu, S.L.; Zou, Y.F.; Chen, X.F.; Yu, L.; Wan, J.; Zhang, H.Y.; Chen, Q.; Xiong, Y.; Yu, B.; et al. MicroRNA-223 is involved in the pathogenesis of atopic dermatitis by affecting histamine-N-methyltransferase. Cell. Mol. Biol. 2018, 64, 103–107. [Google Scholar] [CrossRef]
- Chen, X.F.; Zhang, L.J.; Zhang, J.; Dou, X.; Shao, Y.; Jia, X.J.; Zhang, W.; Yu, B. MiR-151a is involved in the pathogenesis of atopic dermatitis by regulating interleukin-12 receptor β2. Exp. Dermatol. 2018, 27, 427–432. [Google Scholar] [CrossRef]
- Jia, Q.N.; Zeng, Y.P. Rapamycin blocks the IL-13-induced deficiency of Epidermal Barrier Related Proteins via upregulation of miR-143 in HaCaT Keratinocytes. Int. J. Med. Sci. 2020, 17, 2087–2094. [Google Scholar] [CrossRef]
- Kuhn, A.; Landmann, A. The classification and diagnosis of cutaneous lupus erythematosus. J. Autoimmun. 2014, 48, 14–19. [Google Scholar] [CrossRef]
- Durosaro, O.; Davis, M.; Reed, K.B.; Rohlinger, A.L. Incidence of cutaneous lupus erythematosus, 1965-2005: A population-based study. Arch. Dermatol. 2009, 145, 249–253. [Google Scholar] [CrossRef]
- Achtman, J.C.; Werth, V.P. Pathophysiology of cutaneous lupus erythematosus. Arthritis Res. Ther. 2015, 17, 182. [Google Scholar] [CrossRef]
- Dinulovic, I.; Furrer, R.; Di Fulvio, S.; Ferry, A.; Beer, M.; Handschin, C. PGC-1α modulates necrosis, inflammatory response, and fibrotic tissue formation in injured skeletal muscle. Skelet. Muscle 2016, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xue, H.B.; Wang, F.; Shu, C.M.; Zhang, J.H. MicroRNA-155 may be involved in the pathogenesis of atopic dermatitis by modulating the differentiation and function of T helper type 17 (Th17) cells. Clin. Exp. Immunol. 2015, 181, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Vennegaard, M.T.; Bonefeld, C.M.; Hagedorn, P.H.; Bangsgaard, N.; Løvendorf, M.B.; Ødum, N.; Woetmann, A.; Geisler, C.; Skov, L. Allergic contact dermatitis induces upregulation of identical microRNAs in humans and mice. Contact Dermat. 2012, 67, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, Y.; Yuan, W.; Yao, L.; Wang, S.; Jia, Z.; Wu, P.; Li, L.; Wei, P.; Wang, X.; et al. MicroRNA-155-5p is a key regulator of allergic inflammation, modulating the epithelial barrier by targeting PKIα. Cell Death Dis. 2019, 10, 884. [Google Scholar] [CrossRef] [PubMed]
- Schwingen, J.; Kaplan, M.; Kurschus, F.C. Current concepts in inflammatory skin diseases evolved by transcriptome analysis: In-depth analysis of atopic dermatitits and psoriasis. Int. J. Mol. Sci. 2020, 21, 699. [Google Scholar] [CrossRef]
- Choy, D.F.; Hsu, D.K.; Seshasayee, D.; Fung, M.A.; Modrusan, Z.; Martin, F.; Liu, F.; Arron, J.R. Comparative transcriptomic analyses of atopic dermatitis and psoriasis reveal shared neutrophilic inflammation. J. Allergy Clin. Immunol. 2012, 130, 1335–1343. [Google Scholar] [CrossRef]
- Jabbari, A.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Gonzalez, J.; Cueto, I.; Franks, A.G.; Krueger, J.G. Dominant Th1 and minimal Th17 skewing in discoid lupus revealed by transcriptomic comparison with psoriasis. J. Investig. Dermatol. 2014, 134, 87–95. [Google Scholar] [CrossRef]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stähler, C.; Meese, E.; et al. Distribution of miRNA expression across human tissues. Nucleic Acids Res. 2016, 44, 3865–3877. [Google Scholar] [CrossRef]
- Ciesla, M.; Skrzypek, K.; Kozakowska, M.; Loboda, A.; Jozkowicz, A.; Dulak, J. MicroRNAs as biomarkers of disease onset. Anal. Bioanal. Chem. 2011, 401, 2051–2061. [Google Scholar] [CrossRef]
- Løvendorf, M.B.; Zibert, J.R.; Gyldenløve, M.; Røpke, M.A.; Skov, L. MicroRNA-223 and miR-143 are important systemic biomarkers for disease activity in psoriasis. J. Dermatol. Sci. 2014, 75, 133–139. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, W.; Wei, C.; Wang, L.; Zhu, G.; Shi, Q.; Li, S.; Ge, R.; Li, K.; Gao, L.; et al. Serum and skin levels of miR-369-3p in patients with psoriasis and their correlation with disease severity. Eur. J. Dermatol. 2013, 23, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Tsuru, Y.; Jinnin, M.; Ichihara, A.; Fujisawa, A.; Moriya, C.; Sakai, K.; Fukushima, S.; Ihn, H. miR-424 levels in hair shaft are increased in psoriatic patients. J. Dermatol. 2014, 41, 382–385. [Google Scholar] [CrossRef] [PubMed]
- Hirao, H.; Jinnin, M.; Ichihara, A.; Fujisawa, A.; Makino, K.; Kajihara, I.; Sakai, K.; Fukushima, S.; Inoue, Y.; Ihn, H. Detection of hair root miR-19a as a novel diagnostic marker for psoriasis. Eur. J. Dermatol. 2013, 23, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Chatzikyriakidou, A.; Voulgari, P.V.; Georgiou, I.; Drosos, A.A. The role of microRNA-146a (miR-146a) and its target IL-1R-associated kinase (IRAK1) in psoriatic arthritis susceptibility. Scand J. Immunol. 2010, 71, 382–385. [Google Scholar] [CrossRef]
- Wang, G.; Tam, L.S.; Li, E.K.; Kwan, B.C.H.; Chow, K.M.; Luk, C.C.W.; Li, P.K.T.; Szeto, C.C. Serum and urinary free microRNA level in patients with systemic lupus erythematosus. Lupus 2011, 20, 493–500. [Google Scholar] [CrossRef]
- Méndez-Flores, S.; Furuzawa-Carballeda, J.; Hernández-Molina, G.; Ramírez-Martinez, G.; Regino-Zamarripa, N.E.; Ortiz-Quintero, B.; Jiménez-Alvarez, L.; Cruz-Lagunas, A.; Zúñiga, J. MicroRNA Expression in Cutaneous Lupus: A New Window to Understand Its Pathogenesis. Mediat. Inflamm. 2019, 2019, 5049245. [Google Scholar] [CrossRef]
- Hashad, D.; Abdelmagid, M.; Elsherif, S. microRNA146a expression in lupus patients with and without renal complications. J. Clin. Lab Anal. 2012, 26, 35–40. [Google Scholar] [CrossRef]
- Lv, Y.; Qi, R.; Xu, J.; Di, Z.; Zheng, H.; Huo, W.; Zhang, L.; Chen, H.; Gao, X. Profiling of serum and urinary microRNAs in children with atopic dermatitis. PLoS ONE 2014, 9, e115448. [Google Scholar] [CrossRef]
- Meng, L.; Li, M.; Gao, Z.; Ren, H.; Chen, J.; Liu, X.; Cai, Q.; Jiang, L.; Ren, X.; Yu, Q.; et al. Possible role of hsa-miR-194-5p, via regulation of HS3ST2, in the pathogenesis of atopic dermatitis in children. Eur. J. Dermatol. 2019, 29, 603–613. [Google Scholar] [CrossRef]
- Spuls, P.I.; Lecluse, L.; Poulsen, M.; Bos, J.; Stern, R.S.; Nijsten, T. How good are clinical severity and outcome measures for psoriasis?: Quantitative evaluation in a systematic review. J. Investig. Dermatol. 2010, 130, 933–943. [Google Scholar] [CrossRef]
- Yang, Z.; Zeng, B.; Tang, X.; Wang, H.; Wang, C.; Yan, Z.; Huang, P.; Pan, Y.; Xu, B. MicroRNA-146a and miR-99a are potential biomarkers for disease activity and clinical efficacy assessment in psoriasis patients treated with traditional Chinese medicine. J. Ethnopharmacol. 2016, 194, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zou, J.; Mao, J.; Guo, D.; Wu, M.; Xu, N.; Zhou, J.; Zhang, Y.; Guo, W.; Jin, W. Plasma miR-126 expression correlates with risk and severity of psoriasis and its high level at baseline predicts worse response to Tripterygium wilfordii Hook F in combination with acitretin. Biomed. Pharmacother. 2019, 115, 108761. [Google Scholar] [CrossRef] [PubMed]
- Magenta, A.; D’Agostino, M.; Sileno, S.; Di Vito, L.; Uras, C.; Abeni, D.; Martino, F.; Barillà, F.; Madonna, S.; Albanesi, C.; et al. The Oxidative Stress-Induced miR-200c Is Upregulated in Psoriasis and Correlates with Disease Severity and Determinants of Cardiovascular Risk. Oxid. Med. Cell. Longev. 2019, 2019, 8061901. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Su, Y.; Wu, H.; Wu, R.; Zhang, P.; Dai, Y.; Chan, T.-M.; Zhao, M.; Lu, Q. The role of microRNAs in autoinmune diseases with skin involvement. Scand. J. Immunol. 2015, 81, 153–165. [Google Scholar] [CrossRef]
- Chicharro, P.; Rodríguez-Jiménez, P.; Llamas-Velasco, M.; Montes, N.; Sanz-García, A.; Cibrian, D.; Vara, A.; Gómez, M.J.; Jiménez-Fernández, M.; Martínez-Fleta, P.; et al. Expression of miR-135b in Psoriatic Skin and Its Association with Disease Improvement. Cells 2020, 9, 1603. [Google Scholar] [CrossRef]
- Albrecht, J.; Taylor, L.; Berlin, J.A.; Dulay, S.; Ang, S.; Fakharzadeh, S.; Kantor, J.; Kim, E.; Militello, G.; McGinnis, K.; et al. The CLASI (Cutaneous Lupus Erythematosus Disease Area and Severity Index): An outcome instrument for cutaneous lupus erythematosus. J. Investig. Dermatol. 2005, 125, 889–894. [Google Scholar] [CrossRef]
- Zhou, H.; Hasni, S.A.; Perez, P.; Tandon, M.; Jang, S.I.; Zheng, C.; Kopp, J.B.; Austin, H., 3rd; Balow, J.E.; Alevizos, I.; et al. miR-150 promotes renal fibrosis in lupus nephritis by downregulating SOCS1. J. Am. Soc. Nephrol. 2013, 24, 1073–1087. [Google Scholar] [CrossRef]
- Honda, N.; Jinnin, M.; Kira-Etoh, T.; Makino, K.; Kajihara, I.; Makino, T.; Fukushima, S.; Inoue, Y.; Okamoto, Y.; Hasegawa, M.; et al. miR-150 down-regulation contributes to the constitutive type I collagen overexpression in scleroderma dermal fibroblasts via the induction of integrin β3. Am. J. Pathol. 2013, 182, 206–216. [Google Scholar] [CrossRef]
- Pivarcsi, A.; Meisgen, F.; Xu, N.; Ståhle, M.; Sonkoly, E. Changes in the level of serum microRNAs in patients with psoriasis after antitumour necrosis factor-α therapy. Br. J. Dermatol. 2013, 169, 563–570. [Google Scholar] [CrossRef]
- Raaby, L.; Langkilde, A.; Kjellerup, R.B.; Vinter, H.; Khatib, S.H.; Hjuler, K.F.; Johansen, C.; Iversen, L. Changes in mRNA expression precede changes in microRNA expression in lesional psoriatic skin during treatment with adalimumab. Br. J. Dermatol. 2015, 173, 436–447. [Google Scholar] [CrossRef]
- Mensà, E.; Recchioni, R.; Marcheselli, F.; Giuliodorri, K.; Consales, V.; Molinelli, E.; Prattichizzo, F.; Rippo, M.R.; Campanati, A.; Procopio, A.D.; et al. MiR-146a-5p correlates with clinical efficacy in patients with psoriasis treated with the tumour necrosis factor-alpha inhibitor adalimumab. Br. J. Dermatol. 2018, 179, 787–789. [Google Scholar] [CrossRef]
- Pei, D.; Cao, J.; Qin, G.; Wang, X. Measurement of circulating miRNA-125a exhibits good value in the management of etanercept-treated psoriatic patients. J. Dermatol. 2020, 47, 140–146. [Google Scholar] [CrossRef]
- García-Rodríguez, S.; Arias-Santiago, S.; Orgaz-Molina, J.; Magro-Checa, C.; Valenzuela, I.; Navarro, P.; Naranjo-Sintes, R.; Sancho, J.; Zubiaur, M. Abnormal levels of expression of plasma microRNA-33 in patients with psoriasis. Actas Dermosifiliogr. 2014, 105, 497–503. [Google Scholar] [CrossRef]
- Seifeldin, N.S.; El Sayed, S.B.; Asaad, M.K. Increased MicroRNA-1266 levels as a biomarker for disease activity in psoriasis vulgaris. Int. J. Dermatol. 2016, 55, 1242–1247. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Han, H.D.; Lopez-Berestein, G.; Sood, A.K. MicroRNA therapeutics: Principles, expectations, and challenges. Chin. J. Cancer 2011, 30, 368–370. [Google Scholar] [CrossRef]
- Lawrence, P.; Ceccoli, J. Advances in the Application and Impact of MicroRNAs as Therapies for Skin Disease. BioDrugs 2017, 31, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Guinea-Viniegra, J.; Jiménez, M.; Schonthaler, H.B.; Navarro, R.; Delgado, Y.; Concha-Garzón, M.J.; Tshachler, E.; Obad, S.; Daudén, E.; Wagner, E.F. Targeting miR-21 to treat psoriasis. Sci. Transl. Med. 2014, 6, 225re1. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Liu, R.; Fan, T.; Liao, L.; Wang, S.; Geng, W.; Wang, T.; Shi, W.; Ruan, Q. miR-340 Alleviates Psoriasis in Mice through Direct Targeting of IL-17A. J. Immunol. 2018, 201, 1412–1420. [Google Scholar] [CrossRef]
- Srivastava, A.; Nikamo, P.; Lohcharoenkal, W.; Dongquing, L.; Meisgen, F.; Landén, N.X.; Stahle, M.; Pivarcsi, A.; Sonkoly, E. MicroRNA-146a suppresses IL-17-mediated skin inflammation and is genetically associated with psoriasis. J. Allergy Clin. Immunol. 2017, 139, 550–561. [Google Scholar] [CrossRef]
- Feng, H.; Wu, R.; Zhang, S.; Kong, Y.; Liu, Z.; Wu, H.; Wang, H.; Su, Y.; Zhao, M.; Lu, Q. Topical administration of nanocarrier miRNA-210 antisense ameliorates imiquimod-induced psoriasis-like dermatitis in mice. J. Dermatol. 2020, 47, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Han, S.B.; Ham, H.J.; Park, J.H.; Lee, J.S.; Hwang, D.Y.; Jung, Y.S.; Yoon, D.Y.; Hong, J.T. IL-32γ suppressed atopic dermatitis through inhibition of miR-205 expression via inactivation of nuclear factor-kappa B. J. Allergy Clin. Immunol. 2020, 146, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, D.; Wang, A.; Chu, T.; Lohcharoenkal, W.; Zheng, X.; Grünler, J.; Narayanan, S.; Eliasson, S.; Herter, E.K.; et al. MicroRNA-132 with Therapeutic Potential in Chronic Wounds. J. Investig. Dermatol. 2017, 137, 2630–2638. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Shu, B.; Xu, Y.; Liu, J.; Wnag, P.; Chen, L.; Zhao, J.; Liu, X.; Qi, S.; Xiong, K.; et al. microRNA-203 Modulates Wound Healing and Scar Formation via Suppressing Hes1 Expression in Epidermal Stem Cells. Cell. Physiol. Biochem. 2018, 49, 2333–2347. [Google Scholar] [CrossRef]
- Maurer, B.; Stanczyk, J.; Jüngel, A.; Akhmetshina, A.; Trenkmann, M.; Brock, M.; Kowal-Bielecka, O.; Gay, R.E.; Michel, B.A.; Distle, J.H.W.; et al. MicroRNA-29, a key regulator of collagen expression in systemic sclerosis. Arthritis Rheum. 2010, 62, 1733–1743. [Google Scholar] [CrossRef]
- Gallant-Behm, C.L.; Piper, J.; Lynch, J.M.; Seto, A.G.; Hong, S.J.; Mustoe, T.A.; Maari, C.; Pestano, L.A.; Dalby, C.M.; Jackson, A.L.; et al. A MicroRNA-29 Mimic (Remlarsen) Represses Extracellular Matrix Expression and Fibroplasia in the Skin. J. Investig. Dermatol. 2019, 139, 1073. [Google Scholar] [CrossRef]
- Ross, K. Towards topical microRNA-directed threap for epidermal disorders. J. Control. Release 2018, 269, 13–147. [Google Scholar] [CrossRef]
- Nastiti, C.M.R.R.; Ponto, T.; Abd, E.; Grice, J.E.; Benson, H.A.; Roberts, M.S. Topical nano and microemulsions for skin delivery. Pharmaceutics 2017, 9, 37. [Google Scholar] [CrossRef]
- Kasting, G.B.; Barai, N.D.; Wang, T.F.; Nitsche, J.M. Mobility of water in human stratum corneum. J. Pharm. Sci. 2003, 92, 2326–2340. [Google Scholar] [CrossRef]
- Abdel-Mottaleb, M.M.; Try, C.; Pellequer, Y.; Lamprecht, A. Nanomedicine strategies for targeting skin inflammation. Nanomedicine 2014, 9, 1727–1743. [Google Scholar] [CrossRef]
- Liu, F.; Wang, C.; Gao, Y.; Li, X.; Tian, F.; Zhang, Y.; Fu, M.; Li, P.; Wang, Y.; Wang, F. Current transport systems and clinical applications for small interfering RNA (siRNA) drugs. Mol. Diagn. Ther. 2018, 22, 551–569. [Google Scholar] [CrossRef] [PubMed]
- Desmet, E.; Bracke, S.; Forier, K.; Taevernier, L.; Stuart, M.C.A.; Spiegeleer, B.D.; Raemdonck, K.; Gele, M.V.; Lambert, J. An elastic liposomal formulation for RNAi-based topical treatment of skin disorders: Proof-of-concept in the treatment of psoriasis. Int. J. Pharm. 2016, 500, 268–274. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Condition | Tissue/Cell/Fluid | Expression | Target Genes | Biological Role | Ref. |
|---|---|---|---|---|---|---|
| miR-203 | Psoriasis | Keratinocytes | Upregulated | SOCS3 NR1H3 PPARG | Keratinocyte proliferation, modulation of cytokines: TNF-α, IL-24 and IL-8 and angiogenesis. | [14,19,20,21,22] |
| miR-31 | Psoriasis DLE | Keratinocytes Blood DMSCs | Upregulated (Blood and Keratinocytes) Downregulated (DMSCs) | PPP6C STK40 | Keratinocyte proliferation and apoptosis. Promotes Inflammation via NFKB1 activation and chemokine and cytokine production (CXCL1, CXCL5, IL-8, IL-1B and IL-12). Neutrophile and intermediate monocyte recruitment. | [16,24,25,26] |
| miR-146a | Psoriasis AD | Keracinocytes Serum | Upregulated | CCL5 IRAK1 CARD10 | Protective role disminishing keratinocyte proliferation and inflammation supressing IL-17, CCL5, CCL8 and IFNγ. | [14,28,29,30,31] |
| miR-155 | Psoriasis AD | Keratinocytes Blood T cells | Upregulated | CTLA4 PKIA GATA3 CASP3 | Promoted epidermal proliferation, inflammation, TJ disruption and inhibits apoptosis. T cell proliferation and promotion of TH17 responses. | [32,33,34,35,36,37,38,71,72,73,74] |
| miR-21 | Psoriasis | Keratinocytes Blood T cells | Upregulated | CASP8 SMAD7 | T cell activation and inhibition of apoptosis. Keratinocyte proliferation and inflammation (IL-1β, CCL5 and CXCL10). | [39,40,41,42] |
| miR-125b | Psoriasis | Keratinocytes | Downregulated | FGFR2 | Keratinocyte proliferation and differentiation. | [43] |
| miR-424 | Psoriasis | Keratinocytes Serum | Downregulated | n.d. | Keratinocyte proliferation via MEK1/cyclin E1. | [44] |
| miR-486-3p | Psoriasis | Keratinocytes | Downegulated | K17 | Keratinocyte proliferation. | [45] |
| miR-138 | Psoriasis | Keratinocytes | Downregulated | K17 | Keratinocyte proliferation and apoptosis reduction. | [46] |
| miR-744-3p | Psoriasis | Keratinocytes | Upregulated | KLLN | Keratinocyte proliferation and differentiation. | [47] |
| miR-150 | Psoriasis | Keratinocytes | Downregulated | HIF1A VEGFA | Keratinocyte proliferation in hypoxic conditions. | [48] |
| miR-876-5p | Psoriasis | Skin Blood | Downregulated | ANG-1 | HaCAT proliferation via PI3K/AKT, cell adhesion and invasion. | [49] |
| miR-183-3p | Psoriasis | Keratinocytes | Downregulated | GAB1 | Proliferation and migration of HaCat cells. | [50] |
| miR-548a-3p | Psoriasis | Keratinocytes | Upregulated | PPP3R1 | Keratinocyte proliferation. | [51] |
| miR-217 | Psoriasis | Keratinocytes | Downregulated | GFHL2 | Keratinocyte differentiation. | [52] |
| miR-4516 | Psoriasis | Keratinocytes | Downregulation | FN1 ITGA9 | Accelerated migration, resistance to apoptosis and differentiation. | [53] |
| miR-194-5p | Psoriasis AD | Keratinocytes | Downregulated | GRHL2 HS3ST2 | Keratinocyte proliferation and inhibition of differentiation. | [54] |
| miR-187 | Psoriasis | Keratinocytes | Downregulated | CD276 | Keratinocute proliferation. | [55] |
| miR-99a | Psoriasis | Keratinocytes | Downregulated | FZD5/FDZ8 | Keratinocyte proliferation. | [56] |
| miR-130a | Psoriasis | Keratinocytes | Upregulated | STK40 | Apoptosis inhibition and cell viability and migration promotion. Direct regulation NFKB pathway via STK40 and inditect regulation of JNK/MAPK pathway via SOX9. | [57] |
| miR-122-5p | Psoriasis | Keratinocytes | Upregulated | SPRY2 | Keratinocyte proliferation. | [58] |
| miR-126 | Psoriasis | Keratinocytes | Upregulated | n.d. | Keraintocyte proliferation and inflammation increasing TNFa, IFNg, IL17A, IL-22 and decreasing IL-10. Apoptosis inhibition. | [59] |
| miR-145-5p | Psoriasis | Keratinocytes | Downregulated | MLK3 | Cell proliferation and chemokine secretion via NF-kB and STAT 3 activation. | [60] |
| miR-17-92 | Psoriasis | Keratinocytes | Upregulated | CDKN2B | Keratinocyte proliferation and immune chemotaxis via secretion CXCL9, CXCL10, supression of SOCS1 and STAT1 activation. | [61] |
| miR-320b | Psoriasis | Keratinocytes | Downregulation | AKT3 | Keratinocyte proliferation and modulation of STAT3 and SAPK/JNKsingaling pathways. | [62] |
| miR-124-3p | Psoriasis | Keratinocytes | Downregulated | FGFR2 | Keratinocyte prolfieration, migration and inflammation. | [63] |
| miR-184 | Psoriasis | Keratinocytes | Upregulated | AGO2 | Cytokine dependent depletion of AGO2. | [64] |
| miR-221/222 | Psoriasis | Keratinocytes | Upregulated | n.d. | Keratinocyte and immune cells proliferation. | [65] |
| miR-181-b | Psoriasis | Keratinocytes | Downregulated | TLR4 | Inflammation and keratinocyte proliferation. | [66] |
| miR-1307-3p | Psoriasis | Keratinocytes | Upregulated | n.d. | Induces inflammatory mediators IL-8, IL-6 and CCL20. | [67] |
| miR-381-3p | Psoriasis | Keratinocytes (EVs) | Upregulated | FOXO1 UBR5 | Crosslink with T cells inducing TH1/TH17 polarisation. | [68] |
| miR-210 | Psoriasis | CD4+ T cells | Upregulated | FOXP3 | Induces immune T cell dysfunction. | [69] |
| miR-138 | Psoriasis | CD4+ T cells | Downregulated | RUNX3 | Modulation of TH1/TH2 balance. | [70] |
| miR-485-3p | DLE | T cells Fibroblasts | Upregulated | PPARGC1A | T cell activation and promotion of fibrotic processes. | [16] |
| miR-10a-5p | AD | Keratinocytes | Upregulated | HAS3 | Inhibitis keratinocyte proliferation. | [75] |
| miR-29b | AD | keratinocytes | Upregulated | BCL2 | Keratinocyte apoptosis. | [76] |
| miR-223 | AD | Blood | Upregulated | n.d. | Upregulation of HNMT indirectly to degrade excessive histamine. | [77] |
| miR-151a | AD | Blood | Upregulated | IL12RB2 | Regulation of TH1 cytokines (IL-2, IFN γ). | [78] |
| miR-143 | AD | Keratinocytes | Downregulatd | IL13RA1 | Regulation of IL-13 activitu and TH2 inflammation. | [79] |
| miRNA | Condition | Tissue/Cell/Fluid | Expression | Potential Application | Ref. |
|---|---|---|---|---|---|
| miR-223 miR-143 | Psoriasis | PBMCs | Upregulated | Diagnosis, assess disease severity and monitor treatment (metotrexate) response | [92] |
| miR-424 | Psoriasis | Hair shaft | Upregulated | Diagnosis | [94] |
| miR-19a | Psoriasis | Hair root | Upregulated | Diagnosis and duration of disease | [95] |
| miR-369-3p | Psoriasis | Serum Skin | Upregulated | Diagnosis (skin and serum) and severity of disease (skin) | [93,106] |
| miR-1266 | Psoriasis | Serum | Upregulated | Diasease activity | [116] |
| miR-126 | Psoriasis | Plasma | Downregulated | Disease Severity Comorbidities (carotid thickness) | [104] |
| miR-200c | Psoriasis | Plasma | Upregulated | Diseaase activity and Comorbidities (cardiovascular disease) | [105] |
| miR-155 | Psoriasis | PBMCs | Upregulated | Disease activity | [33] |
| miR-146a | Psoriasis | PBMCs | Upregulated | Disease activity Monitor treatment response (adalimumab) | [96,103] |
| miR-99a | Psoriasis | PBMCs | Downregulated | Disease activity | [103] |
| miR-135b | Psoriasis | Skin | Upregulated | Disease improvement | [107] |
| miR-125a | Psoriasis | Plasma | Downregulated | Diagnosis, and Monitor treatment response (etanercept) | [114] |
| miR-33 | Psoriasis | Plasma | Upregulated | Detection of comorbidities (elevated insulin levels) | [115] |
| miR-150 miR-1246 miR-21 | CLE (SCLE and DLE) | Serum | Downregulation | Diagnosis | [98] |
| miR-23b miR1246 miR-146 | SCLE | Serum | Downregulated | Diagnosis | [99] |
| miR-150 | SCLE | Serum | Downregulated | Disease Seveity | [109,110] |
| miR-203 | AD in children | Serum Urine | Upregulated in serum Downregulated in urine | Diagnosis and Disease severity | [100] |
| miR-483-5p | AD in children | Serum | Upregulated | Diagnosis and Detection of comorbidities (asthma/rhinitis) | [100] |
| miR-194-5p | AD in children | Plasma | Downregulated | Diagnosis | [101] |
| miR-155 | AD | CD4+ T cells | Upregulated | Diagnosis | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domingo, S.; Solé, C.; Moliné, T.; Ferrer, B.; Cortés-Hernández, J. MicroRNAs in Several Cutaneous Autoimmune Diseases: Psoriasis, Cutaneous Lupus Erythematosus and Atopic Dermatitis. Cells 2020, 9, 2656. https://doi.org/10.3390/cells9122656
Domingo S, Solé C, Moliné T, Ferrer B, Cortés-Hernández J. MicroRNAs in Several Cutaneous Autoimmune Diseases: Psoriasis, Cutaneous Lupus Erythematosus and Atopic Dermatitis. Cells. 2020; 9(12):2656. https://doi.org/10.3390/cells9122656
Chicago/Turabian StyleDomingo, Sandra, Cristina Solé, Teresa Moliné, Berta Ferrer, and Josefina Cortés-Hernández. 2020. "MicroRNAs in Several Cutaneous Autoimmune Diseases: Psoriasis, Cutaneous Lupus Erythematosus and Atopic Dermatitis" Cells 9, no. 12: 2656. https://doi.org/10.3390/cells9122656
APA StyleDomingo, S., Solé, C., Moliné, T., Ferrer, B., & Cortés-Hernández, J. (2020). MicroRNAs in Several Cutaneous Autoimmune Diseases: Psoriasis, Cutaneous Lupus Erythematosus and Atopic Dermatitis. Cells, 9(12), 2656. https://doi.org/10.3390/cells9122656