The miRNA Profile in Non-Hodgkin’s Lymphoma Patients with Secondary Myelodysplasia

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Isolation of Total RNA
2.3. NanoString nCounter miRNA Assay for miRNA Profiling
2.4. Validation of NanoString Results by RT-qPCR Analysis
2.5. Reverse Transcription
2.6. Real-Time PCR
2.7. Functional Analysis of miRNAs Using DIANA-miRPath
3. Results
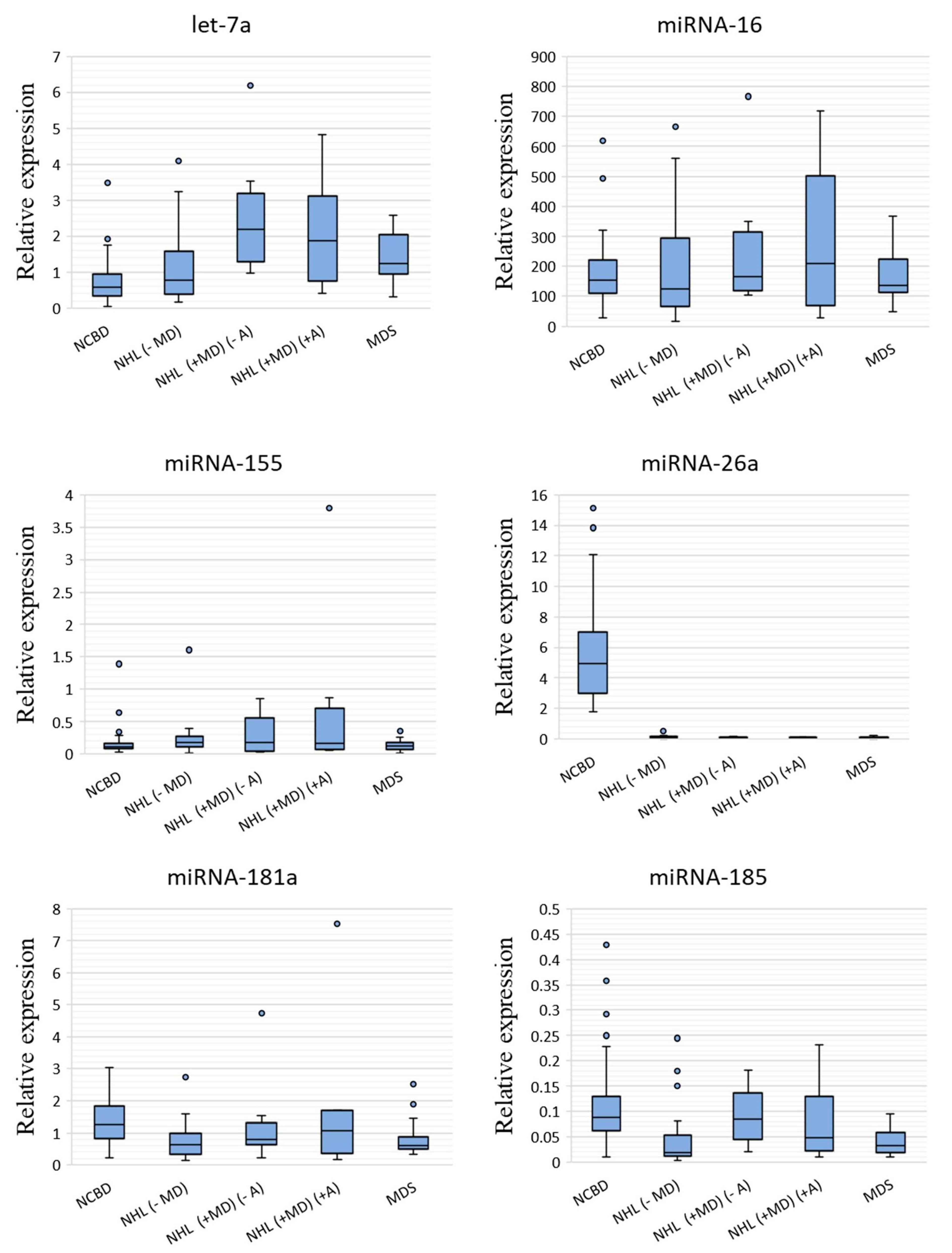
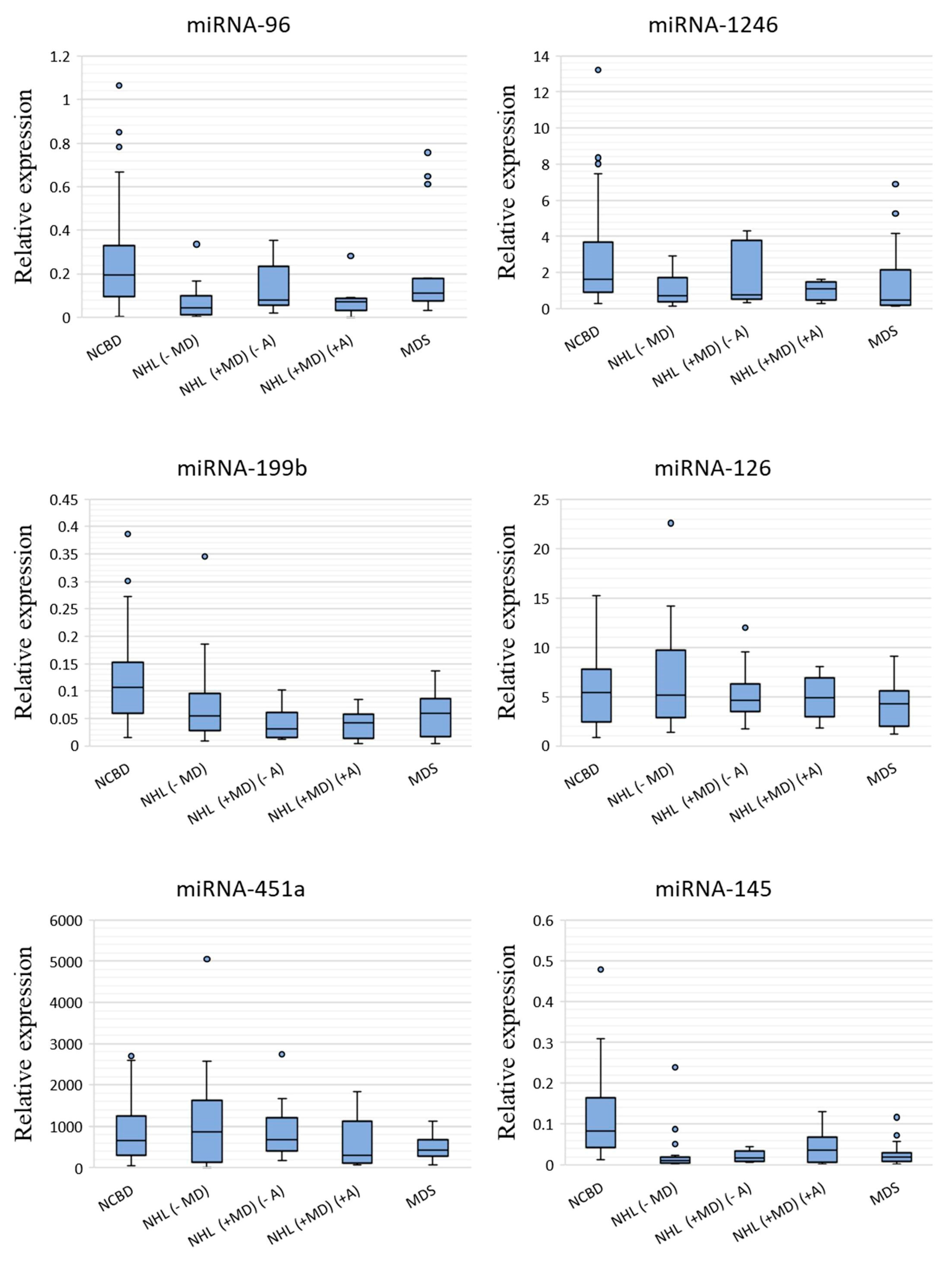
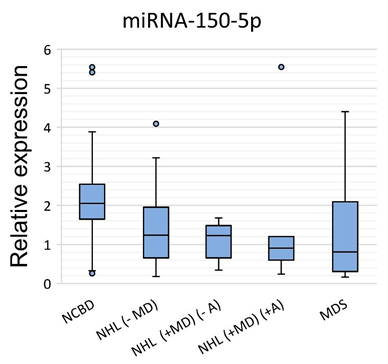
3.1. The Profile of miRNA Expression in BM Tumor Samples and NCBD
3.2. Bioinformatic Analysis of Genes Targeted by miRNAs and the Pathways Involved in Cancer Processes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mohammad, A.A. Myelodysplastic syndrome from theoretical review to clinical application view. Oncol. Rev. 2018, 12, 134–142. [Google Scholar] [CrossRef]
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.R.; et al. (Eds.) SEER Cancer Statistics Review, 1975–2017; Based on November 2019 SEER Data Submission, Posted to the SEER Web Site; National Cancer Institute: Bethesda, MD, USA, 2020. Available online: https://seer.cancer.gov/csr/1975_2017/ (accessed on 18 October 2020).
- Tanaka, T.N.; Bejar, R. MDS overlap disorders and diagnostic boundaries. Blood 2019, 133, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, R.; Deeg, H.J. Treatment-related myelodysplastic syndrome: Molecular characteristics and therapy. Curr. Opin. Hematol. 2011, 18, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Reis, G.P.; Bahadir, A.; Erduran, E. T-cell Non-Hodgkin lymphoma associated with myelodysplasia: A case report in a child. Med. Sci. Discov. 2018, 5, 202–206. [Google Scholar] [CrossRef]
- Yasmeen, T.; Ali, J.; Khan, K.; Siddiqui, N. Frequency and causes of anemia in Lymphoma patients. Pak. J. Med. Sci. 2019, 5, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Dozzo, M.; Zaja, F.; Volpetti, S.; Chiozzotto, M.; Puglisi, S.; Mazzucco, M.; Perali, G.; Fanin, R. Two cases of concomitant diffuse large B-cell lymphoma and myelodysplastic syndrome. Am. J. Hematol. 2014, 89, 1011–1013. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.H.; Zhu, J.Y.; Han, J.Y.; Shen, L.J.; Xu, L.; Chen, X.L.; Zheng, L.Y.; Chen, F.Y. Co-existent de novo myelodysplastic syndrome and T-cell non-Hodgkin lymphoma: A common origin or not? J. Int. Med. Res. 2009, 37, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Kotaki, R.; Koyama-Nasu, R.; Yamakawa, N.; Kotani, A. miRNAs in Normal and Malignant Hematopoiesis. Int. J. Mol. Sci. 2017, 18, 1495. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Hrustincova, A.; Krejcik, Z.; Kundrat, D.; Szikszai, K.; Belickova, M.; Pecherkova, P.; Klema, J.; Vesela, J.; Hruba, M.; Cermak, J.; et al. Circulating Small Noncoding RNAs Have Specific Expression Patterns in Plasma and Extracellular Vesicles in Myelodysplastic Syndromes and Are Predictive of Patient Outcome. Cells 2020, 9, 794. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Veryaskina, Y.A.; Titov, S.E.; Kovynev, I.B.; Pospelova, T.I.; Zhimulev, I.F. Prognostic Markers of Myelodysplastic Syndromes. Medicina 2020, 56, 376. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jiang, X.; Chen, J. The role of miR-150 in normal and malignant hematopoiesis. Oncogene 2014, 33, 3887–3893. [Google Scholar] [CrossRef] [PubMed]
- Sokol, L.; Caceres, G.; Volinia, S.; Alder, H.; Nuovo, G.J.; Liu, C.G.; McGraw, K.; Clark, J.A.; Sigua, C.A.; Chen, D.T.; et al. Identification of a risk dependent microRNA expression signature in myelodysplastic syndromes. Br. J. Haematol. 2011, 153, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Mazan-Mamczarz, K.; Gartenhaus, R.B. Role of microRNA deregulation in the pathogenesis of diffuse large B-cell lymphoma (DLBCL). Leuk. Res. 2013, 37, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.H.; Wang, S.L.; Zhao, J.T.; Lin, Z.J.; Chen, L.Y.; Su, R.; Xie, S.T.; Carter, B.Z.; Xu, B. miR-150 exerts antileukemia activity in vitro and in vivo through regulating genes in multiple pathways. Cell Death. Dis. 2016, 7, e2371. [Google Scholar] [CrossRef]
- Undi, R.B.; Kandi, R.; Gutti, R.K. MicroRNAs as Haematopoiesis Regulators. Adv. Hematol. 2013, 2013, 695754. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.; van den Bosch, M.; Wegrzyn, J.; Parker, J.; Ibrahim, R.; Slowski, K.; Chang, L.; Martinez-Høyer, S.; Condorelli, G.; Boldin, M.; et al. miR-143/145 differentially regulate hematopoietic stem and progenitor activity through suppression of canonical TGFβ signaling. Nat. Commun. 2018, 9, 2418. [Google Scholar] [CrossRef]
- Starczynowski, D.T.; Kuchenbauer, F.; Argiropoulos, B.; Sung, S.; Morin, R.; Muranyi, A.; Hirst, M.; Hogge, D.; Marra, M.; Wells, R.A.; et al. Identification of miR-145 and miR-146a as mediators of the 5q-syndrome phenotype. Nat. Med. 2010, 16, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Getaneh, Z.; Asrie, F.; Melku, M. MicroRNA profiles in B-cell non-Hodgkin lymphoma. EJIFCC 2019, 30, 195–214. [Google Scholar]
- Chung, S.S.; Hu, W.; Park, C.Y. The Role of MicroRNAs in Hematopoietic Stem Cell and Leukemic Stem Cell Function. Ther. Adv. Hematol. 2011, 2, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; He, M.; Chen, L.; Chen, C.; Zheng, J.; Cai, Z. The loss of miR-26a-mediated post-transcriptional regulation of cyclin E2 in pancreatic cancer cell proliferation and decreased patient survival. PLoS ONE 2013, 8, e76450. [Google Scholar] [CrossRef]
- Zhao, X.X.; Yuan, Q.Z.; Mu, D.P.; Sun, D.W.; Bo, Q.A.; Pan, G.Z.; Li, G.Q.; Cui, T.; Ding, P.P.; You, F.P.; et al. MicroRNA-26a inhibits proliferation by targeting high mobility group AT-hook 1 in breast cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 368–373. [Google Scholar] [PubMed]
- Chen, L.; Zheng, J.; Zhang, Y.; Yang, L.X.; Wang, J.Q.; Ni, J.; Cui, D.X.; Yu, C.Q.; Cai, Z.L. Tumor-specific expression of microRNA-26a suppresses human hepatocellular carcinoma growth via cyclin-dependent and -independent pathways. Mol. Ther. 2011, 19, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Zheng, S.; Xu, Y.; Xu, A.; Chen, B.; Wen, Y. Loss of miR-26a-5p promotes proliferation, migration, and invasion in prostate cancer through negatively regulating SERBP1. Tumour Biol. 2016, 37, 12843–12854. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Guo, W.; Zhao, Y.; Wang, Y.; Zha, R.; Ding, J.; Liang, L.; Hu, J.; Shen, H.; Chen, Z.; et al. MicroRNA-26a acts as a tumor suppressor inhibiting gallbladder cancer cell proliferation by directly targeting HMGA2. Int. J. Oncol. 2014, 44, 2050–2058. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Min, J.; Sui, X.; Qian, Y.; Liu, Y.; Liu, Z.; Zhou, H.; Li, X.; Gong, Y. MicroRNA-26a-5p and microRNA-23b-3p up-regulate peroxiredoxin III in acute myeloid leukemia. Leukemia Lymphoma 2015, 56, 460–471. [Google Scholar]
- Farina, F.; Inguscio, A.; Kunderfranco, P.; Cortesi, A.; Elia, L.; Quintavalle, M. MicroRNA-26a/cyclin-dependent kinase 5 axis controls proliferation, apoptosis and in vivo tumor growth of diffuse large B-cell lymphoma cell lines. Cell Death Dis. 2017, 8, e2890. [Google Scholar] [CrossRef]
- Karaczyn, A.A.; Jachimowicz, E.; Kohli, J.S.; Sathyanarayana, P. Microrna 199b Regulates Mouse Hematopoietic Stem Cells Maintenance. Blood 2019, 134 (Suppl. 1), 3704. [Google Scholar] [CrossRef]
- Li, Y.; Bai, H.; Zhang, Z.; Li, W.; Dong, L.; Wei, X.; Ma, Y.; Zhang, J.; Yu, J.; Sun, G.; et al. The up-regulation of miR-199b-5p in erythroid differentiation is associated with GATA-1 and NF-E2. Mol. Cells 2014, 37, 21–39. [Google Scholar] [CrossRef]
- Houshmand, M.; Nakhlestani, M.; Soleimani, M.; Hamidieh, A.A.; Abroun, S.; Nikougoftar, Z.M. MicroRNA Microarray Profiling during Megakaryocyte Differentiation of Cord Blood CD133+ Hematopoietic Stem Cells. Cell J. 2018, 20, 195–203. [Google Scholar]
- Luo, S.; Liu, Y.; Liang, G.; Zhao, M.; Wu, H.; Liang, Y.; Qiu, X.; Tan, Y.; Dai, Y.; Yung, S.; et al. The role of microRNA-1246 in the regulation of B cell activation and the pathogenesis of systemic lupus erythematosus. Clin. Epigenet. 2015, 7, 24. [Google Scholar] [CrossRef]
- Azzouzi, I.; Moest, H.; Winkler, J.; Fauchere, J.-C.; Gerber, A.P.; Wollscheid, B.; Stoffel, M.; Schmugge, M.; Speer, O. MicroRNA-96 directly inhibits γ-globin expression in human erythropoiesis. PLoS ONE 2011, 6, e22838. [Google Scholar] [CrossRef] [PubMed]
- Lessard, S.; Beaudoin, M.; Orkin, S.H.; Bauer, D.E.; Lettre, G. 14q32 and let-7 microRNAs regulate transcriptional networks in fetal and adult human erythroblasts. Hum. Mol. Genet. 2018, 27, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Kontos, C.K.; Pappa, V.; Vasilatou, D.; Pavlou, M.A.S.; Kontsioti, F.; Papageorgiou, S.G.; Tsiotra, P.; Economopoulou, C.; Bakou, V.; Papageorgiou, E.; et al. Let-7a, Mir-17 and Mir-20a Expression Levels in CD34+ Bone Marrow Cells of Patients with Myelodysplastic Syndromes (MDS) Are Associated with Established Prognostic Factors, Supporting Their Implication in the Pathogenesis of the Disease. Blood 2011, 118, 3792. [Google Scholar] [CrossRef]
- Choi, Y.; Hur, E.H.; Moon, J.H.; Goo, B.K.; Choi, D.R.; Lee, J.H. Expression and prognostic significance of microRNAs in Korean patients with myelodysplastic syndrome. Korean J. Intern. Med. 2019, 34, 390–400. [Google Scholar] [CrossRef]
- Zuo, Z.; Calin, G.A.; de Paula, H.M.; Medeiros, L.J.; Fernandez, M.H.; Shimizu, M.; Garcia-Manero, G.; Bueso-Ramos, C.E. Circulating microRNAs let-7a and миPHK-16 predict progression-free survival and overall survival in patients with myelodysplastic syndrome. Blood 2011, 118, 413–415. [Google Scholar] [CrossRef]
- Leitão, A.L.; Costa, M.C.; Enguita, F.J. A guide for miRNA target prediction and analysis using web-based applications. Methods Mol. Biol. 2014, 182, 265–277. [Google Scholar]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, 460–466. [Google Scholar] [CrossRef]
- Polivka, J.; Janku, F. Molecular targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol. Ther. 2014, 142, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Farhan, M.; Wang, H.; Gaur, U.; Little, P.J.; Xu, J.; Zheng, W. FOXO Signaling Pathways as Therapeutic Targets in Cancer. Int. J. Biol. Sci. 2017, 13, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, I.; Fernando, J.; Mainez, J.; Sancho, P. TGF-beta signaling in cancer treatment. Curr. Pharm. Des. 2014, 20, 2934–2947. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wei, X.; Li, W.; Udan, R.S.; Yang, Q.; Kim, J.; Xie, J.; Ikenoue, T.; Yu, J.; Li, L.; et al. Inactivation of YAP oncoprotein by the Hippo pathway is involved in cell contact inhibition and tissue growth control. Genes Dev. 2007, 21, 2747–2761. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| NHL (−MD) vs. NCBD | p-Value | NHL (+MD) (−A) vs. NCBD | p-Value | NHL (+MD) (+A) vs. NCBD | p-Value | MDS vs. NCBD | p-Value | |
|---|---|---|---|---|---|---|---|---|
| let-7a-5p | 1.35 | 0.122208 | 3.78 | 0.000002 | 3.25 | 0.002901 | 2.14 | 0.000021 |
| miR-16-5p | −1.23 | 0.647554 | 1.08 | 0.333132 | 1.37 | 0.555872 | −1.12 | 0.985981 |
| miR-155-5p | 1.53 | 0.053346 | 1.54 | 0.739425 | 1.44 | 0.444910 | 1.04 | 0.783056 |
| miR-26a-5p | −51.97 | 7.4 × 10−18 | −54.83 | 3.83 × 10−11 | −44.82 | 1.4 × 10−9 | −50.16 | 8.2 × 10−17 |
| miR-181a-5p | −1.98 | 0.000162 | −1.58 | 0.131210 | −1.18 | 0.388964 | −2.13 | 0.000706 |
| miR-185-5p | −4.50 | 0.000003 | −1.04 | 0.567289 | −1.79 | 0.097465 | −2.67 | 0.000002 |
| miR-96-5p | −4.43 | 0.000001 | −2.48 | 0.067222 | −2.73 | 0.003131 | −1.77 | 0.102073 |
| miR-1246 | −2.38 | 0.000421 | −2.18 | 0.175334 | −1.48 | 0.029906 | −3.48 | 0.007841 |
| miR-199b-5p | −1.98 | 0.004179 | −3.39 | 0.000225 | −2.59 | 0.000908 | −1.79 | 0.000355 |
| miR-126-3p | −1.06 | 0.497818 | −1.16 | 0.884774 | −1.10 | 0.839624 | −1.25 | 0.237218 |
| miR-451a | 1.29 | 0.720385 | 1.01 | 0.739425 | −2.22 | 0.239566 | −1.54 | 0.065864 |
| miR-145-5p | −8.31 | 0.000000 | −5.05 | 0.000002 | −2.27 | 0.008440 | −4.35 | 0.000000 |
| miR-150-5p | −1.66 | 0.007797 | −1.67 | 0.000426 | −2.27 | 0.024672 | −2.54 | 0.002474 |
| KEGG Pathway | Genes in the Pathway, Total | p-Value |
|---|---|---|
| Signaling pathway | ||
| Hippo signaling pathway (hsa04390) | 86 | 2.3 × 10−14 |
| TGF-beta signaling pathway (hsa04350) | 54 | 1.4 × 10−11 |
| Signaling pathways regulating pluripotency of stem cells (hsa04550) | 77 | 1.2 × 10−8 |
| FoxO signaling pathway (hsa04068) | 72 | 4.2 × 10−5 |
| HIF-1 signaling pathway (hsa04066) | 58 | 5.8 × 10−5 |
| ErbB signaling pathway (hsa04012) | 43 | 2 × 10−3 |
| MAPK signaling pathway (hsa04010) | 107 | 5 × 10−3 |
| PI3K-Akt signaling pathway (hsa04151) | 142 | 1 × 10−2 |
| mTOR signaling pathway (hsa04150) | 32 | 2 × 10−2 |
| Wnt signaling pathway (hsa04310) | 58 | 4 × 10−2 |
| Cell biology | ||
| Cell cycle (hsa04110) | 75 | 3.5 × 10−11 |
| Apoptosis (hsa04210) | 38 | 4 × 10−2 |
| RNA transport (hsa03013) | 73 | 2 × 10−2 |
| RNA degradation (hsa03018) | 39 | 3 × 10−2 |
| Cancer-associated pathways | ||
| Transcriptional misregulation in cancer (hsa05202) | 85 | 1.1 × 10−6 |
| Pathways in cancer (hsa05200) | 175 | 3.6 × 10−5 |
| Proteoglycans in cancer (hsa05205) | 111 | 8.6 × 10−16 |
| Chronic myeloid leukemia (hsa05220) | 39 | 1 × 10−3 |
| Bladder cancer (hsa05219) | 25 | 2 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veryaskina, Y.A.; Titov, S.E.; Kovynev, I.B.; Pospelova, T.I.; Zhimulev, I.F. The miRNA Profile in Non-Hodgkin’s Lymphoma Patients with Secondary Myelodysplasia. Cells 2020, 9, 2318. https://doi.org/10.3390/cells9102318
Veryaskina YA, Titov SE, Kovynev IB, Pospelova TI, Zhimulev IF. The miRNA Profile in Non-Hodgkin’s Lymphoma Patients with Secondary Myelodysplasia. Cells. 2020; 9(10):2318. https://doi.org/10.3390/cells9102318
Chicago/Turabian StyleVeryaskina, Yuliya Andreevna, Sergei Evgenievich Titov, Igor Borisovich Kovynev, Tatiana Ivanovna Pospelova, and Igor Fyodorovich Zhimulev. 2020. "The miRNA Profile in Non-Hodgkin’s Lymphoma Patients with Secondary Myelodysplasia" Cells 9, no. 10: 2318. https://doi.org/10.3390/cells9102318
APA StyleVeryaskina, Y. A., Titov, S. E., Kovynev, I. B., Pospelova, T. I., & Zhimulev, I. F. (2020). The miRNA Profile in Non-Hodgkin’s Lymphoma Patients with Secondary Myelodysplasia. Cells, 9(10), 2318. https://doi.org/10.3390/cells9102318