Nucleoporin 62-Like Protein is Required for the Development of Pharyngeal Arches through Regulation of Wnt/β-Catenin Signaling and Apoptotic Homeostasis in Zebrafish

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry
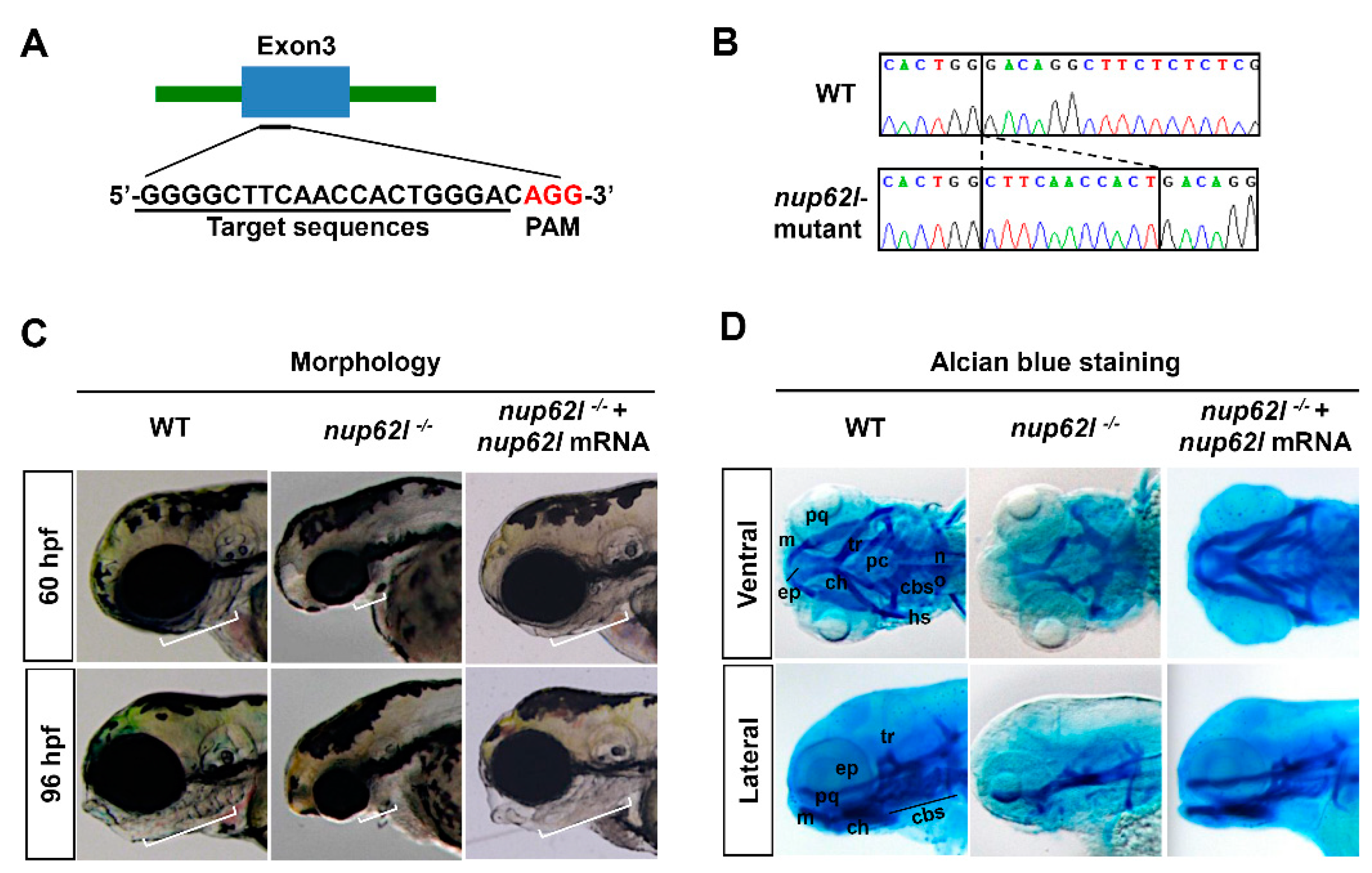
2.2. CRISPR/Cas9-Mediated Gene Knockout
2.3. Constructs, Antisense Morpholino (MO), mRNA Microinjection and Chemical Treatments
2.4. Quantitative Real-Time PCR (qRT–PCR)
2.5. Whole-Mount In Situ Hybridization (WISH)
2.6. Alcian Blue Staining
2.7. TUNEL Assay and Detection of Caspase 3 Activity
3. Results
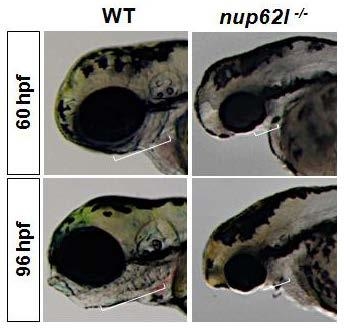
3.1. Nup62l Depletion Impaired the Development of PA
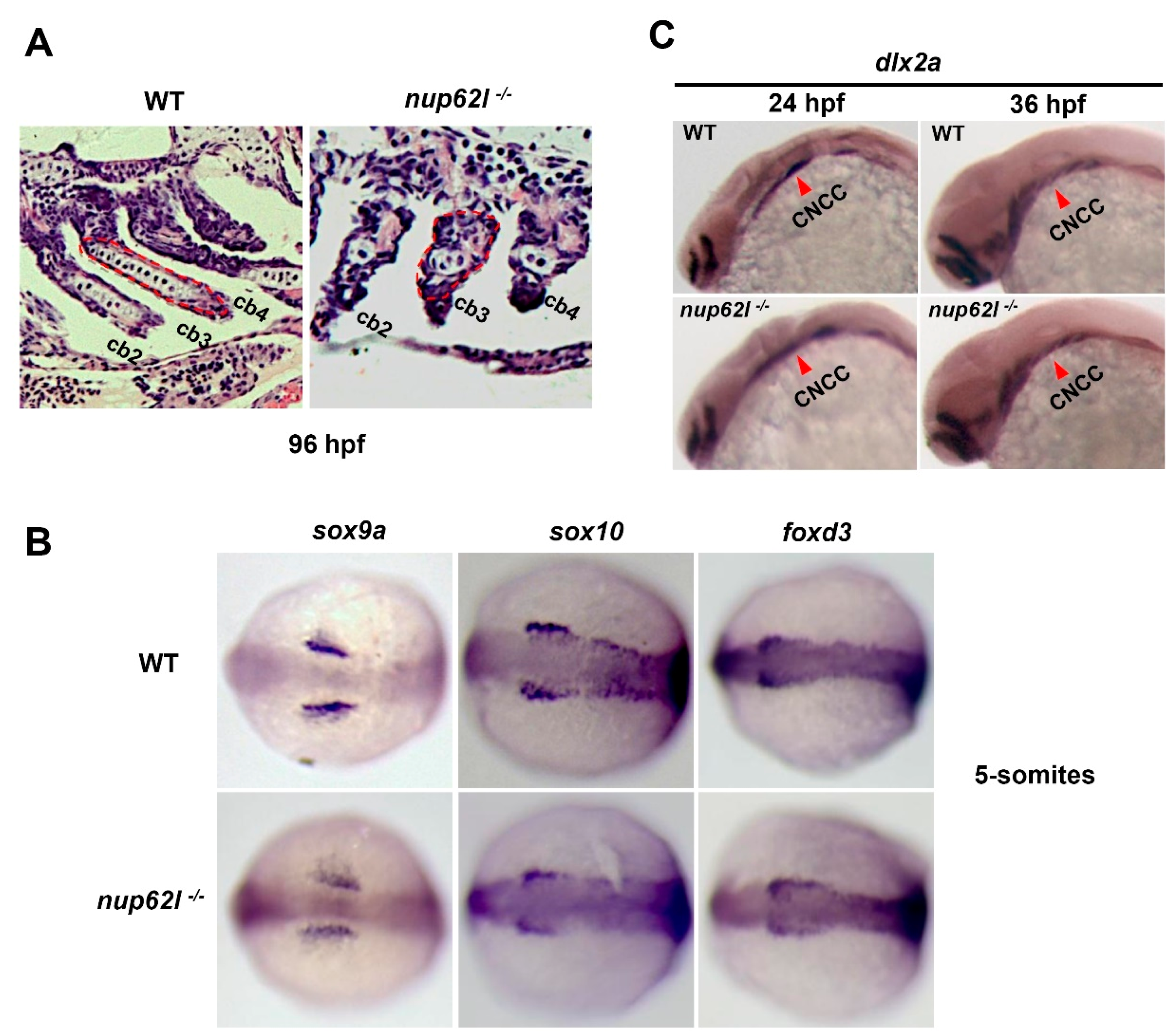
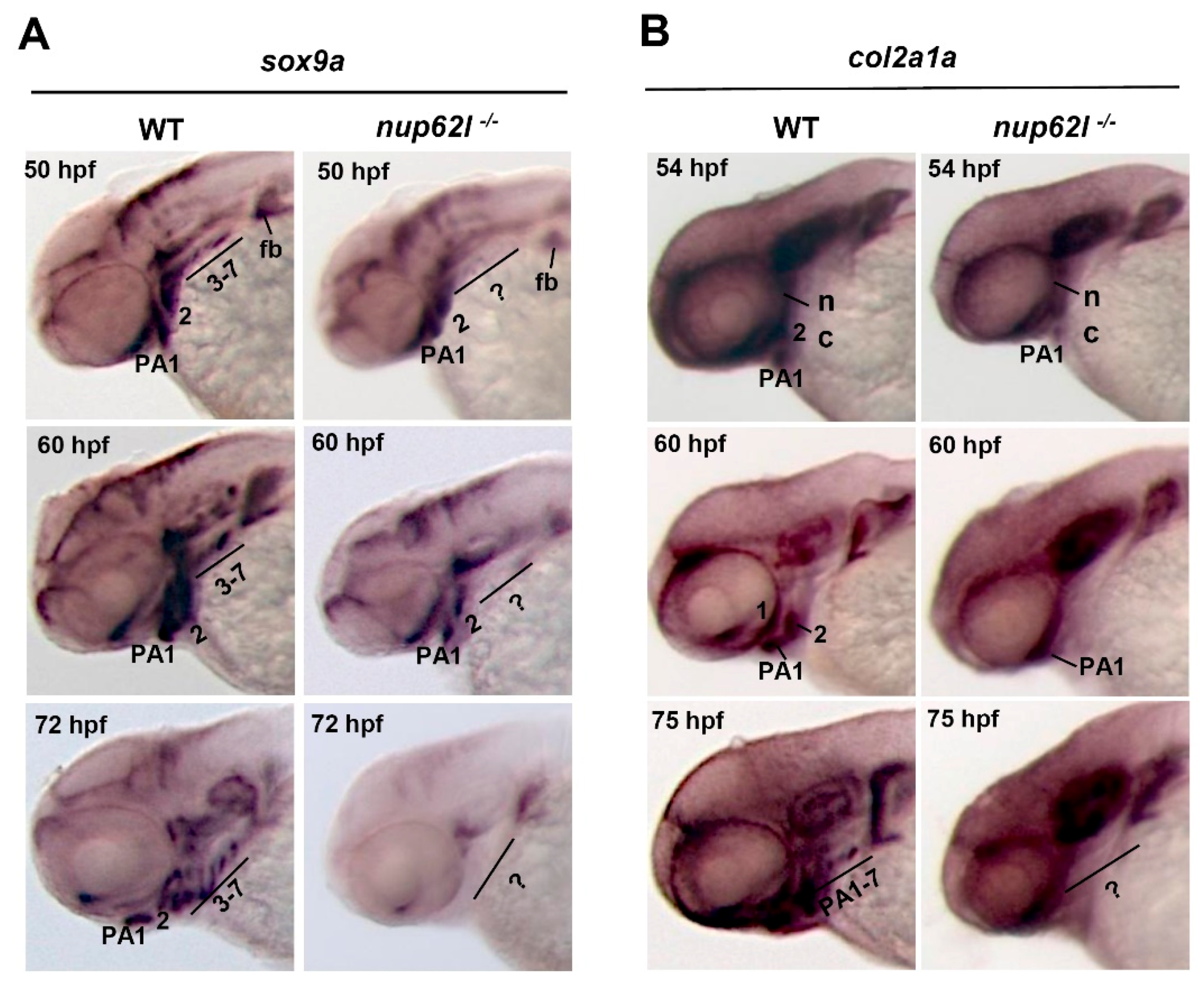
3.2. Nup62l Loss-of-Function Disrupted Precondrogenic Condensation and Chondrogenic Differentiation during Pharyngeal Cartilage Formation
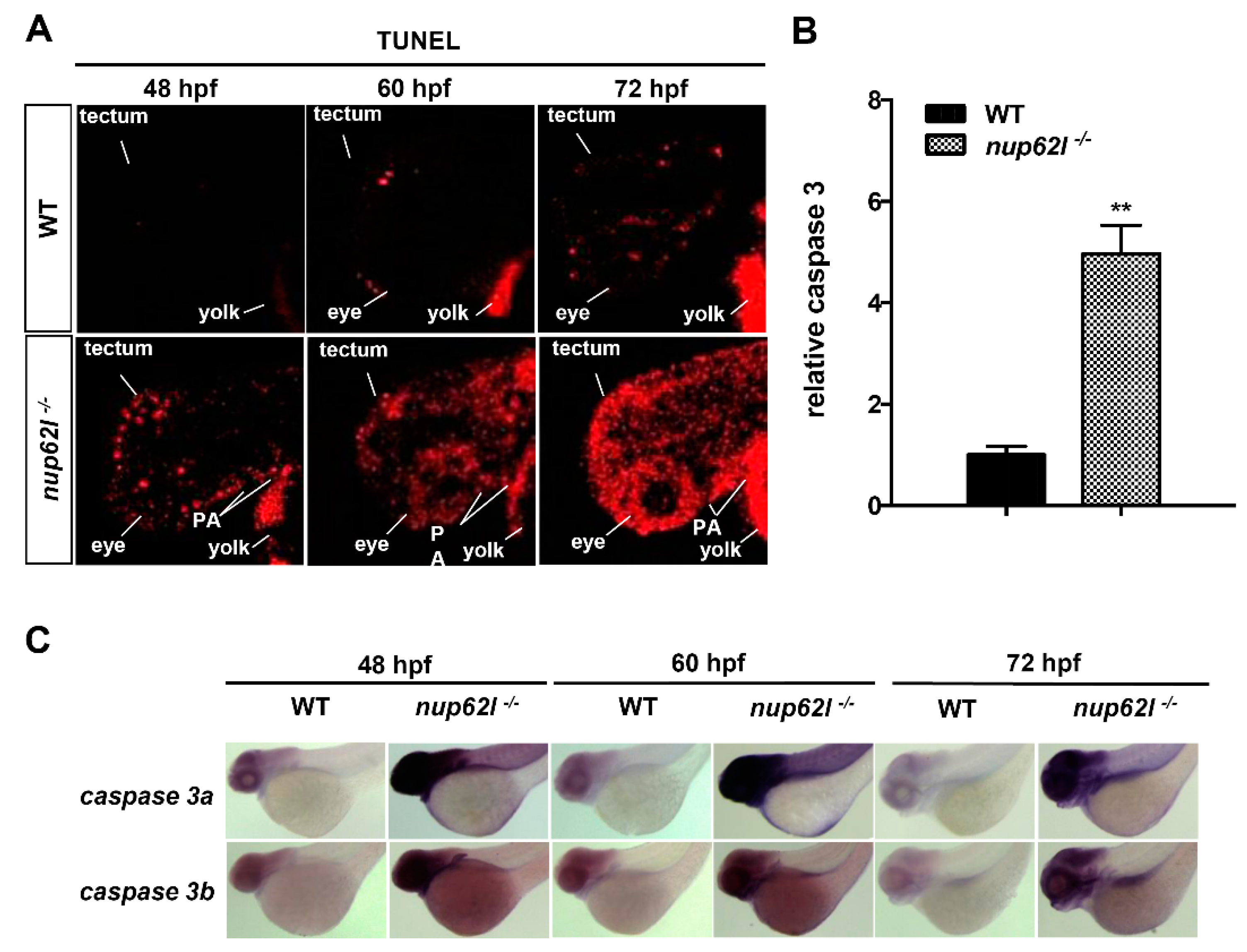
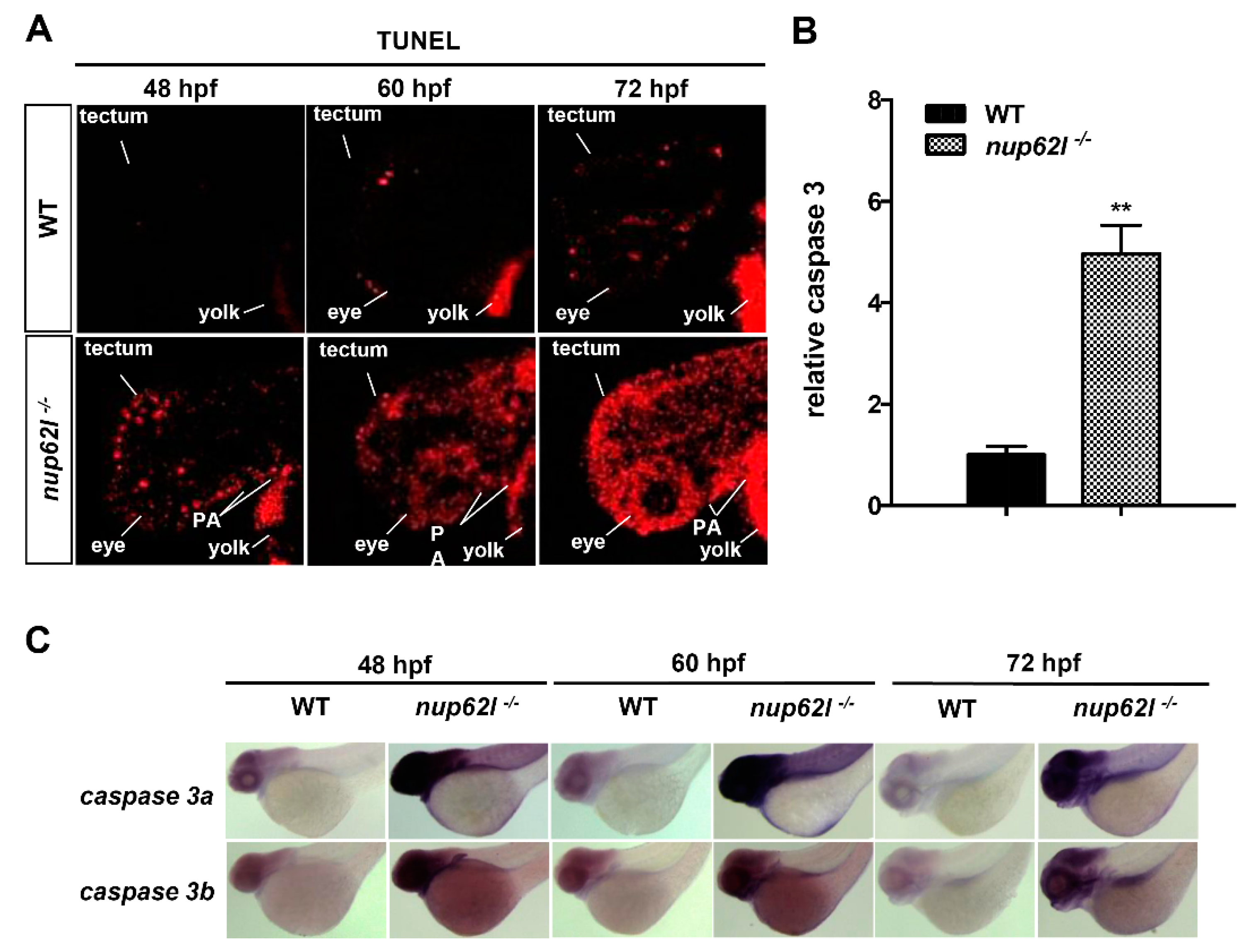
3.3. Extensive Apoptosis Occurred in the Defective PA of Nup62l-Mutants
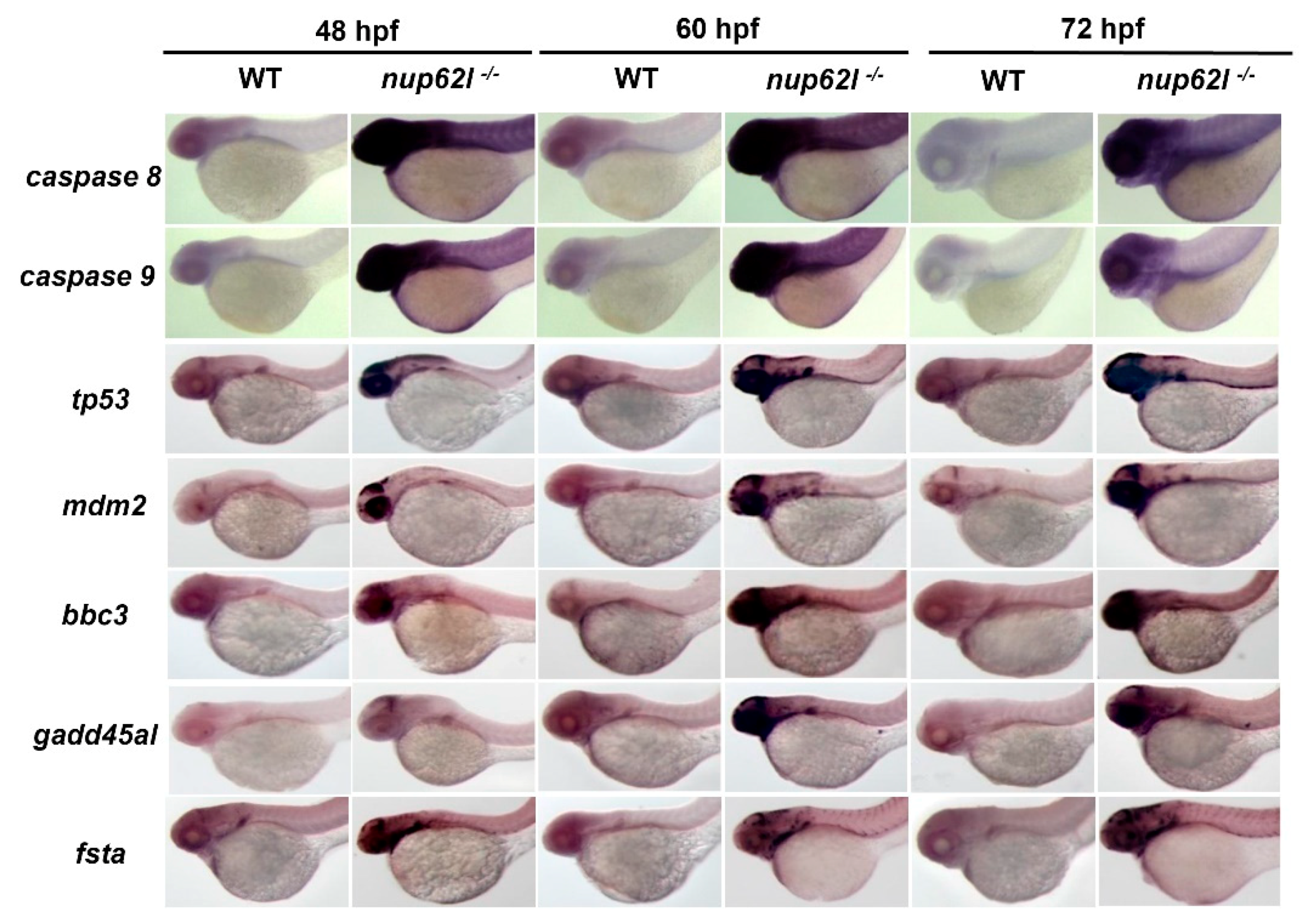
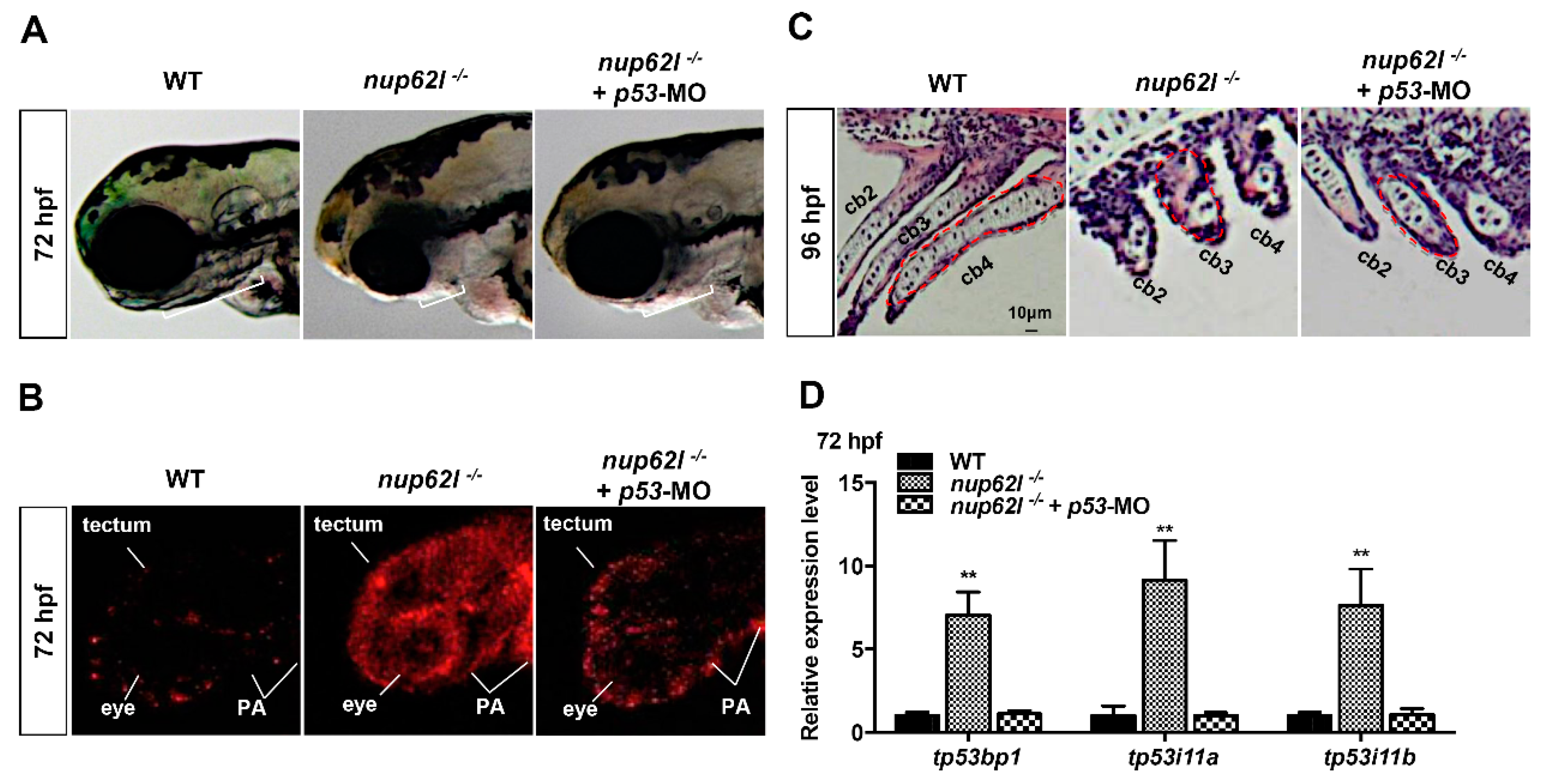
3.4. Activation of the p53-Driven Apoptotic Pathway Contributed to the Impaired Formation of PA in Nup62l-Mutants
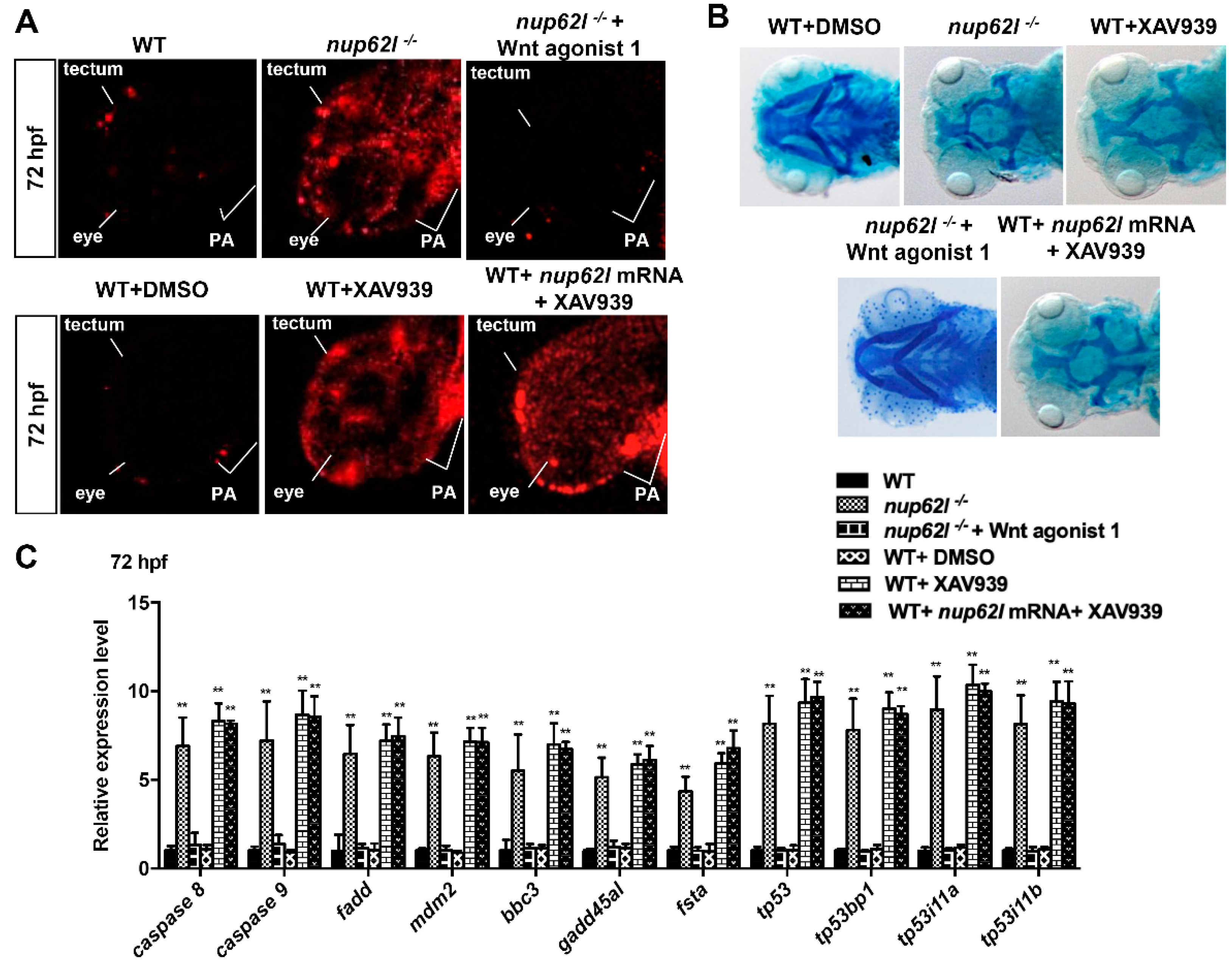
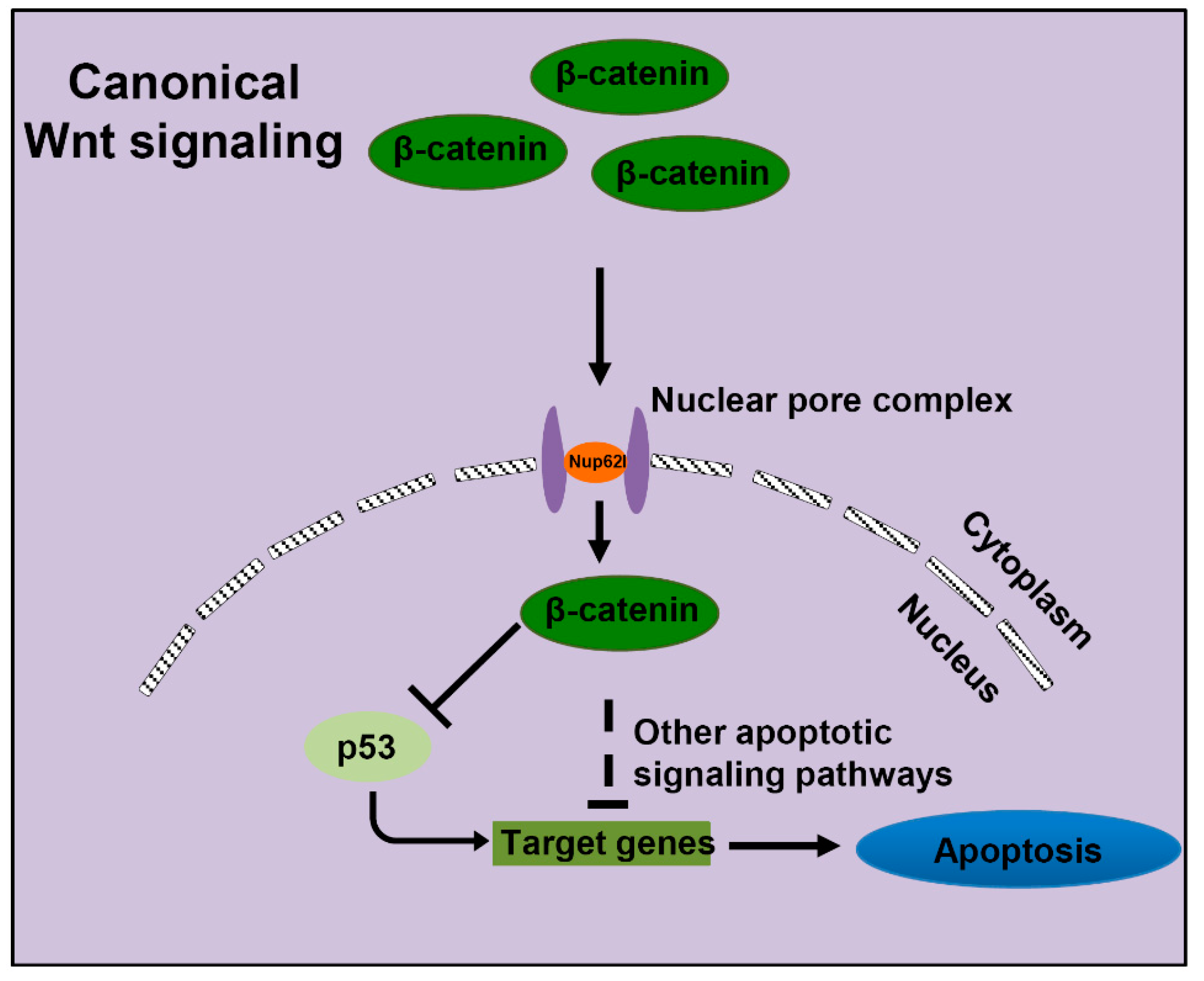
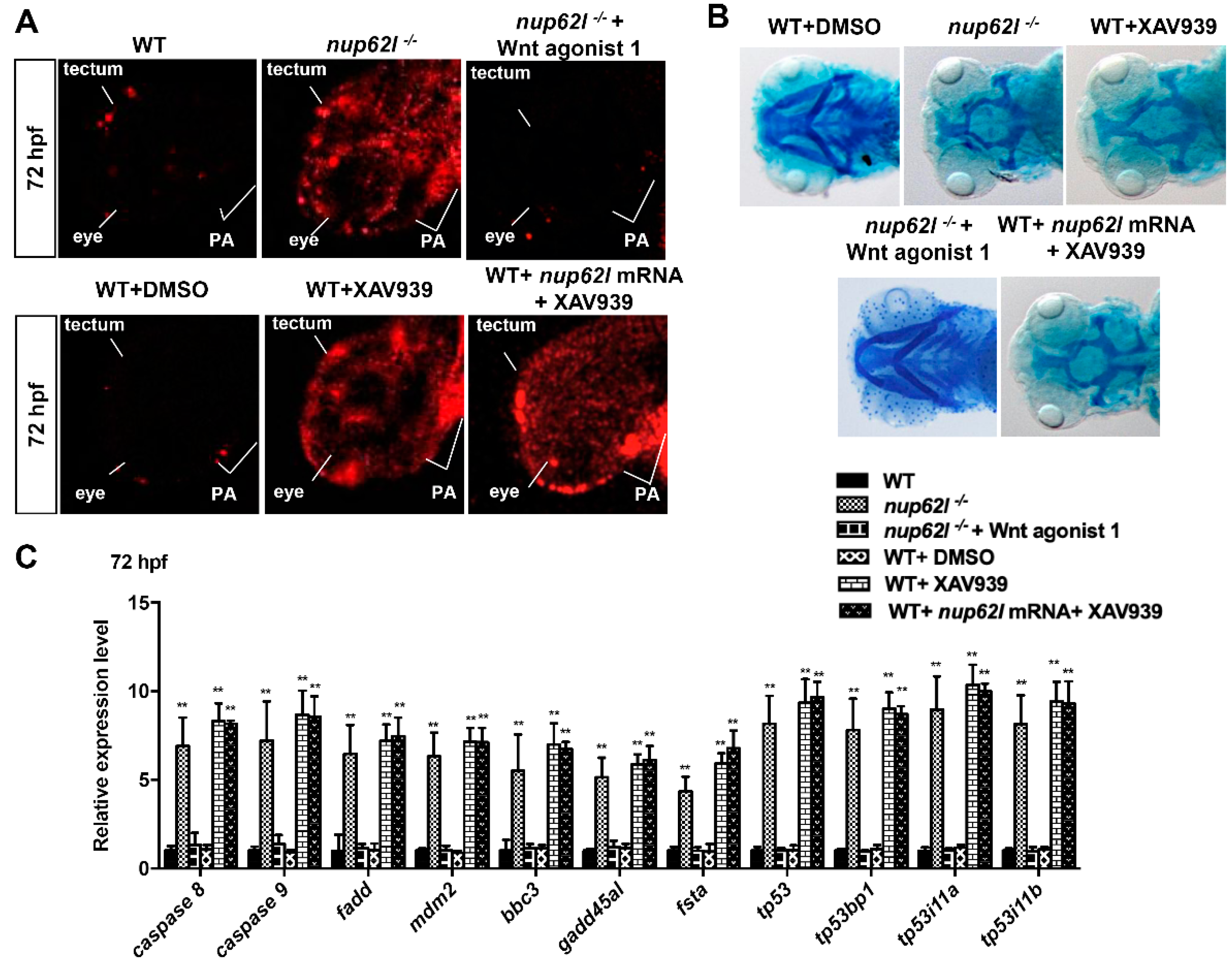
3.5. Extensive Apoptosis Occurring in PA of Nup62l-Mutants is Attributable to Inactivation of Wnt/β-Catenin Signaling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ning, G.; Liu, X.; Dai, M.; Meng, A.; Wang, Q. MicroRNA-92a Upholds Bmp Signaling by Targeting noggin3 during Pharyngeal Cartilage Formation. Dev. Cell 2013, 24, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Clouthier, D.E.; Garcia, E.; Schilling, T.F. Regulation of Facial Morphogenesis by Endothelin Signaling: Insights from Mice and Fish. Am. J. Med Genet. Part A 2010, 152, 2962–2973. [Google Scholar] [CrossRef]
- Schilling, T.F.; Kimmel, C.B. Segment and cell type lineage restrictions during pharyngeal arch development in the zebrafish embryo. Development 1994, 120, 483–494. [Google Scholar]
- Kimmel, C.B.; Schilling, T.F.; Hatta, K. Patterning of Body Segments of the Zebrafish Embryo. Curr. Top. Dev. Biol. 1991, 25, 77–110. [Google Scholar]
- Noden, D.M. Interactions and Fates of Avian Craniofacial Mesenchyme. Development 1988, 103, 121–140. [Google Scholar]
- Schilling, T.F.; Kimmel, C.B. Musculoskeletal patterning in the pharyngeal segments of the zebrafish embryo. Development 1997, 124, 2945–2960. [Google Scholar]
- Thorogood, P. Morphogenesis of Cartilage. In Cartilage; Hall, B.K., Ed.; Academic Press: New York, NY, USA, 1983; Volume 2, pp. 223–254. [Google Scholar]
- Hall, B.; Miyake, T. The membranous skeleton: The role of cell condensations in vertebrate skeletogenesis. Brain Struct. Funct. 1992, 186, 107–124. [Google Scholar] [CrossRef]
- Piotrowski, T.; Schilling, T.F.; Brand, M.; Jiang, Y.J.; Heisenberg, C.P.; Beuchle, D.; Grandel, H.; Van Eeden, F.J.; Furutani-Seiki, M.; Granato, M.; et al. Jaw and branchial arch mutants in zebrafish II: anterior arches and cartilage differentiation. Development 1996, 123, 345–356. [Google Scholar]
- Yelick, P.C.; Schilling, T.F. Molecular Dissection of Craniofacial Development Using Zebrafish. Crit. Rev. Oral Boil. Med. 2002, 13, 308–322. [Google Scholar] [CrossRef]
- Kurihara, Y.; Kurihara, H.; Maemura, K.; Kuwaki, T.; Kumada, M.; Yazaki, Y. Impaired Development of the Thyroid and Thymus in Endothelin-1 Knockout Mice. J. Cardiovasc. Pharmacol. 1995, 26, S13–S16. [Google Scholar] [CrossRef]
- Hong, R. The Digeorge Anomaly (Catch 22, Digeorge/Velocardiofacial Syndrome). Semin. Hematol. 1998, 35, 282–290. [Google Scholar]
- A Dutton, K.; Pauliny, A.; Lopes, S.S.; Elworthy, S.; Carney, T.J.; Rauch, J.; Geisler, R.; Haffter, P.; Kelsh, R.N. Zebrafish colourless encodes sox10 and specifies non-ectomesenchymal neural crest fates. Development 2001, 128, 4113–4125. [Google Scholar]
- Hassinger, D.D.; Mulvihill, J.J.; Chandler, J.B. Aarskog’s syndrome with Hirschsprung’s disease, midgut malrotation, and dental anomalies. J. Med Genet. 1980, 17, 235–238. [Google Scholar] [CrossRef]
- Muenke, M.; Cohen, M.M. Genetic approaches to understanding brain development: Holoprosencephaly as a model. Ment. Retard. Dev. Disabil. Res. Rev. 2000, 6, 15–21. [Google Scholar] [CrossRef]
- Graham, A. Development of the pharyngeal arches. Am. J. Med Genet. 2003, 119, 251–256. [Google Scholar] [CrossRef]
- Bush, J.O.; Jiang, R. Palatogenesis: Morphogenetic and molecular mechanisms of secondary palate development. Development 2012, 139, 828. [Google Scholar] [CrossRef]
- Sperber, S.M.; Dawid, I.B. barx1 is necessary for ectomesenchyme proliferation and osteochondroprogenitor condensation in the zebrafish pharyngeal arches. Dev. Boil. 2008, 321, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.; Tucker, A.S. Fgf and Bmp signals repress the expression of Bapx1 in the mandibular mesenchyme and control the position of the developing jaw joint. Dev. Boil. 2004, 266, 138–150. [Google Scholar] [CrossRef]
- Alexander, C.; Zuniga, E.; Blitz, I.L.; Wada, N.; Le Pabic, P.; Javidan, Y.; Zhang, T.; Cho, K.W.; Crump, J.G.; Schilling, T.F. Combinatorial roles for BMPs and Endothelin 1 in patterning the dorsal-ventral axis of the craniofacial skeleton. Development 2011, 138, 5135–5146. [Google Scholar] [CrossRef] [Green Version]
- Zuniga, E.; Stellabotte, F.; Crump, J.G. Jagged-Notch signaling ensures dorsal skeletal identity in the vertebrate face. Development 2010, 137, 1843–1852. [Google Scholar] [CrossRef] [Green Version]
- Carroll, T.J.; Park, J.-S.; Hayashi, S.; Majumdar, A.; McMahon, A.P. Wnt9b Plays a Central Role in the Regulation of Mesenchymal to Epithelial Transitions Underlying Organogenesis of the Mammalian Urogenital System. Dev. Cell 2005, 9, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Niemann, S.; Zhao, C.; Pascu, F.; Stahl, U.; Aulepp, U.; Niswander, L.; Weber, J.L.; Müller, U. Homozygous WNT3 Mutation Causes Tetra-Amelia in a Large Consanguineous Family. Am. J. Hum. Genet. 2004, 74, 558–563. [Google Scholar] [CrossRef] [Green Version]
- Brault, V.; Moore, R.; Kutsch, S. Inactivation of the (Β)-Catenin Gene by Wnt1-Cre-Mediated Deletion Results in Dramatic Brain Malformation and Failure of Craniofacial Development. Development 2001, 128, 1253–1264. [Google Scholar]
- Song, L.; Li, Y.; Wang, K.; Wang, Y.-Z.; Molotkov, A.; Gao, L.; Zhao, T.; Yamagami, T.; Wang, Y.; Gan, Q.; et al. Lrp6-mediated canonical Wnt signaling is required for lip formation and fusion. Development 2009, 136, 3161–3171. [Google Scholar] [CrossRef] [Green Version]
- Reinhold, M.I.; Kapadia, R.M.; Liao, Z. The Wnt-Inducible Transcription Factor Twist1 Inhibits Chondrogenesis. J. Biol. Chem. 2006, 281, 1381–1388. [Google Scholar] [CrossRef]
- Choe, C.P.; Collazo, A.; Trinh, L.A.; Pan, L.; Moens, C.B.; Crump, J.G. Wnt-dependent epithelial transitions drive pharyngeal pouch formation. Dev. Cell 2013, 24, 296–309. [Google Scholar] [CrossRef]
- Curtin, E.; Hickey, G.; Kamel, G.; Davidson, A.J.; Liao, E.C. Zebrafish wnt9a is expressed in pharyngeal ectoderm and is required for palate and lower jaw development. Mech. Dev. 2011, 128, 104–115. [Google Scholar] [CrossRef]
- Heisenberg, C.P.; Tada, M.; Rauch, G.J. Silberblick/Wnt11 Mediates Convergent Extension Movements During Zebrafish Gastrulation. Nature 2000, 405, 76–81. [Google Scholar] [CrossRef]
- Wu, B.-T.; Wen, S.-H.; Hwang, S.-P.L.; Huang, C.-J.; Kuan, Y.-S. Control of Wnt5b secretion by wntless modulates chondrogenic cell proliferation through fine-tuning fgf3 expression. J. Cell Sci. 2015, 128, 2328–2339. [Google Scholar] [CrossRef]
- Davuluri, G.; Gong, W.; Yusuff, S.; Lorent, K.; Muthumani, M.; Dolan, A.C.; Pack, M. Mutation of the Zebrafish Nucleoporin elys Sensitizes Tissue Progenitors to Replication Stress. PLoS Genet. 2008, 4, 1000240. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, S.; Han, Y.; Zhao, X.; Zhao, L.; Tian, T.; Tong, J.; Xu, P.; Xiong, C.; Meng, A. Loss of Zygotic NUP107 Protein Causes Missing of Pharyngeal Skeleton and Other Tissue Defects with Impaired Nuclear Pore Function in Zebrafish Embryos*. J. Boil. Chem. 2012, 287, 38254–38264. [Google Scholar] [CrossRef]
- Hu, T.; Guan, T.; Gerace, L. Molecular and functional characterization of the p62 complex, an assembly of nuclear pore complex glycoproteins. J. Cell Boil. 1996, 134, 589–601. [Google Scholar] [CrossRef]
- Frey, S.; Görlich, D. A Saturated FG-Repeat Hydrogel Can Reproduce the Permeability Properties of Nuclear Pore Complexes. Cell 2007, 130, 512–523. [Google Scholar] [CrossRef] [Green Version]
- Van Impe, K.; Hübert, T.; De Corte, V.; Vanloo, B.; Boucherie, C.; Vandekerckhove, J.; Gettemans, J. A New Role for Nuclear Transport Factor 2 and Ran: Nuclear Import of CapG. Traffic 2008, 9, 695–707. [Google Scholar] [CrossRef]
- Basel-Vanagaite, L.; Muncher, L.; Straussberg, R. Mutated Nup62 Causes Autosomal Recessive Infantile Bilateral Striatal Necrosis. Ann. Neurol. 2006, 60, 214–222. [Google Scholar] [CrossRef]
- Yang, X.; Gu, Q.; Lin, L.; Li, S.; Zhong, S.; Li, Q.; Cui, Z. Nucleoporin 62-Like Protein Activates Canonical Wnt Signaling through Facilitating the Nuclear Import of β-Catenin in Zebrafish. Mol. Cell. Boil. 2015, 35, 1110–1124. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Jao, L.-E.; Wente, S.R.; Chen, W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc. Natl. Acad. Sci. 2013, 110, 13904–13909. [Google Scholar] [CrossRef] [Green Version]
- Langheinrich, U.; Hennen, E.; Stott, G.; Vacun, G. Zebrafish as a Model Organism for the Identification and Characterization of Drugs and Genes Affecting p53 Signaling. Curr. Boil. 2002, 12, 2023–2028. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.; Yang, X.; He, X.; Li, Q.; Cui, Z. Generation and Characterization of a Transgenic Zebrafish Expressing the Reverse Tetracycline Transactivator. J. Genet. Genom. 2013, 40, 523–531. [Google Scholar] [CrossRef]
- Gu, Q.; Yang, X.; Lin, L.; Li, S.; Li, Q.; Zhong, S.; Peng, J.; Cui, Z. Genetic ablation of solute carrier family 7a3a leads to hepatic steatosis in zebrafish during fasting. Hepatol. 2014, 60, 1929–1941. [Google Scholar] [CrossRef]
- Diekmann, H.; Anichtchik, O.; Fleming, A.; Futter, M.; Goldsmith, P.; Roach, A.; Rubinsztein, D.C. Decreased BDNF Levels Are a Major Contributor to the Embryonic Phenotype of Huntingtin Knockdown Zebrafish. J. Neurosci. 2009, 29, 1343–1349. [Google Scholar] [CrossRef] [Green Version]
- Schilling, T.F.; Piotrowski, T.; Grandel, H.; Brand, M.; Heisenberg, C.P.; Jiang, Y.J.; Beuchle, D.; Hammerschmidt, M.; A Kane, D.; Mullins, M.C.; et al. Jaw and branchial arch mutants in zebrafish I: branchial arches. Dev. 1996, 123, 329–344. [Google Scholar]
- Köntges, G.; Lumsden, A. Rhombencephalic neural crest segmentation is preserved throughout craniofacial ontogeny. Dev. 1996, 122, 3229–3242. [Google Scholar]
- Akimenko, M.; Ekker, M.; Wegner, J.; Lin, W.; Westerfield, M. Combinatorial expression of three zebrafish genes related to distal- less: part of a homeobox gene code for the head. J. Neurosci. 1994, 14, 3475–3486. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.-L. A pair of Sox: distinct and overlapping functions of zebrafish sox9 co-orthologs in craniofacial and pectoral fin development. Dev. 2005, 132, 1069–1083. [Google Scholar] [CrossRef] [Green Version]
- Chiang, E.F.-L.; Pai, C.I.; Wyatt, M. Two Sox9 Genes on Duplicated Zebrafish Chromosomes: Expression of Similar Transcription Activators in Distinct Sites. Dev. Biol. 2001, 231, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Schwend, T.; Ahlgren, S.C. Zebrafish con/disp1 reveals multiple spatiotemporal requirements for Hedgehog-signaling in craniofacial development. BMC Dev. Boil. 2009, 9, 59. [Google Scholar] [CrossRef]
- Vandenberg, P.; Khillan, J.S.; Prockop, D.J. Expression of a Partially Deleted Gene of Human Type Ii Procollagen (Col2a1) in Transgenic Mice Produces a Chondrodysplasia. Proc. Natl. Acad. Sci. 1991, 88, 7640–7644. [Google Scholar] [CrossRef]
- Yan, Y.-L.; Hatta, K.; Riggleman, B.; Postlethwait, J.H. Expression of a type II collagen gene in the zebrafish embryonic axis. Dev. Dyn. 1995, 203, 363–376. [Google Scholar] [CrossRef]
- Yan, Y.-L.; Miller, C.T.; Nissen, R.M.; Singer, A.; Liu, D.; Kirn, A.; Draper, B.; Willoughby, J.; A Morcos, P.; Amsterdam, A.; et al. A zebrafish sox9 gene required for cartilage morphogenesis. Development 2002, 129, 5065–5079. [Google Scholar]
- Wolf, B.B.; Schuler, M.; Echeverri, F.; Green, D.R. Caspase-3 Is the Primary Activator of Apoptotic DNA Fragmentation via DNA Fragmentation Factor-45/Inhibitor of Caspase-activated DNase Inactivation. J. Boil. Chem. 1999, 274, 30651–30656. [Google Scholar] [CrossRef] [Green Version]
- Haupt, S.; Berger, M.; Goldberg, Z.; Haupt, Y. Apoptosis-the P53 Network. J. Cell Sci. 2003, 116, 4077–4085. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/β-Catenin Signaling in Development and Disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef]
- Chen, S.; Guttridge, D.C.; You, Z.; Zhang, Z.; Fribley, A.; Mayo, M.W.; Kitajewski, J.; Wang, C.-Y. WNT-1 Signaling Inhibits Apoptosis by Activating β-Catenin/T Cell Factor–Mediated Transcription. J. Cell Boil. 2001, 152, 87–96. [Google Scholar] [CrossRef]
- Tian, X.-H.; Hou, W.-J.; Fang, Y.; Fan, J.; Tong, H.; Bai, S.-L.; Chen, Q.; Xu, H.; Li, Y. XAV939, a tankyrase 1 inhibitior, promotes cell apoptosis in neuroblastoma cell lines by inhibiting Wnt/β-catenin signaling pathway. J. Exp. Clin. Cancer Res. 2013, 32, 100. [Google Scholar] [CrossRef]
- Liu, J.; Wu, X.; Mitchell, B.; Kintner, C.; Ding, S.; Schultz, P.G. A Small-Molecule Agonist of the Wnt Signaling Pathway. Angew. Chem. Int. Ed. 2005, 44, 1987–1990. [Google Scholar] [CrossRef]
- De Jong–Curtain, T.A.; Parslow, A.C.; Trotter, A.J.; Hall, N.E. Abnormal Nuclear Pore Formation Triggers Apoptosis in the Intestinal Epithelium of Elys-Deficient Zebrafish. Gastroenterology 2009, 136, 902–911. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Miller, C.T.; Kruze, G.; Ullmann, B.; BreMiller, R.A.; Larison, K.D.; Snyder, H.C. The Shaping of Pharyngeal Cartilages during Early Development of the Zebrafish. Dev. Boil. 1998, 203, 245–263. [Google Scholar] [CrossRef] [Green Version]
- Melčák, I.; Hoelz, A.; Blobel, G. Structure of Nup58/45 Suggests Flexible Nuclear Pore Diameter by Intermolecular Sliding. Science 2007, 315, 1729–1732. [Google Scholar] [CrossRef] [Green Version]
- Fukuhara, T.; Sakaguchi, N.; Katahira, J.; Yoneda, Y.; Ogino, K.; Tachibana, T. Functional Analysis of Nuclear Pore Complex Protein Nup62/p62 Using Monoclonal Antibodies. Hybrid. 2006, 25, 51–59. [Google Scholar] [CrossRef]
- Pećina-Šlaus, N. Wnt signal transduction pathway and apoptosis: a review. Cancer Cell Int. 2010, 10, 22. [Google Scholar] [CrossRef]
- Li, F.; Chong, Z.; Maiese, K. Winding through the WNT pathway during cellular development and demise. Histol. Histopathol. 2006, 21, 103–124. [Google Scholar]
- Ellies, D.L.; Church, V.; Francis-West, P.; Lumsden, A. The WNT antagonist cSFRP2 modulates programmed cell death in the developing hindbrain. Development 2000, 127, 5285–5295. [Google Scholar]
- Kozlovsky, N.; Belmaker, R.; Agam, G. GSK-3 and the neurodevelopmental hypothesis of schizophrenia. Eur. Neuropsychopharmacol. 2002, 12, 13–25. [Google Scholar] [CrossRef]
- Honoré, S.M.; Aybar, M.J.; Mayor, R. Sox10 is required for the early development of the prospective neural crest in Xenopus embryos. Dev. Boil. 2003, 260, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Kavitha, K.; Kowshik, J.; Kishore, T.K.K.; Baba, A.B.; Nagini, S. Astaxanthin Inhibits Nf-Κb and Wnt/Β-Catenin Signaling Pathways Via Inactivation of Erk/Mapk and Pi3k/Akt to Induce Intrinsic Apoptosis in a Hamster Model of Oral Cancer. BBA-Gen. Subj. 2013, 1830, 4433–4444. [Google Scholar] [CrossRef]
- Li, H.; Pamukcu, R.; Thomson, W.J. β-Catenin Signaling: Therapeutic Strategies in Oncology. Cancer Boil. Ther. 2002, 1, 621–625. [Google Scholar] [CrossRef]
- Huang, M.; Wang, Y.; Sun, D.; Zhu, H.; Yin, Y.; Zhang, W.; Yang, S.; Quan, L.; Bai, J.; Wang, S.; et al. Identification of genes regulated by Wnt/β-catenin pathway and involved in apoptosis via microarray analysis. BMC Cancer 2006, 6, 221. [Google Scholar] [CrossRef]
- Iwabuchi, K.; Li, B.; Massa, H.F.; Trask, B.J.; Date, T.; Fields, S. Stimulation of p53-mediated Transcriptional Activation by the p53-binding Proteins, 53BP1 and 53BP2. J. Boil. Chem. 1998, 273, 26061–26068. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.-Q.; Cao, E.-H.; Zhang, Y.; Qin, J.-F. A P53 target gene, PIG11, contributes to chemosensitivity of cells to arsenic trioxide. FEBS Lett. 2004, 569, 94–98. [Google Scholar] [CrossRef] [Green Version]
- Papagiannouli, F.; Fuller, M.T.; Lohmann, I. Nuclear Export in Somatic Cyst Cells Controls Cyst Cell-Germline Coordination and Germline Differentiation in the Drosophila Testis. bioRxiv 2018, 452466. [Google Scholar]
- Parrott, B.B.; Chiang, Y.; Hudson, A.; Sarkar, A.; Guichet, A.; Schulz, C. Nucleoporin98-96 Function Is Required for Transit Amplification Divisions in the Germ Line of Drosophila melanogaster. PLOS ONE 2011, 6, e25087. [Google Scholar] [CrossRef]
- Grandi, P.; Schlaich, N.; Tekotte, H.; Hurt, E.C. Functional Interaction of Nic96p with a Core Nucleoporin Complex Consisting of Nsp1p, Nup49p and a Novel Protein Nup57p. EMBO J. 1995, 14, 76–87. [Google Scholar] [CrossRef]
- Nakielny, S.; Shaikh, S.; Burke, B.; Dreyfuss, G. Nup153 is an M9-containing mobile nucleoporin with a novel Ran-binding domain. EMBO J. 1999, 18, 1982–1995. [Google Scholar] [CrossRef]
- Favaloro, B.; Allocati, N.; Graziano, V.; Di Ilio, C.; De Laurenzi, V. Role of Apoptosis in Disease. Aging 2012, 4, 330–349. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/β-Catenin Signaling and Disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Scott, E.L.; Brann, D.W. Estrogen Regulation of Dkk1 and Wnt/Β-Catenin Signaling in Neurodegenerative Disease. Brain Res. 2013, 1514, 63–74. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Primer Sequences (5’–3’) |
|---|---|
| sox9a-F | CAG AGC GCA GTA CGA CTA TT |
| sox9a-R | GTA GCT GAA GGT GGA GTA CAG |
| col2a1a-F | CTG GTG ATC GTG GTG AGA TTG |
| col2a1a-R | TCA CCC TGC TCT CCC TTA TT |
| sox9b-F | GGC GCT CCT GCT AAC AAT AA |
| sox9b-R | ACC CTA ACC CTA ACC CTA ACC |
| caspase 8-F | GGA GAG AGA AAG GAG GAG AAA C |
| caspase 8-R | CCG CTG GGT CAG TAT GTA AT |
| bbc3-F | GGT TTC AAG CAC TTC CCT TAG A |
| bbc3-R | CCG ACG CAA ACA CAG AAA TG |
| fadd-F | AGT CGG TCA GAC AGT TCT TAT TG |
| fadd-R | GTG TTG ATT CTC TCT CGC TCT T |
| fsta-F | TGT GCC AAA CAG CAC ATT ATT C |
| fsta-R | CAG GAC CAC AGT CCA CAT TAT C |
| gadd45al -F | GAC GGA AGC TCC TTC AGA ATA C |
| gadd45al -R | GTC CTC AGA AAG TCC CAC AAA |
| mdm2-F | AAC AGC AAC TCG GAT GTA GG |
| mdm2-R | CCA CCT CAA ACT CCA CAC TAA |
| caspase 9-F | GGA GGA GGT GAG AAG GAT ATT G |
| caspase 9-R | CTG CTA GAG GAC ATG GGA ATA G |
| tp53-F | GTA CAA GTC CCT CCT GGA AAT C |
| tp53-R | GGC AAA TGC GTG TAA ACA GTA A |
| tp53bp1-F | CTA ACC CTG TCG CAT CCT TAT G |
| tp53bp1-R | AGG CTG AGA GTC CTC AAC TAT |
| tp53i11a-F | GAC CTC AGT AGA TGG TGG AAT G |
| tp53i11a-R | GAG ATG GAC GCA GAA CTC AA |
| tp53i11b-F | AGG AGA GGA TGA TGA TGG AGA G |
| tp53i11b-R | GGC GGA GAA GAG AAT CCA TAA C |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Li, X.; Gu, Q.; Li, Q.; Cui, Z. Nucleoporin 62-Like Protein is Required for the Development of Pharyngeal Arches through Regulation of Wnt/β-Catenin Signaling and Apoptotic Homeostasis in Zebrafish. Cells 2019, 8, 1038. https://doi.org/10.3390/cells8091038
Yang X, Li X, Gu Q, Li Q, Cui Z. Nucleoporin 62-Like Protein is Required for the Development of Pharyngeal Arches through Regulation of Wnt/β-Catenin Signaling and Apoptotic Homeostasis in Zebrafish. Cells. 2019; 8(9):1038. https://doi.org/10.3390/cells8091038
Chicago/Turabian StyleYang, Xiaojie, Xixi Li, Qilin Gu, Qing Li, and Zongbin Cui. 2019. "Nucleoporin 62-Like Protein is Required for the Development of Pharyngeal Arches through Regulation of Wnt/β-Catenin Signaling and Apoptotic Homeostasis in Zebrafish" Cells 8, no. 9: 1038. https://doi.org/10.3390/cells8091038
APA StyleYang, X., Li, X., Gu, Q., Li, Q., & Cui, Z. (2019). Nucleoporin 62-Like Protein is Required for the Development of Pharyngeal Arches through Regulation of Wnt/β-Catenin Signaling and Apoptotic Homeostasis in Zebrafish. Cells, 8(9), 1038. https://doi.org/10.3390/cells8091038