Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma

 ,
, 

Abstract
1. Introduction
2. Biosynthesis, Metabolism, Actions, and Signaling Pathways of Melatonin
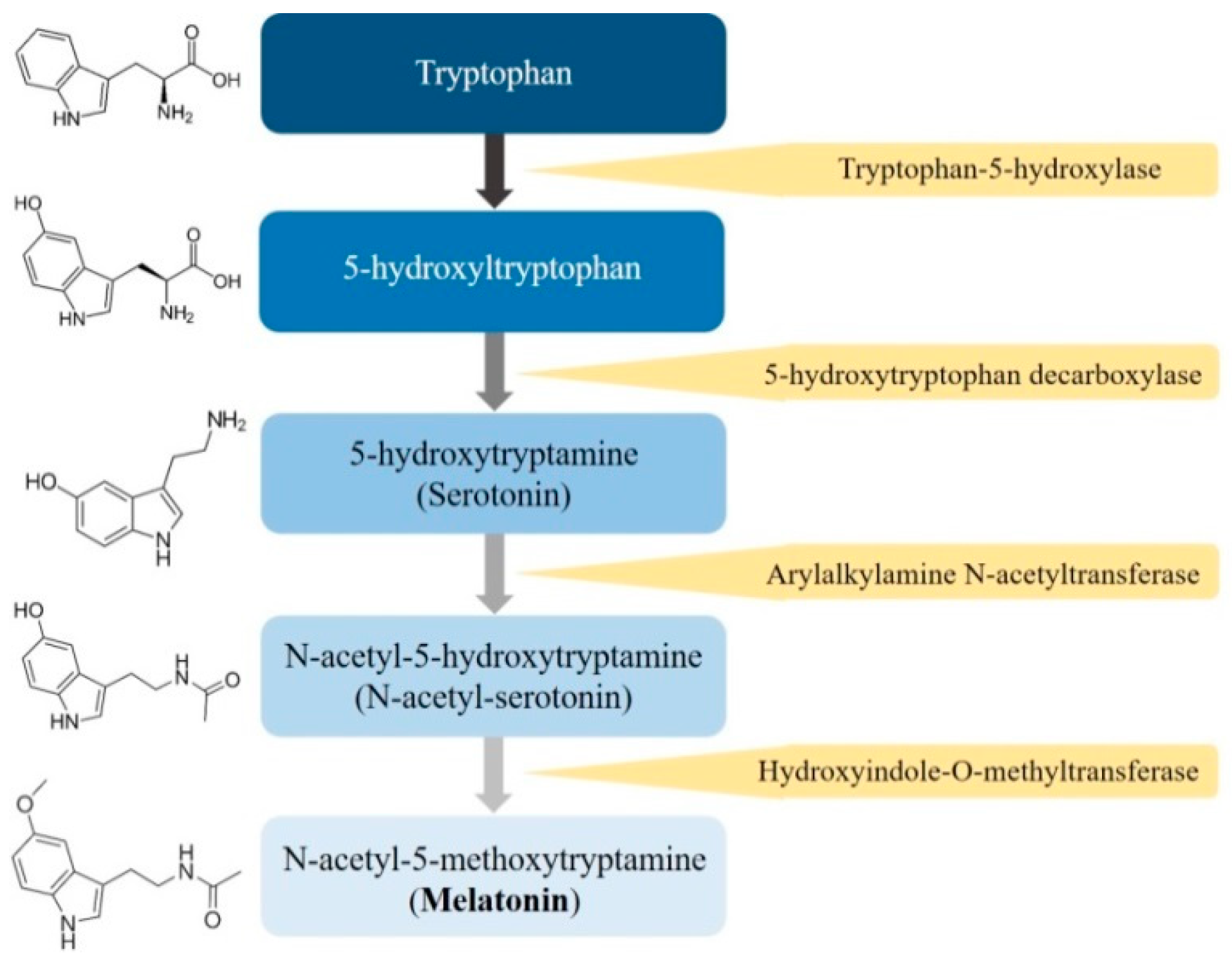
2.1. Biosynthesis of Melatonin
2.2. Metabolism of Melatonin
2.3. Actions of Melatonin
2.4. Signaling Pathways of Melatonin
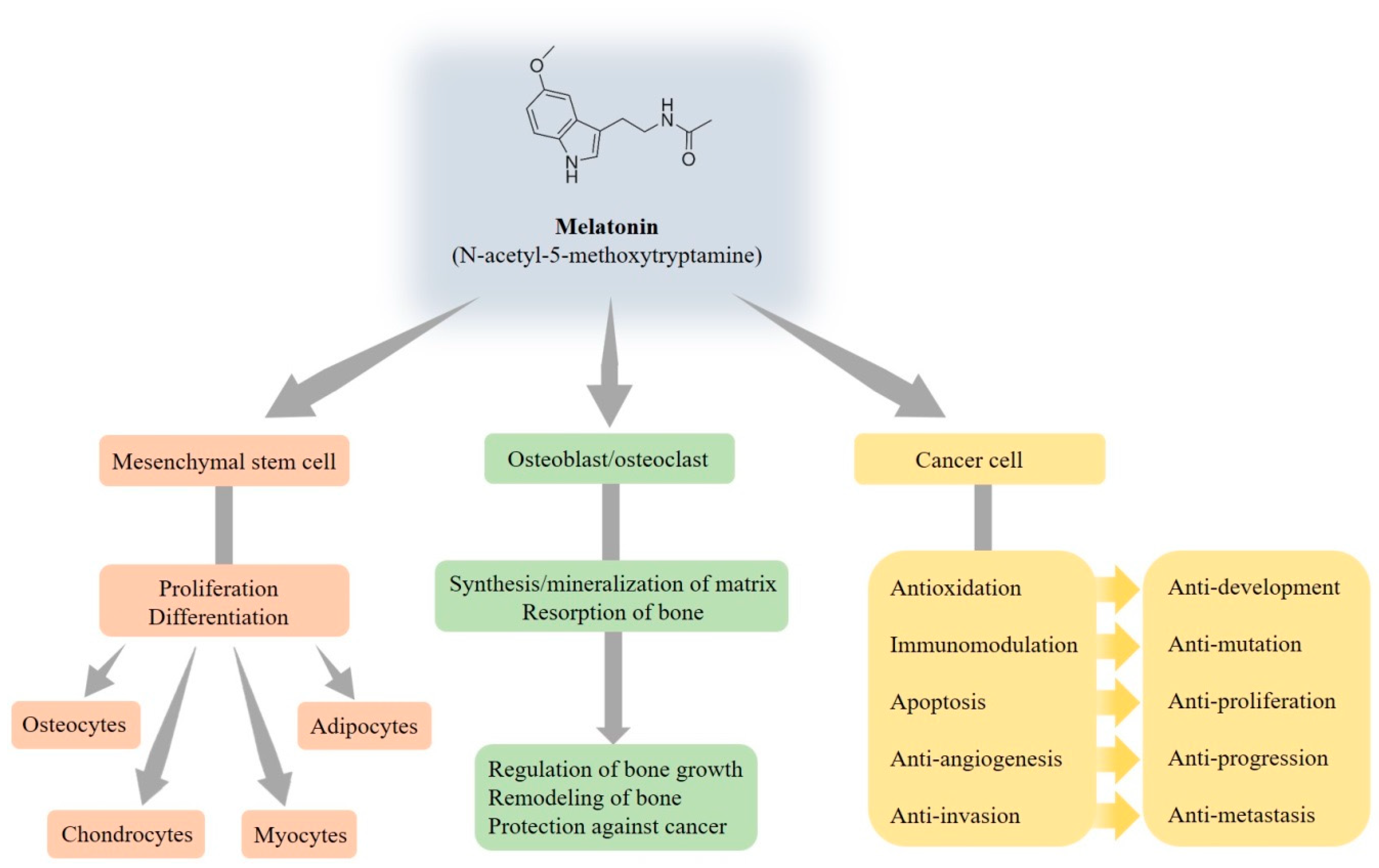
3. Function of Melatonin Relating to the Bone
3.1. Effects of Melatonin on Mesenchymal Stem Cells
3.2. Effects of Melatonin on Osteoblasts and Osteoclasts
3.3. Effects of Melatonin on Cancer Cells
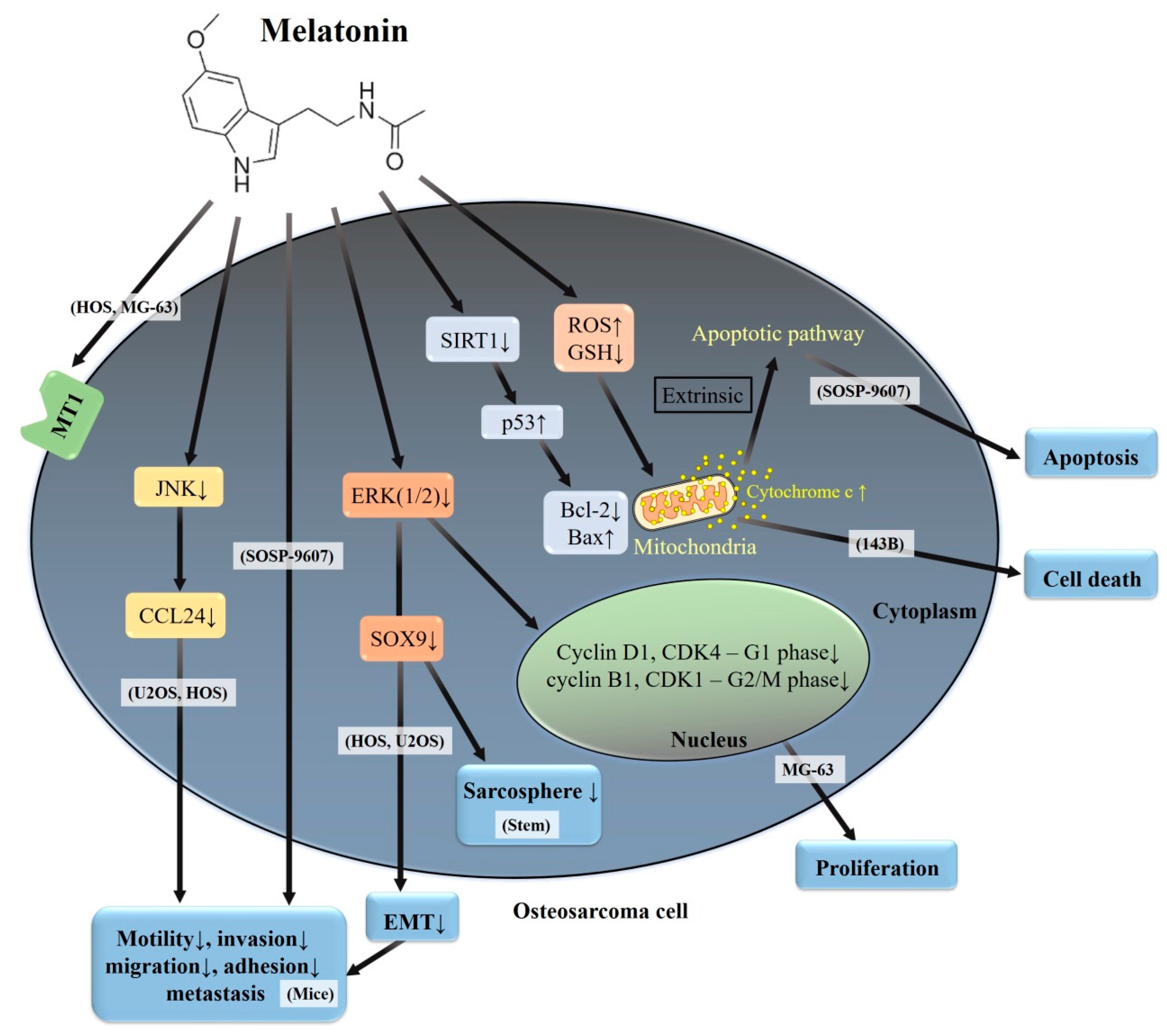
4. Molecular Actions of Melatonin in Osteosarcoma
4.1. Cytotoxic Activity of Melatonin in Human Osteosarcoma
4.2. Anti-Metastatic Effects of Melatonin in Human Osteosarcoma
4.3. Synergistic Effects and Utilization of Micro/Nanoparticles Delivery and Inclusion Complex Systems in Human Osteosarcoma
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BAX | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma/leukemia 2 |
| CCL24 | C-C motif chemokine ligand 24 |
| EMT | epithelial–mesenchymal transition |
| ERK | extracellular signal-regulated protein kinase |
| GSH | glutathione |
| JNK | c-Jun N-terminal kinase |
| MT1 | melatonin receptor 1 |
| ROS | reactive oxygen species |
| SIRT1 | sirtuin 1 |
References
- Li, T.; Jiang, S.; Yang, Y. Database selection and heterogeneity-more details, more credibility. JAMA Oncol. 2018, 4, 1295. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organization (WHO). Cancer. Available online: https://www.who.int/cancer/en/ (accessed on 10 December 2019).
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma incidence and survival rates from 1973 to 2004: Data from the surveillance, epidemiology, and end results program. Cancer 2009, 115, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Picci, P.; Mercuri, M.; Ferrari, S.; Alberghini, M.; Briccoli, A.; Ferrari, C.; Pignotti, E.; Bacci, G. Survival in high-grade osteosarcoma: Improvement over 21 years at a single institution. Ann. Oncol. 2010, 21, 1366–1373. [Google Scholar] [CrossRef] [PubMed]
- Mertens, W.C.; Bramwell, V. Osteosarcoma and other tumors of bone. Curr. Opin. Oncol. 1994, 6, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Briccoli, A.; Mercuri, M.; Bertoni, F.; Picci, P.; Tienghi, A.; Del Prever, A.B.; Fagioli, F.; Comandone, A.; Bacci, G. Postrelapse survival in osteosarcoma of the extremities: Prognostic factors for long-term survival. J. Clin. Oncol. 2003, 21, 710–715. [Google Scholar] [CrossRef]
- Ciernik, I.F.; Niemierko, A.; Harmon, D.C.; Kobayashi, W.; Chen, Y.L.; Yock, T.I.; Ebb, D.H.; Choy, E.; Raskin, K.A.; Liebsch, N.; et al. Proton-based radiotherapy for unresectable or incompletely resected osteosarcoma. Cancer 2011, 117, 4522–4530. [Google Scholar] [CrossRef]
- Oertel, S.; Blattmann, C.; Rieken, S.; Jensen, A.; Combs, S.E.; Huber, P.E.; Bischof, M.; Kulozik, A.; Debus, J.; Schulz-Ertner, D. Radiotherapy in the treatment of primary osteosarcoma--a single center experience. Tumori 2010, 96, 582–588. [Google Scholar] [CrossRef]
- Karasek, M.; Winczyk, K. Melatonin in humans. J. Physiol. Pharm. 2006, 57, 19–39. [Google Scholar]
- Waldhauser, F.; Weiszenbacher, G.; Frisch, H.; Zeitlhuber, U.; Waldhauser, M.; Wurtman, R.J. Fall in nocturnal serum melatonin during prepuberty and pubescence. Lancet 1984, 1, 362–365. [Google Scholar] [CrossRef]
- Panzer, A. Melatonin in osteosarcoma: An effective drug? Med. Hypotheses 1997, 48, 523–525. [Google Scholar] [CrossRef]
- Sanchez-Barcelo, E.J.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Reiter, R.J. Melatonin uses in oncology: Breast cancer prevention and reduction of the side effects of chemotherapy and radiation. Expert. Opin. Investig. Drugs 2012, 21, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Acuna-Castroviejo, D.; Escames, G.; Venegas, C.; Diaz-Casado, M.E.; Lima-Cabello, E.; Lopez, L.C.; Rosales-Corral, S.; Tan, D.X.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]
- Remy, P.; Doder, M.; Lees, A.; Turjanski, N.; Brooks, D. Depression in parkinson’s disease: Loss of dopamine and noradrenaline innervation in the limbic system. Brain 2005, 128, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Schomerus, C.; Korf, H.W. Mechanisms regulating melatonin synthesis in the mammalian pineal organ. Ann. N. Y. Acad. Sci. 2005, 1057, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. Pineal melatonin: Cell biology of its synthesis and of its physiological interactions. Endocr. Rev. 1991, 12, 151–180. [Google Scholar] [CrossRef]
- Li, T.; Jiang, S.; Lu, C.; Yang, W.; Yang, Z.; Hu, W.; Xin, Z.; Yang, Y. Melatonin: Another avenue for treating osteoporosis? J. Pineal Res. 2019, 66, e12548. [Google Scholar] [CrossRef]
- Brzezinski, A. Melatonin in humans. N. Engl. J. Med. 1997, 336, 186–195. [Google Scholar] [CrossRef]
- Di, W.L.; Kadva, A.; Johnston, A.; Silman, R. Variable bioavailability of oral melatonin. N. Engl. J. Med. 1997, 336, 1028–1029. [Google Scholar] [CrossRef]
- Vijayalaxmi; Thomas, C.R., Jr.; Reiter, R.J.; Herman, T.S. Melatonin: From basic research to cancer treatment clinics. J. Clin. Oncol. 2002, 20, 2575–2601. [Google Scholar] [CrossRef]
- Tan, D.; Manchester, L.C.; Reiter, R.J.; Qi, W.; Hanes, M.A.; Farley, N.J. High physiological levels of melatonin in the bile of mammals. Life Sci. 1999, 65, 2523–2529. [Google Scholar] [CrossRef]
- Wetterberg, L. Melatonin and clinical application. Reprod. Nutr. Dev. 1999, 39, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Lynch, H.J.; Wurtman, R.J.; Moskowitz, M.A.; Archer, M.C.; Ho, M.H. Daily rhythm in human urinary melatonin. Science 1975, 187, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Stehle, J.H.; von Gall, C.; Korf, H.W. Melatonin: A clock-output, a clock-input. J. Neuroendocr. 2003, 15, 383–389. [Google Scholar] [CrossRef]
- Luchetti, F.; Canonico, B.; Betti, M.; Arcangeletti, M.; Pilolli, F.; Piroddi, M.; Canesi, L.; Papa, S.; Galli, F. Melatonin signaling and cell protection function. FASEB J. 2010, 24, 3603–3624. [Google Scholar] [CrossRef]
- Stevens, R.G.; Brainard, G.C.; Blask, D.E.; Lockley, S.W.; Motta, M.E. Breast cancer and circadian disruption from electric lighting in the modern world. CA Cancer J. Clin. 2014, 64, 207–218. [Google Scholar] [CrossRef]
- Carlberg, C. Gene regulation by melatonin. Ann. N. Y. Acad. Sci. 2000, 917, 387–396. [Google Scholar] [CrossRef]
- Reiter, R.J. Oxidative damage in the central nervous system: Protection by melatonin. Prog. Neurobiol. 1998, 56, 359–384. [Google Scholar] [CrossRef]
- Srinivasan, V.; Spence, D.W.; Pandi-Perumal, S.R.; Trakht, I.; Cardinali, D.P. Therapeutic actions of melatonin in cancer: Possible mechanisms. Integr. Cancer 2008, 7, 189–203. [Google Scholar] [CrossRef]
- Deming, S.L.; Lu, W.; Beeghly-Fadiel, A.; Zheng, Y.; Cai, Q.; Long, J.; Shu, X.O.; Gao, Y.T.; Zheng, W. Melatonin pathway genes and breast cancer risk among chinese women. Breast Cancer Res. Treat. 2012, 132, 693–699. [Google Scholar] [CrossRef]
- Maria, S.; Samsonraj, R.M.; Munmun, F.; Glas, J.; Silvestros, M.; Kotlarczyk, M.P.; Rylands, R.; Dudakovic, A.; van Wijnen, A.J.; Enderby, L.T.; et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: Implications of a role for mt2 melatonin receptors, mek1/2, and mek5 in melatonin-mediated osteoblastogenesis. J. Pineal Res. 2018, 64. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.M.; Belancio, V.P.; Dauchy, R.T.; Xiang, S.; Brimer, S.; Mao, L.; Hauch, A.; Lundberg, P.W.; Summers, W.; Yuan, L.; et al. Melatonin: An inhibitor of breast cancer. Endocr. Relat. Cancer 2015, 22, R183–R204. [Google Scholar] [CrossRef] [PubMed]
- Levi, B.; Longaker, M.T. Concise review: Adipose-derived stromal cells for skeletal regenerative medicine. Stem Cells 2011, 29, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F.; Canonico, B.; Bartolini, D.; Arcangeletti, M.; Ciffolilli, S.; Murdolo, G.; Piroddi, M.; Papa, S.; Reiter, R.J.; Galli, F. Melatonin regulates mesenchymal stem cell differentiation: A review. J. Pineal Res. 2014, 56, 382–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Su, P.; Xu, C.; Chen, C.; Liang, A.; Du, K.; Peng, Y.; Huang, D. Melatonin inhibits adipogenesis and enhances osteogenesis of human mesenchymal stem cells by suppressing ppargamma expression and enhancing runx2 expression. J. Pineal Res. 2010, 49, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Radio, N.M.; Doctor, J.S.; Witt-Enderby, P.A. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via mt2 melatonin receptors and the mek/erk (1/2) signaling cascade. J. Pineal Res. 2006, 40, 332–342. [Google Scholar] [CrossRef]
- Arendt, J. Melatonin and human rhythms. Chronobiol. Int. 2006, 23, 21–37. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Ladizesky, M.G.; Boggio, V.; Cutrera, R.A.; Mautalen, C. Melatonin effects on bone: Experimental facts and clinical perspectives. J. Pineal Res. 2003, 34, 81–87. [Google Scholar] [CrossRef]
- Park, K.H.; Kang, J.W.; Lee, E.M.; Kim, J.S.; Rhee, Y.H.; Kim, M.; Jeong, S.J.; Park, Y.G.; Kim, S.H. Melatonin promotes osteoblastic differentiation through the bmp/erk/wnt signaling pathways. J. Pineal Res. 2011, 51, 187–194. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Fuentes-Broto, L.; Paredes, S.D.; Reiter, R.J. Significance and application of melatonin in the regulation of brown adipose tissue metabolism: Relation to human obesity. Obes. Rev. 2011, 12, 167–188. [Google Scholar] [CrossRef]
- Nakade, O.; Koyama, H.; Ariji, H.; Yajima, A.; Kaku, T. Melatonin stimulates proliferation and type i collagen synthesis in human bone cells in vitro. J. Pineal Res. 1999, 27, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D. How important is the pineal gland in children. In Adv. Pineal Research; Reiter, R.J., Pang, S.F., Eds.; John Libbey: London, UK, 1989; pp. 291–297. [Google Scholar]
- Ho, H.Y.; Lin, C.W.; Chien, M.H.; Reiter, R.J.; Su, S.C.; Hsieh, Y.H.; Yang, S.F. Melatonin suppresses tpa-induced metastasis by downregulating matrix metalloproteinase-9 expression through jnk/sp-1 signaling in nasopharyngeal carcinoma. J. Pineal Res. 2016, 61, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.H.; Su, S.C.; Lin, C.W.; Hsieh, Y.H.; Lin, Y.C.; Chien, M.H.; Reiter, R.J.; Yang, S.F. Melatonin attenuates osteosarcoma cell invasion by suppression of c-c motif chemokine ligand 24 through inhibition of the c-jun n-terminal kinase pathway. J. Pineal Res. 2018, 65, e12507. [Google Scholar] [CrossRef] [PubMed]
- Hoang, B.X.; Shaw, D.G.; Pham, P.T.; Levine, S.A. Neurobiological effects of melatonin as related to cancer. Eur. J. Cancer Prev. 2007, 16, 511–516. [Google Scholar] [CrossRef]
- Su, S.C.; Hsieh, M.J.; Yang, W.E.; Chung, W.H.; Reiter, R.J.; Yang, S.F. Cancer metastasis: Mechanisms of inhibition by melatonin. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.A.; Tan, D.X.; Acuna-Castroviejo, D.; Qin, L.; Yang, S.F.; Xu, K. Melatonin, a full service anti-cancer agent: Inhibition of initiation, progression and metastasis. Int. J. Mol. Sci. 2017, 18, 843. [Google Scholar] [CrossRef]
- Yu, Q.; Miller, S.C.; Osmond, D.G. Melatonin inhibits apoptosis during early b-cell development in mouse bone marrow. J. Pineal Res. 2000, 29, 86–93. [Google Scholar] [CrossRef]
- Bizzarri, M.; Proietti, S.; Cucina, A.; Reiter, R.J. Molecular mechanisms of the pro-apoptotic actions of melatonin in cancer: A review. Exp. Opin. Targets 2013, 17, 1483–1496. [Google Scholar] [CrossRef]
- Hardeland, R. Antioxidative protection by melatonin: Multiplicity of mechanisms from radical detoxification to radical avoidance. Endocrine 2005, 27, 119–130. [Google Scholar] [CrossRef]
- Maestroni, G.J. The immunoneuroendocrine role of melatonin. J. Pineal Res. 1993, 14, 1–10. [Google Scholar] [CrossRef]
- Garcia-Maurino, S.; Gonzalez-Haba, M.G.; Calvo, J.R.; Rafii-El-Idrissi, M.; Sanchez-Margalet, V.; Goberna, R.; Guerrero, J.M. Melatonin enhances il-2, il-6, and ifn-gamma production by human circulating cd4+ cells: A possible nuclear receptor-mediated mechanism involving t helper type 1 lymphocytes and monocytes. J. Immunol. 1997, 159, 574–581. [Google Scholar] [PubMed]
- Blask, D.E.; Brainard, G.C.; Dauchy, R.T.; Hanifin, J.P.; Davidson, L.K.; Krause, J.A.; Sauer, L.A.; Rivera-Bermudez, M.A.; Dubocovich, M.L.; Jasser, S.A.; et al. Melatonin-depleted blood from premenopausal women exposed to light at night stimulates growth of human breast cancer xenografts in nude rats. Cancer Res. 2005, 65, 11174–11184. [Google Scholar] [CrossRef] [PubMed]
- Leon-Blanco, M.M.; Guerrero, J.M.; Reiter, R.J.; Calvo, J.R.; Pozo, D. Melatonin inhibits telomerase activity in the mcf-7 tumor cell line both in vivo and in vitro. J. Pineal Res. 2003, 35, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Leon, J.; Casado, J.; Jimenez Ruiz, S.M.; Zurita, M.S.; Gonzalez-Puga, C.; Rejon, J.D.; Gila, A.; Munoz de Rueda, P.; Pavon, E.J.; Reiter, R.J.; et al. Melatonin reduces endothelin-1 expression and secretion in colon cancer cells through the inactivation of foxo-1 and nf-kappabeta. J. Pineal Res. 2014, 56, 415–426. [Google Scholar] [CrossRef]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Lu, K.H.; Chen, P.N.; Lue, K.H.; Lai, M.T.; Lin, M.S.; Hsieh, Y.S.; Chu, S.C. 2’-hydroxyflavanone induces apoptosis of human osteosarcoma 143 b cells by activating the extrinsic trail- and intrinsic mitochondria-mediated pathways. Nutr. Cancer 2014, 66, 625–635. [Google Scholar] [CrossRef]
- Degterev, A.; Boyce, M.; Yuan, J. A decade of caspases. Oncogene 2003, 22, 8543–8567. [Google Scholar] [CrossRef]
- Davis, R.J. Signal transduction by the jnk group of map kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.W. Nf-kappab in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef]
- Toma, C.D.; Svoboda, M.; Arrich, F.; Ekmekcioglu, C.; Assadian, O.; Thalhammer, T. Expression of the melatonin receptor (mt) 1 in benign and malignant human bone tumors. J. Pineal Res. 2007, 43, 206–213. [Google Scholar] [CrossRef]
- Peng, T.I.; Hsiao, C.W.; Reiter, R.J.; Tanaka, M.; Lai, Y.K.; Jou, M.J. Mtdna t8993g mutation-induced mitochondrial complex v inhibition augments cardiolipin-dependent alterations in mitochondrial dynamics during oxidative, ca(2+), and lipid insults in narp cybrids: A potential therapeutic target for melatonin. J. Pineal Res. 2012, 52, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Panzer, A.; Lottering, M.L.; Bianchi, P.; Glencross, D.K.; Stark, J.H.; Seegers, J.C. Melatonin has no effect on the growth, morphology or cell cycle of human breast cancer (mcf-7), cervical cancer (hela), osteosarcoma (mg-63) or lymphoblastoid (tk6) cells. Cancer Lett. 1998, 122, 17–23. [Google Scholar] [CrossRef]
- Liu, L.; Xu, Y.; Reiter, R.J. Melatonin inhibits the proliferation of human osteosarcoma cell line mg-63. Bone 2013, 55, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xu, Y.; Reiter, R.J.; Pan, Y.; Chen, D.; Liu, Y.; Pu, X.; Jiang, L.; Li, Z. Inhibition of erk1/2 signaling pathway is involved in melatonin’s antiproliferative effect on human mg-63 osteosarcoma cells. Cell Physiol. Biochem. 2016, 39, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Cai, L.; Jiang, P.; Wang, J.; Gao, C.; Feng, H.; Wang, C.; Pan, H.; Yang, Y. Sirt1 inhibition by melatonin exerts antitumor activity in human osteosarcoma cells. Eur. J. Pharm. 2013, 715, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Jung-Hynes, B.; Reiter, R.J.; Ahmad, N. Sirtuins, melatonin and circadian rhythms: Building a bridge between aging and cancer. J. Pineal Res. 2010, 48, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Valastyan, S.; Weinberg, R.A. Tumor metastasis: Molecular insights and evolving paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef]
- Hsieh, Y.S.; Chu, S.C.; Yang, S.F.; Chen, P.N.; Liu, Y.C.; Lu, K.H. Silibinin suppresses human osteosarcoma mg-63 cell invasion by inhibiting the erk-dependent c-jun/ap-1 induction of mmp-2. Carcinogenesis 2007, 28, 977–987. [Google Scholar] [CrossRef]
- Cheng, H.L.; Lin, C.W.; Yang, J.S.; Hsieh, M.J.; Yang, S.F.; Lu, K.H. Zoledronate blocks geranylgeranylation not farnesylation to suppress human osteosarcoma u2os cells metastasis by emt via rho a activation and fak-inhibited jnk and p38 pathways. Oncotarget 2016, 7, 9742–9758. [Google Scholar] [CrossRef]
- Kahlert, U.D.; Joseph, J.V.; Kruyt, F.A.E. Emt- and met-related processes in nonepithelial tumors: Importance for disease progression, prognosis, and therapeutic opportunities. Mol. Oncol. 2017, 11, 860–877. [Google Scholar] [CrossRef]
- Kim, E.H.; Kim, M.S.; Lee, K.H.; Koh, J.S.; Jung, W.G.; Kong, C.B. Zoledronic acid is an effective radiosensitizer in the treatment of osteosarcoma. Oncotarget 2016, 7, 70869–70880. [Google Scholar] [CrossRef] [PubMed]
- Daw, N.C.; Chou, A.J.; Jaffe, N.; Rao, B.N.; Billups, C.A.; Rodriguez-Galindo, C.; Meyers, P.A.; Huh, W.W. Recurrent osteosarcoma with a single pulmonary metastasis: A multi-institutional review. Br. J. Cancer 2015, 112, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Jung-Hynes, B.; Ahmad, N. Sirt1 controls circadian clock circuitry and promotes cell survival: A connection with age-related neoplasms. FASEB J. 2009, 23, 2803–2809. [Google Scholar] [CrossRef] [PubMed]
- Jung-Hynes, B.; Ahmad, N. Role of p53 in the anti-proliferative effects of sirt1 inhibition in prostate cancer cells. Cell Cycle 2009, 8, 1478–1483. [Google Scholar] [CrossRef]
- Qu, H.; Xue, Y.; Lian, W.; Wang, C.; He, J.; Fu, Q.; Zhong, L.; Lin, N.; Lai, L.; Ye, Z.; et al. Melatonin inhibits osteosarcoma stem cells by suppressing sox9-mediated signaling. Life Sci. 2018, 207, 253–264. [Google Scholar] [CrossRef]
- Fan, L.L.; Sun, G.P.; Wei, W.; Wang, Z.G.; Ge, L.; Fu, W.Z.; Wang, H. Melatonin and doxorubicin synergistically induce cell apoptosis in human hepatoma cell lines. World J. Gastroenterol. 2010, 16, 1473–1481. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Z.; Chua, C.C.; Ma, Y.S.; Youngberg, G.A.; Hamdy, R.; Chua, B.H. Melatonin as an effective protector against doxorubicin-induced cardiotoxicity. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H254–H263. [Google Scholar] [CrossRef]
- Wang, Y.P.; Yang, Z.P. Effects of melatonin combined with cis-platinum or methotrexate on the proliferation of osteosarcoma cell line saos-2. Acta Acad. Med. Sin. 2015, 37, 215–220. [Google Scholar]
- Nataraj, V.; Rastogi, S.; Khan, S.A.; Sharma, M.C.; Agarwala, S.; Vishnubhatla, S.; Bakhshi, S. Prognosticating metastatic osteosarcoma treated with uniform chemotherapy protocol without high dose methotrexate and delayed metastasectomy: A single center experience of 102 patients. Clin. Transl. Oncol. 2016, 18, 937–944. [Google Scholar] [CrossRef]
- Liu, L.; Zhu, Y.; Xu, Y.; Reiter, R.J. Melatonin delays cell proliferation by inducing g1 and g2/m phase arrest in a human osteoblastic cell line hfob 1.19. J. Pineal Res. 2011, 50, 222–231. [Google Scholar] [CrossRef]
- Altindal, D.C.; Gumusderelioglu, M. Melatonin releasing plga micro/nanoparticles and their effect on osteosarcoma cells. J. Microencapsul. 2016, 33, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, H.; Kato, K.I.; Ibayashi, H. Melatonin serum levels and metabolic clearance rate in patients with liver cirrhosis. J. Clin. Endocrinol. Metab. 1982, 54, 1025–1027. [Google Scholar] [CrossRef] [PubMed]
- Topal, B.; Cetin Altindal, D.; Gumusderelioglu, M. Melatonin/hpbetacd complex: Microwave synthesis, integration with chitosan scaffolds and inhibitory effects on mg-63cells. Int. J. Pharm. 2015, 496, 801–811. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Action | Pathway | Cell Line/In Vivo | Dose | Reference |
|---|---|---|---|---|
| No effect on the growth, morphology or cell cycle | MG-63 | 10−5–10−13 M | Panzer A, et al. 1998. [64] | |
| Abundant expression of MT1-mRNA | HOS, MG-63 | Toma CD, et al. 2007. [62] | ||
| Rescues cardiolipin-dependent mitochondrial dynamics-associated mitochondrial pathologies | Reduces mitochondrial ROS, cell death, and depletion of cardiolipin to improve retardation of mitochondrial movement and dynamics | 143B | 100 μM | Peng TI, et al. 2012. [63] |
| Inhibits proliferation | Down-regulates cyclin D1 and CDK4, (G1 phase), and cyclin B1 and CDK1 (G2/M phase) | MG-63 | 4(−10) mM | Liu L, et al. 2013. [65] |
| Induces apoptosis, increases ROS, and decreases GSH | Down-regulates SIRT1 and mitochondrial apoptotic pathway, and up-regulates acetylated-p53 | SOSP-9607 | (250−)1000 μM | Cheng Y, et al. 2013. [67] |
| Reduces adhesion and migration | 12.5–50 μM | |||
| Inhibits proliferation, induces G1 and G2/M phase arrest, and down-regulates cyclin D1, CDK4, cyclin B1 and CDK1 | Inhibits the ERK1/2 pathway | MG-63 | 4 mM | Liu L, et al. 2016. [66] |
| Action | Pathway | Cell Line/In Vivo | Dose | Reference |
|---|---|---|---|---|
| Induces apoptosis, increases ROS, and decreases GSH | Down-regulates SIRT1 and mitochondrial apoptotic pathway, and up-regulates acetylated-p53 | SOSP-9607 | (250−)1000 μM | Cheng Y, et al. 2013. [67] |
| Reduces adhesion and migration | 12.5–50 μM | |||
| Reduces the number (anti-proliferation) | Suppresses EMT via downregulation of SOX9, via the ERK pathway | Stem cell | >0.5 mM | Qu H, et al. 2018. [77] |
| Suppresses migration and invasion | U2OS, HOS | 0.5 mM | ||
| Inhibits initiation and metastasis in vivo | Mice model | 100 mg/kg | ||
| Inhibits motility, migration and invasiveness, and the CCL24 gene expression | Attenuates invasion and migration by suppression of CCL24 through inhibition of the JNK pathway | U2OS, HOS | 2 mM | Lu KH, et al. 2018. [45] |
| Recombinant human CCL24 enhances migration and silencing of CCL24 attenuates migration and invasion | ||||
| Suppresses CCL24 and anti-metastasis through inhibition of the JNK pathway |
| Action | Pathway | Cell Line/In Vivo | Dose | Reference |
|---|---|---|---|---|
| Inhibits cell activity, blocks the cell cycle at G1-stage, and induces apoptosis | Blocks the cell cycle at G1-stage | SaOS-2 | 0.5–5 mM | Wang YP, et al. 2015. [80] |
| An antagonistic effect with lower concentration of cis-platinum | 1 mM | |||
| A synergistic effect with methotrexate or higher concentration of cis-platinum | ||||
| Melatonin releasing PLGA micro/nanoparticles increases the inhibitory effect | Increases the inhibitory effect of proliferation | MG-63 | Altındal DÇ, et al. 2016. [83] | |
| Melatonin/HPβCD inclusion complex loaded into chitosan scaffolds causes cell death | Melatonin/HPβCD system reduces the proportion in the G2/M phase rather than S phase | MG-63 | 9 mM | Topal B, et al. 2015. [85] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, K.-H.; Lin, R.-C.; Yang, J.-S.; Yang, W.-E.; Reiter, R.J.; Yang, S.-F. Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma. Cells 2019, 8, 1618. https://doi.org/10.3390/cells8121618
Lu K-H, Lin R-C, Yang J-S, Yang W-E, Reiter RJ, Yang S-F. Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma. Cells. 2019; 8(12):1618. https://doi.org/10.3390/cells8121618
Chicago/Turabian StyleLu, Ko-Hsiu, Renn-Chia Lin, Jia-Sin Yang, Wei-En Yang, Russel J. Reiter, and Shun-Fa Yang. 2019. "Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma" Cells 8, no. 12: 1618. https://doi.org/10.3390/cells8121618
APA StyleLu, K.-H., Lin, R.-C., Yang, J.-S., Yang, W.-E., Reiter, R. J., & Yang, S.-F. (2019). Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma. Cells, 8(12), 1618. https://doi.org/10.3390/cells8121618