Cobalamin Deficiency: Effect on Homeostasis of Cultured Human Astrocytes

 , and
, and 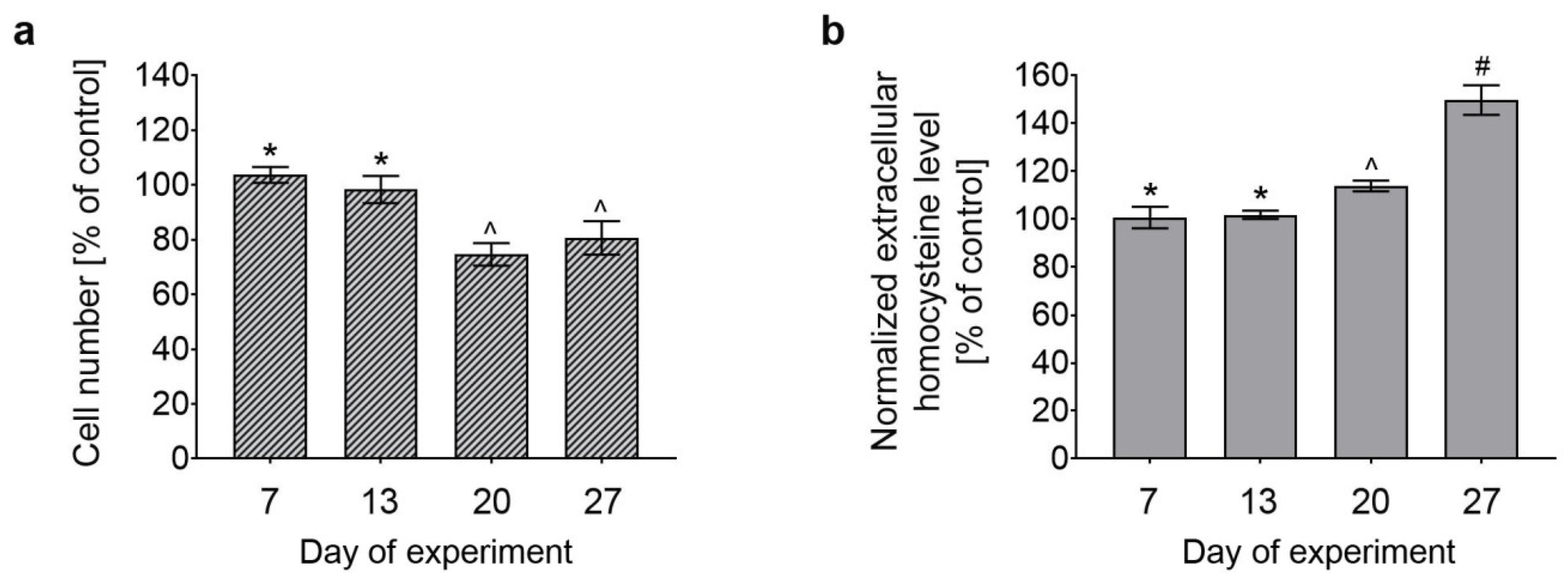{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. The Induction Of Cobalamin Deficiency
2.4. Cell Count Assay
2.5. Analysis of Extracellular Homocysteine Level
2.6. Cell Cycle Assay and DNA Fragmentation Assay
2.7. Mitochondrial Potential Assay
2.8. Annexin V Assay
2.9. Caspase Activity Assay
2.10. Assessment of Cell Morphology
2.11. Antioxidant Enzyme Activity Assays
2.12. Intracellular ROS Level Assay
2.13. Intracellular Thiol Status
2.14. Statistical Analysis
3. Results
3.1. Effect of Long-Term Treatment With (OH)Cbl(c-lactam) on Astrocyte Proliferation and Extracellular Hcy Level
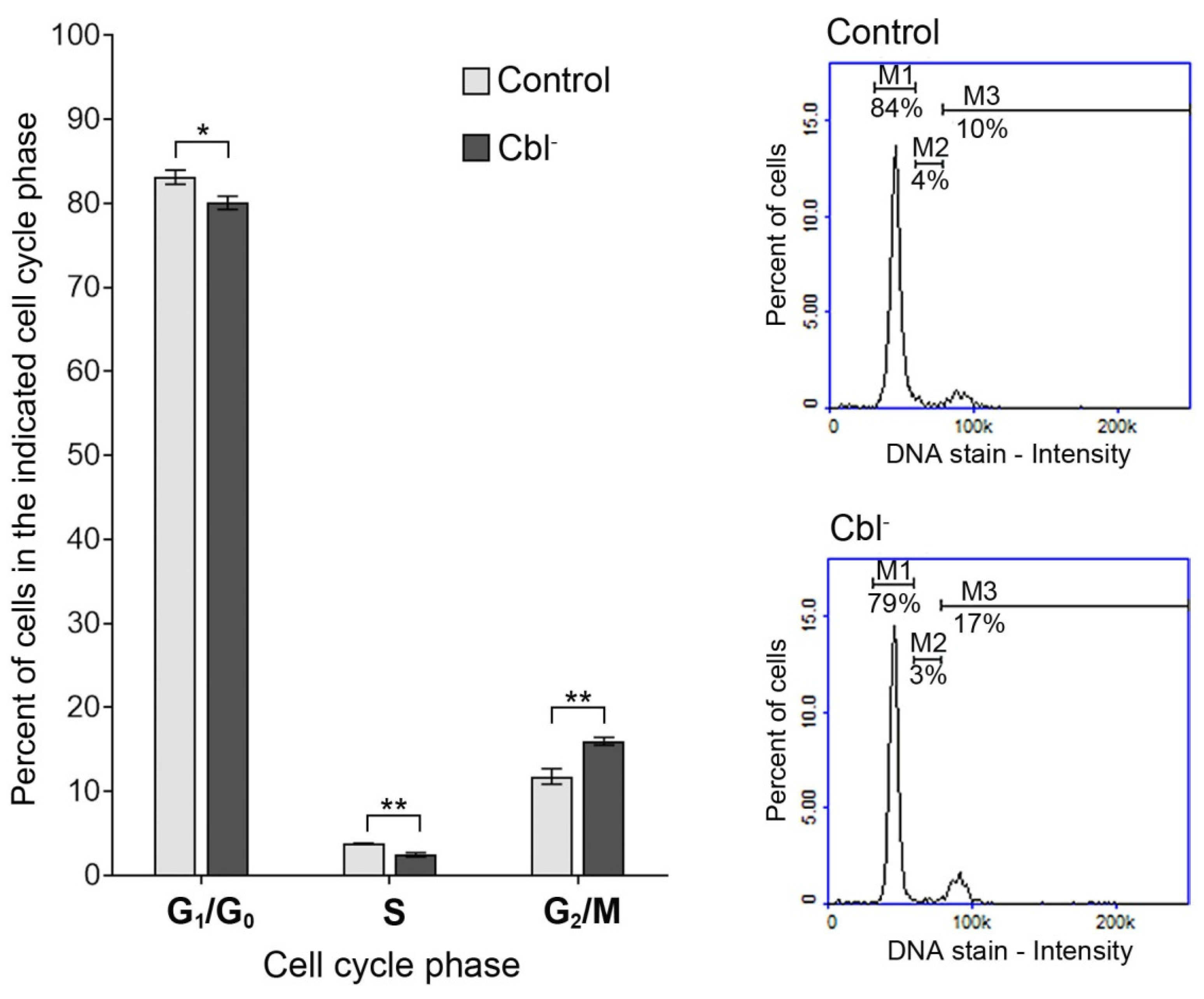
3.2. Cell Cycle of Astrocytes Under Conditions of Cobalamin Deficiency
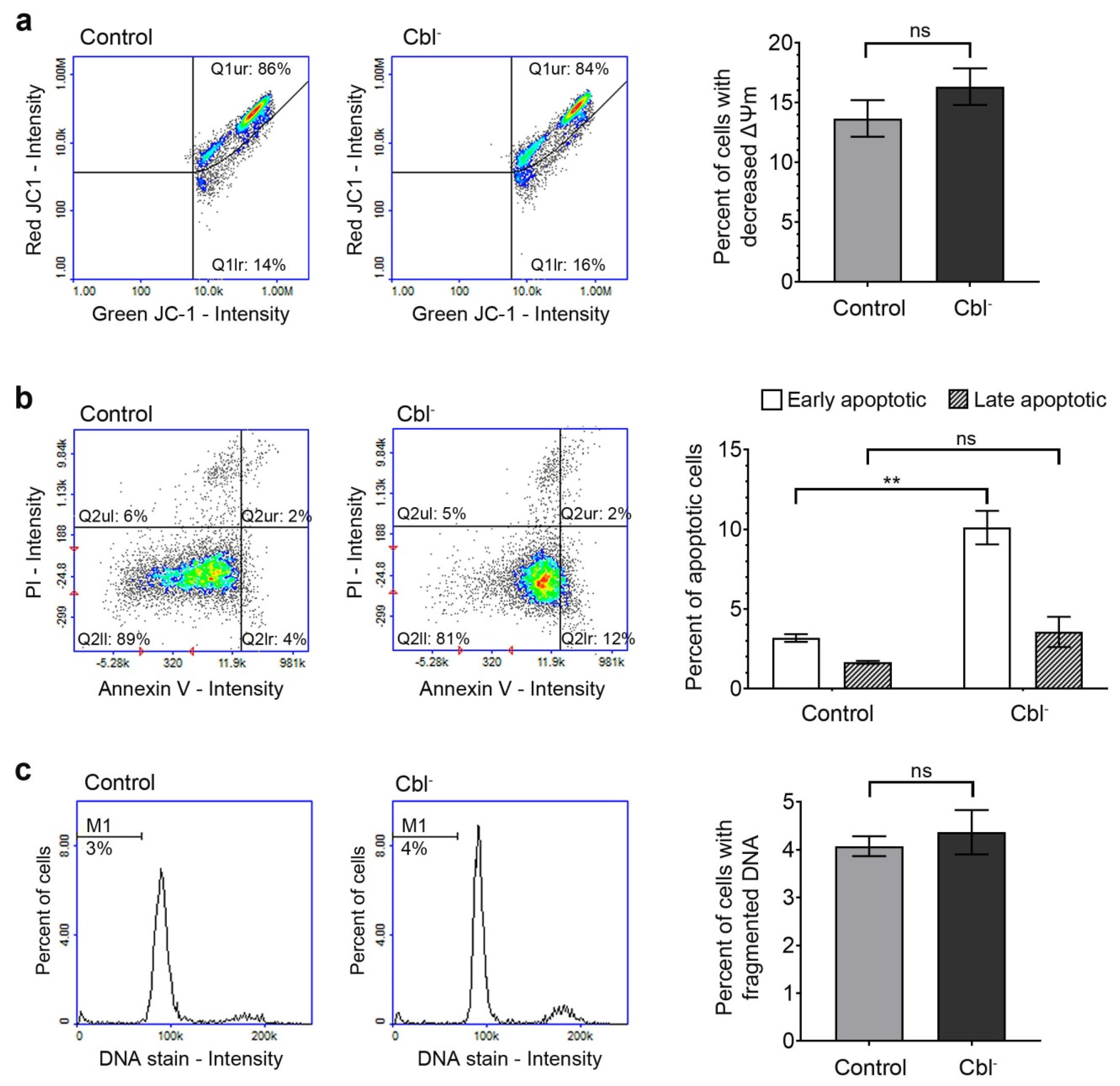
3.3. Evaluation of Apoptotic Markers in the Cobalamin-Deficient Astrocytes
3.3.1. Mitochondrial Membrane Depolarization
3.3.2. Phosphatidylserine Externalization
3.3.3. DNA Fragmentation
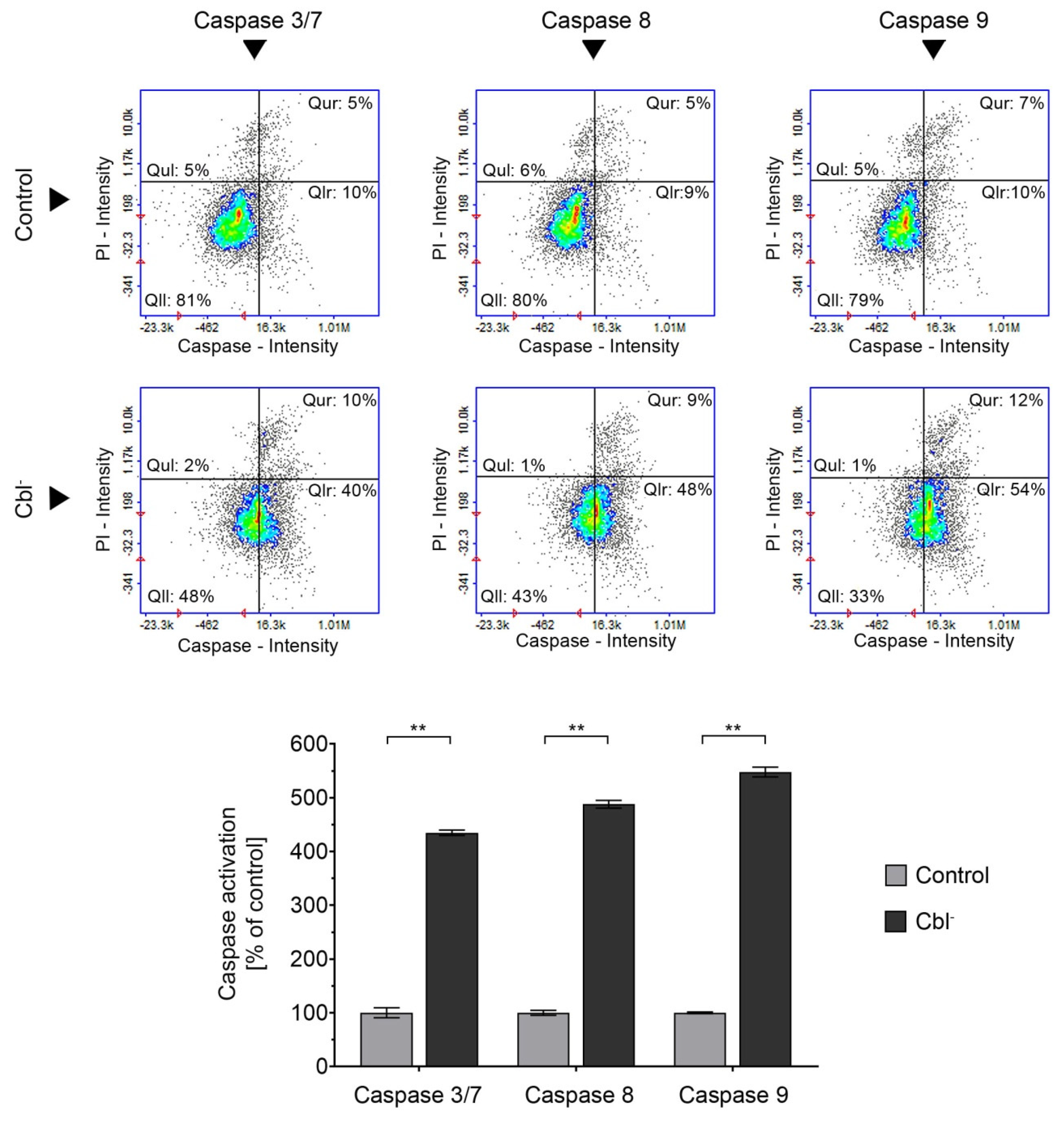
3.4. Caspase Activation in the Cobalamin-Deficient Astrocytes
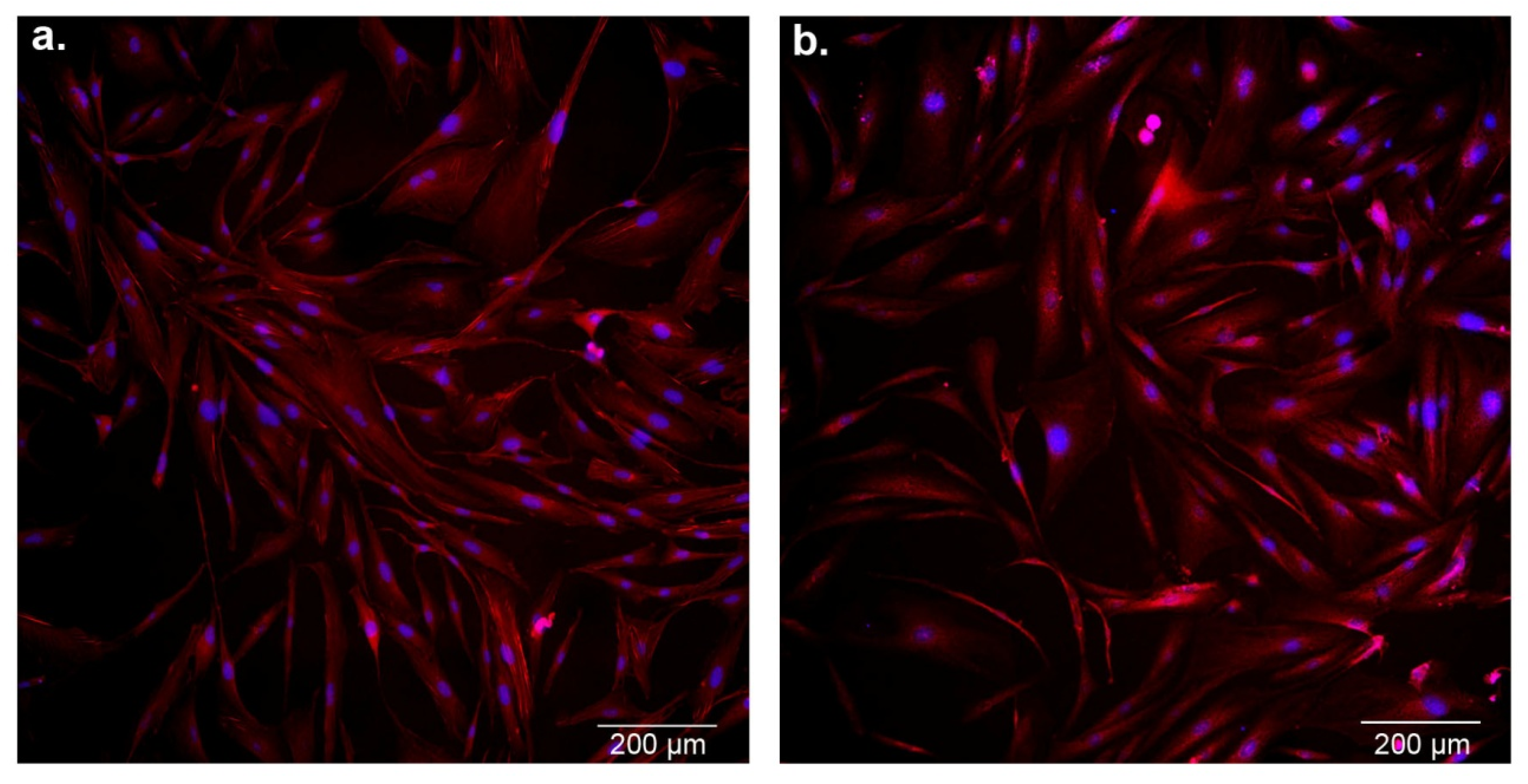
3.5. Effect of Cobalamin Deficiency on Astrocyte Morphology
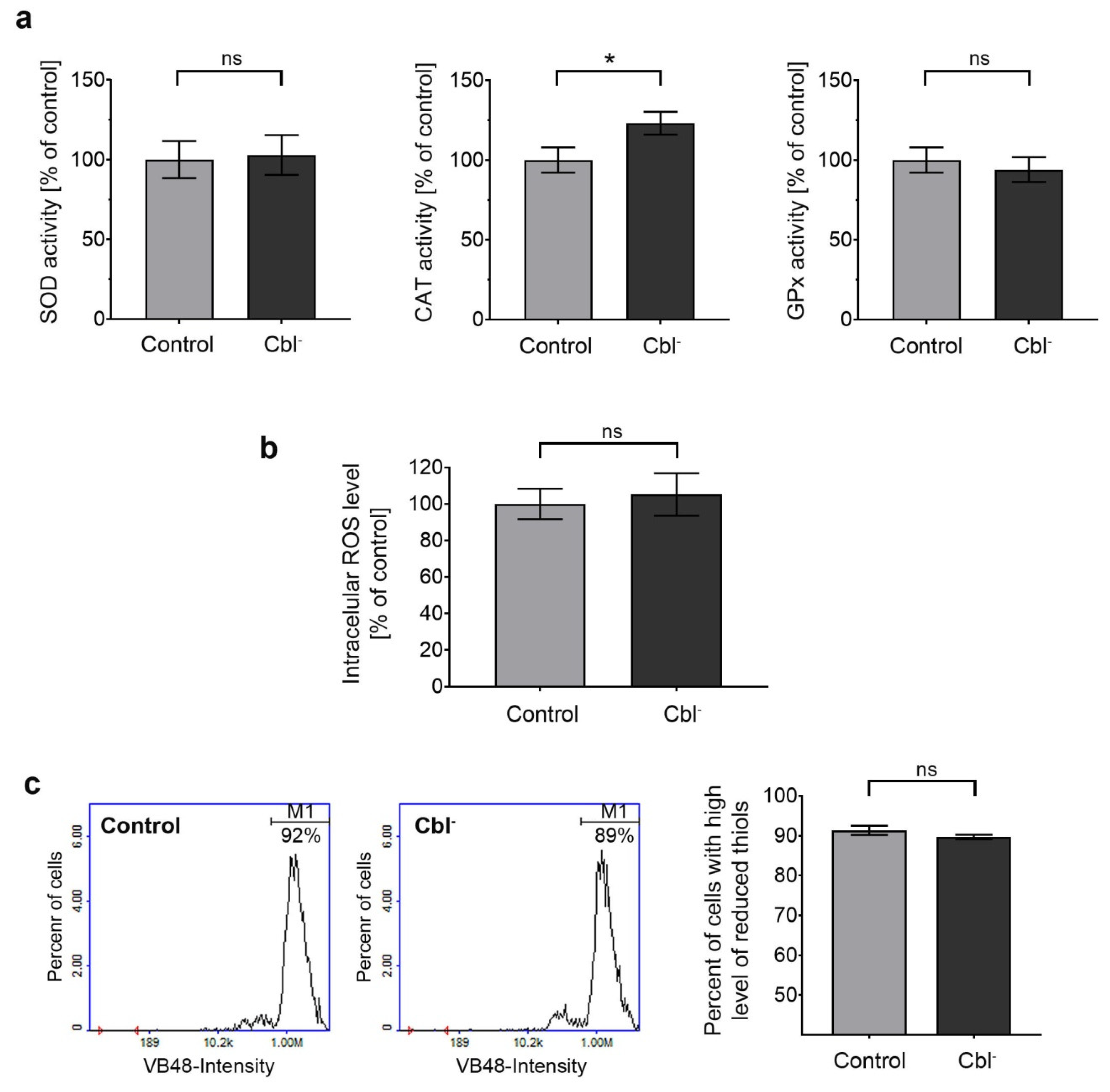
3.6. Redox Homeostasis of Astrocytes under Conditions of Cobalamin Deficiency
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Green, R.; Allen, L.H.; Bjørke-Monsen, A.L.; Brito, A.; Guéant, J.L.; Miller, J.W.; Molloy, A.M.; Nexo, E.; Stabler, S.; Toh, B.H.; et al. Vitamin B12 deficiency. Nat. Rev. Dis. Primers 2017, 3, 17040. [Google Scholar] [CrossRef] [PubMed]
- Dali-Youcef, N.; Andrès, E. An update on cobalamin deficiency in adults. QJM 2009, 102, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Pannérec, A.; Migliavacca, E.; De Castro, A.; Michaud, J.; Karaz, S.; Goulet, L.; Rezzi, S.; Ng, T.P.; Bosco, N.; Larbi, A.; et al. Vitamin B12 deficiency and impaired expression of amnionless during aging. J. Cachexia Sarcopenia Muscle 2018, 9, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Niafar, M.; Hai, F.; Porhomayon, J.; Nader, N.D. The role of metformin on vitamin B12 deficiency: A meta-analysis review. Intern. Emerg. Med. 2015, 10, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.B.; Nagaraja, V.; Kapur, A.; Eslick, G.D. Association between vitamin B12 deficiency and long-term use of acid-lowering agents: A systematic review and meta-analysis. Intern. Med. J. 2015, 45, 409–416. [Google Scholar] [CrossRef]
- Briani, C.; Dalla Torre, C.; Citton, V.; Manara, R.; Pompanin, S.; Binotto, G.; Adami, F. Cobalamin deficiency: Clinical picture and radiological findings. Nutrients 2013, 5, 4521–4539. [Google Scholar] [CrossRef]
- Stabler, S.P.; Brass, E.P.; Marcell, P.D.; Allen, R.H. Inhibition of cobalamin-dependent enzymes by cobalamin analogues in rats. J. Clin. Invest. 1991, 87, 1422–1430. [Google Scholar] [CrossRef]
- Gröber, U.; Kisters, K.; Schmidt, J. Neuroenhancement with vitamin B12-underestimated neurological significance. Nutrients 2013, 5, 5031–5045. [Google Scholar]
- Sadasivan, S.; Friedman, J.H. Vitamin B12 (cobalamin) and Parkinson’s disease. Clin Pract 2012, 9, 353–356. [Google Scholar] [CrossRef]
- Wang, D.; Chen, Y.M.; Ruan, M.H.; Zhou, A.H.; Qian, Y.; Chen, C. Homocysteine inhibits neural stem cells survival by inducing DNA interstrand cross-links via oxidative stress. Neurosci. Lett. 2016, 635, 24–32. [Google Scholar] [CrossRef]
- Tyagi, N.; Sedoris, K.C.; Steed, M.; Ovechkin, A.V.; Moshal, K.S.; Tyagi, S.C. Mechanisms of homocysteine-induced oxidative stress. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2649–H2656. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.M.; Kamynina, E.; Field, M.S.; Stover, P.J. Folate rescues vitamin B12 depletion-induced inhibition of nuclear thymidylate biosynthesis and genome instability. Proc. Natl. Acad. Sci. USA 2017, 114, E4095–E4102. [Google Scholar] [CrossRef] [PubMed]
- Rzepka, Z.; Respondek, M.; Rok, J.; Beberok, A.; Ó Proinsias, K.; Gryko, D.; Wrześniok, D. Vitamin B12 deficiency induces imbalance in melanocytes homeostasis-a cellular basis of hypocobalaminemia pigmentary manifestations. Int. J. Mol. Sci. 2018, 19, 2845. [Google Scholar] [CrossRef] [PubMed]
- Scalabrino, G. The multi-faceted basis of vitamin B12 (cobalamin) neurotrophism in adult central nervous system: Lessons learned from its deficiency. Prog. Neurobiol. 2009, 88, 203–220. [Google Scholar] [CrossRef]
- Ricci, G.; Volpi, L.; Pasquali, L.; Petrozzi, L.; Siciliano, G. Astrocyte–neuron interactions in neurological disorders. J. Biol. Phys. 2009, 35, 317–336. [Google Scholar] [CrossRef]
- Burda, J.E.; Bernstein, A.M.; Sofroniew, M.V. Astrocyte roles in traumatic brain injury. Exp. Neurol. 2016, 275 Pt 3, 305–315. [Google Scholar] [CrossRef]
- Pekny, M.; Pekna, M.; Messing, A.; Steinhäuser, C.; Lee, J.M.; Parpura, V.; Hol, E.M.; Sofroniew, M.V.; Verkhratsky, A. Astrocytes: A central element in neurological diseases. Acta Neuropathol. 2016, 131, 323–345. [Google Scholar] [CrossRef]
- Maragakis, N.J.; Rothstein, J.D. Mechanisms of disease: Astrocytes in neurodegenerative disease. Nat. Clin. Pract. Neurol. 2006, 2, 679–689. [Google Scholar] [CrossRef]
- Scalabrino, G. Cobalamin (vitamin B(12)) in subacute combined degeneration and beyond: Traditional interpretations and novel theories. Exp. Neurol. 2005, 192, 463–479. [Google Scholar] [CrossRef]
- Hultberg, B.; Andersson, A.; Isaksson, A. Metabolism of homocysteine, its relation to the other cellular thiols and its mechanism of cell damage in a cell culture line (human histiocytic cell line U-937). Biochim. Biophys. Acta 1995, 1269, 6–12. [Google Scholar] [CrossRef]
- Beberok, A.; Wrześniok, D.; Minecka, A.; Rok, J.; Delijewski, M.; Rzepka, Z.; Respondek, M.; Buszman, E. Ciprofloxacin-mediated induction of S-phase cell cycle arrest and apoptosis in COLO829 melanoma cells. Pharmacol. Rep. 2018, 70, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Beberok, A.; Rzepka, Z.; Respondek, M.; Rok, J.; Stradowski, M.; Wrześniok, D. Moxifloxacin as an inducer of apoptosis in melanoma cells: A study at the cellular and molecular level. Toxicol. in Vitro 2019, 55, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Wrześniok, D.; Rok, J.; Beberok, A.; Rzepka, Z.; Respondek, M.; Pilawa, B.; Zdybel, M.; Delijewski, M.; Buszman, E. Kanamycin induces free radicals formation in melanocytes: An important factor for aminoglycosides ototoxicity. J. Cell Biochem. 2019, 120, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS using oxidized DCFDA and flow-cytometry. Methods Mol. Biol. 2010, 594, 57–72. [Google Scholar] [PubMed]
- Comini, M.A. Measurement and meaning of cellular thiol: Disufhide redox status. Free Radic. Res. 2016, 50, 246–271. [Google Scholar] [CrossRef]
- Skindersoe, M.E.; Rohde, M.; Kjaerulff, S. A novel and rapid apoptosis assay based on thiol redox status. Cytometry A 2012, 81, 430–436. [Google Scholar] [CrossRef]
- Saraste, A.; Pulkki, K. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc. Res. 2000, 45, 528–537. [Google Scholar] [CrossRef]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (Δψm) in apoptosis; an update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef]
- Tong, C.; Shi, B.; Xiao, X.; Liao, H.; Zheng, Y.; Shen, G.; Tang, D.; Liu, X. An Annexin V-based biosensor for quantitatively detecting early apoptotic cells. Biosens. Bioelectron. 2009, 24, 1777–1782. [Google Scholar] [CrossRef]
- Nakajima, Y.; Kuranaga, E. Caspase-dependent non-apoptotic processes in development. Cell Death Differ. 2017, 24, 1422–1430. [Google Scholar] [CrossRef]
- Guyenet, S.J.; Nguyen, H.T.; Hwang, B.H.; Schwartz, M.W.; Baskin, D.G.; Thalera, J.P. High-fat diet feeding causes rapid, non-apoptotic cleavage of caspase-3 in astrocytes. Brain Res. 2013, 1512, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Aras, R.; Barron, A.M.; Pike, C.J. Caspase activation contributes to astrogliosis. Brain Res. 2012, 1450, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Willems, P.H.G.M.; Rossignol, R.; Dieteren, C.E.J.; Murphy, M.P.; Koopman, W.J.H. Redox homeostasis and mitochondrial dynamics. Cell Metab. 2015, 22, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Próinsias, K.; Giedyk, M.; Gryko, D. Vitamin B12: Chemical modifications. Chem. Soc. Rev. 2013, 42, 6605–6619. [Google Scholar] [CrossRef]
- Matthews, J.H. Cyanocobalamin [c-lactam] inhibits vitamin B12 and causes cytotoxicity in HL60 cells: Methionine protects cells completely. Blood 1997, 89, 4600–4607. [Google Scholar] [CrossRef]
- Sponne, I.E.; Gaire, D.; Stabler, S.P.; Droesch, S.; Barbé, F.M.; Allen, R.H.; Lambert, D.A.; Nicolas, J.P. Inhibition of vitamin B12 metabolism by OH-cobalamin c-lactam in rat oligodendrocytes in culture: A model for studying neuropathy due to vitamin B12 deficiency. Neurosci. Lett. 2000, 288, 191–194. [Google Scholar] [CrossRef]
- Sauer, S.W.; Opp, S.; Haarmann, A.; Okun, J.G.; Kölker, S.; Morath, M.A. Long-term exposure of human proximal tubule cells to hydroxycobalamin[c-lactam] as a possible model to study renal disease in methylmalonic acidurias. J. Inherit. Metab. Dis. 2009, 32, 720–727. [Google Scholar] [CrossRef]
- Shane, B. Folate and vitamin B12 metabolism: Overview and interaction with riboflavin, vitamin B6, and polymorphisms. Food Nutr. Bull 2008, 29, S5–S16. [Google Scholar] [CrossRef]
- Aslinia, F.; Mazza, J.J.; Yale, S.H. Megaloblastic anemia and other causes of macrocytosis. Clin. Med. Res. 2006, 4, 236–241. [Google Scholar] [CrossRef]
- Huang, R.F.; Ho, Y.H.; Lin, H.L.; Wei, J.S.; Liu, T.Z. Folate deficiency induces a cell cycle-specific apoptosis in HepG2 cells. J. Nutr. 1999, 129, 25–31. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- van de Lagemaat, E.E.; de Groot, L.C.; van den Heuvel, E.G. Vitamin B12 in relation to oxidative stress: A Systematic Review. Nutrients 2019, 11, 482. [Google Scholar] [CrossRef] [PubMed]
- Desagher, S.; Glowinski, J.; Premont, J. Astrocytes protect neurons from hydrogen peroxide toxicity. J. Neurosci. 1996, 16, 2553–2562. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rzepka, Z.; Rok, J.; Respondek, M.; Pawlik, J.; Beberok, A.; Gryko, D.; Wrześniok, D. Cobalamin Deficiency: Effect on Homeostasis of Cultured Human Astrocytes. Cells 2019, 8, 1505. https://doi.org/10.3390/cells8121505
Rzepka Z, Rok J, Respondek M, Pawlik J, Beberok A, Gryko D, Wrześniok D. Cobalamin Deficiency: Effect on Homeostasis of Cultured Human Astrocytes. Cells. 2019; 8(12):1505. https://doi.org/10.3390/cells8121505
Chicago/Turabian StyleRzepka, Zuzanna, Jakub Rok, Michalina Respondek, Justyna Pawlik, Artur Beberok, Dorota Gryko, and Dorota Wrześniok. 2019. "Cobalamin Deficiency: Effect on Homeostasis of Cultured Human Astrocytes" Cells 8, no. 12: 1505. https://doi.org/10.3390/cells8121505
APA StyleRzepka, Z., Rok, J., Respondek, M., Pawlik, J., Beberok, A., Gryko, D., & Wrześniok, D. (2019). Cobalamin Deficiency: Effect on Homeostasis of Cultured Human Astrocytes. Cells, 8(12), 1505. https://doi.org/10.3390/cells8121505