Low miR-150-5p and miR-320b Expression Predicts Reduced Survival of COPD Patients

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. miRNAs are Related to 54-Month Survival
2.2. Annotation of miRNAs Correlated with Survival: Higher Expression in Immune Cells
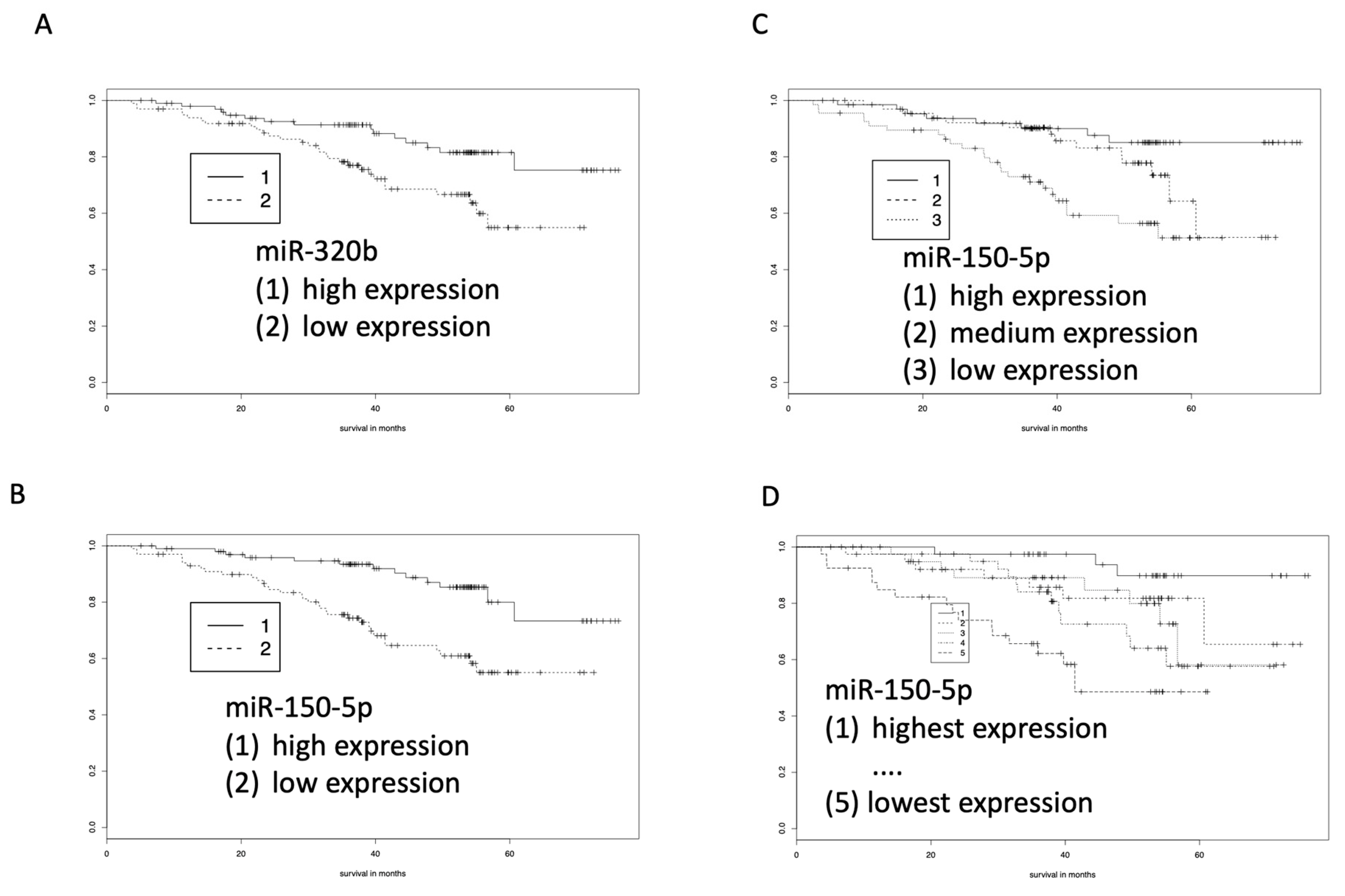
2.3. miR-320b and miR-21-5p as Prognostic Marker for COPD by Kaplan-Meier Estimators
2.4. RT-qPCR Validates miR-320b and miR-150-5p
3. Discussion
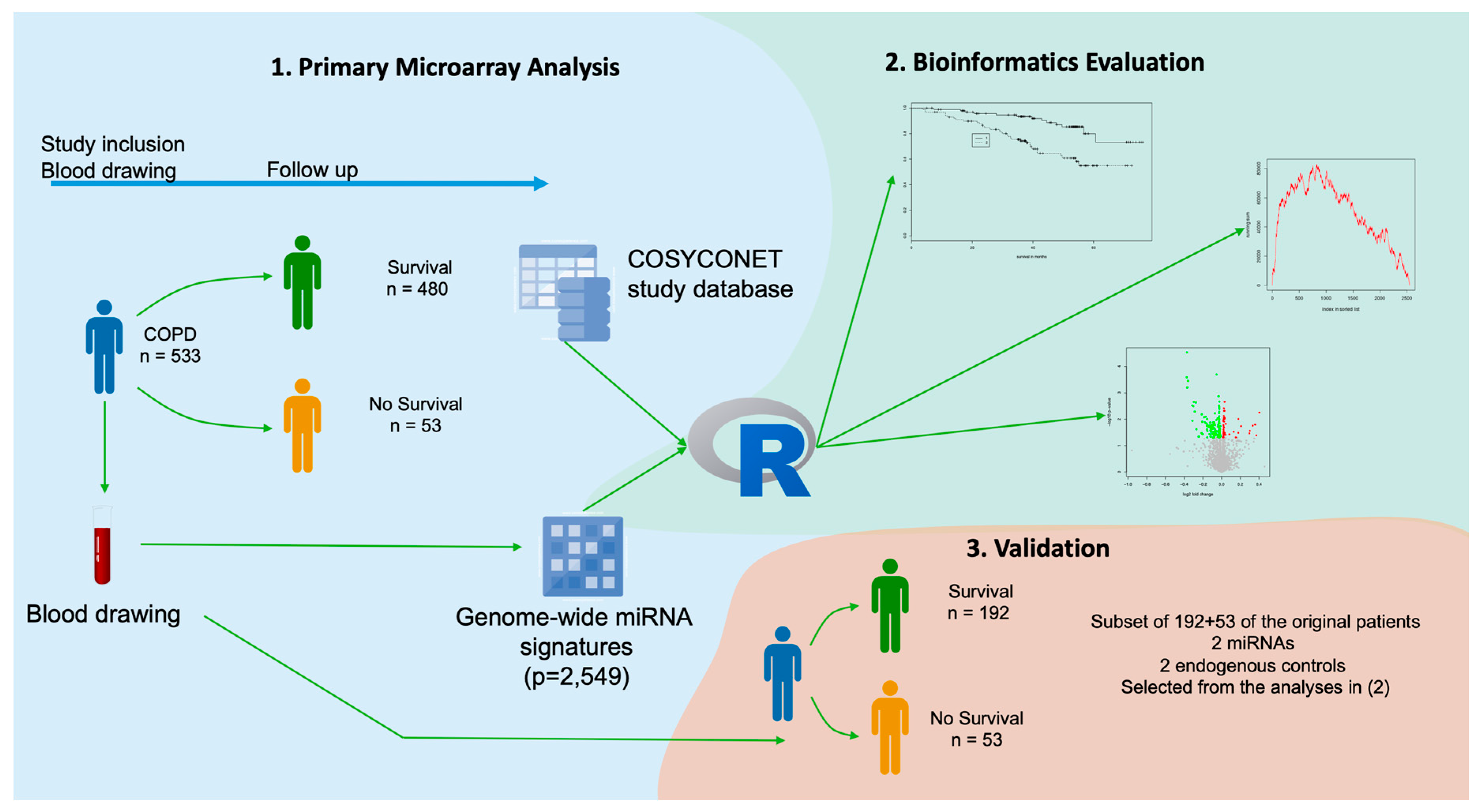
4. Materials and Methods
4.1. Sample Collection and RNA Extraction
4.2. MiRNA Expression Profiles
4.3. RT-qPCR Profiles
4.4. Statistical Analysis
4.5. miRNA Enrichment Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Decramer, M.; Janssens, W. Chronic obstructive pulmonary disease and comorbidities. Lancet Respir. Med. 2013, 1, 73–83. [Google Scholar] [CrossRef]
- Ho, C.H.; Chen, Y.C.; Wang, J.J.; Liao, K.M. Incidence and relative risk for developing cancer among patients with COPD: A nationwide cohort study in Taiwan. BMJ Open 2017, 7, e013195. [Google Scholar] [CrossRef] [PubMed]
- Durham, A.L.; Adcock, I.M. The relationship between COPD and lung cancer. Lung Cancer 2015, 90, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Zakharkina, T.; Heinzel, E.; Koczulla, R.A.; Greulich, T.; Rentz, K.; Pauling, J.K.; Baumbach, J.; Herrmann, M.; Grünewald, C.; Dienemann, H.; et al. Analysis of the Airway Microbiota of Healthy Individuals and Patients with Chronic Obstructive Pulmonary Disease by T-RFLP and Clone Sequencing. PLoS ONE 2013, 8, e68302. [Google Scholar] [CrossRef] [PubMed]
- Buist, A.S.; Vollmer, W.M.; McBurnie, M.A. Worldwide burden of COPD in high- and low-income countries. Part I. The burden of obstructive lung disease (BOLD) initiative. Int. J. Tuberc. Lung Dis. 2008, 12, 703–708. [Google Scholar] [PubMed]
- Chen, W.; Brehm, J.M.; Manichaikul, A.; Cho, M.H.; Boutaoui, N.; Yan, Q.; Burkart, K.M.; Enright, P.L.; Rotter, J.I.; Petersen, H.; et al. A genome-wide association study of chronic obstructive pulmonary disease in Hispanics. Ann. Am. Thorac. Soc. 2015, 12, 340–348. [Google Scholar] [CrossRef]
- Cho, M.H.; Castaldi, P.J.; Wan, E.S.; Siedlinski, M.; Hersh, C.P.; Demeo, D.L.; Himes, B.E.; Sylvia, J.S.; Klanderman, B.J.; Ziniti, J.P.; et al. A genome-wide association study of COPD identifies a susceptibility locus on chromosome 19q13. Hum. Mol. Genet. 2012, 21, 947–957. [Google Scholar] [CrossRef]
- Wain, L.V.; Shrine, N.; Artigas, M.S.; Erzurumluoglu, A.M.; Noyvert, B.; Bossini-Castillo, L.; Obeidat, M.; Henry, A.P.; Portelli, M.A.; Hall, R.J.; et al. Genome-wide association analyses for lung function and chronic obstructive pulmonary disease identify new loci and potential druggable targets. Nat. Genet. 2017, 49, 416–425. [Google Scholar] [CrossRef]
- Kowalczyk, M.S.; Higgs, D.R.; Gingeras, T.R. Molecular biology: RNA discrimination. Nature 2012, 482, 310–311. [Google Scholar] [CrossRef]
- Jankowsky, E.; Harris, M.E. Specificity and nonspecificity in RNA-protein interactions. Nat. Rev. Mol. Cell Boil. 2015, 16, 533–544. [Google Scholar] [CrossRef]
- Dang, X.; Qu, X.; Wang, W.; Liao, C.; Li, Y.; Zhang, X.; Xu, D.; Baglole, C.J.; Shang, D.; Chang, Y. Bioinformatic analysis of microRNA and mRNA Regulation in peripheral blood mononuclear cells of patients with chronic obstructive pulmonary disease. Respir. Res. 2017, 18, 4. [Google Scholar] [CrossRef] [PubMed]
- Kara, M.; Kirkil, G.; Kalemci, S. Differential Expression of MicroRNAs in Chronic Obstructive Pulmonary Disease. Adv. Clin. Exp. Med. 2016, 25, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Leidinger, P.; Keller, A.; Borries, A.; Huwer, H.; Rohling, M.; Huebers, J.; Lenhof, H.-P.; Meese, E. Specific peripheral miRNA profiles for distinguishing lung cancer from COPD. Lung Cancer 2011, 74, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Fehlmann, T.; Ludwig, N.; Kahraman, M.; Laufer, T.; Backes, C.; Vogelmeier, C.; Diener, C.; Biertz, F.; Herr, C.; et al. Genome-wide MicroRNA Expression Profiles in COPD: Early Predictors for Cancer Development. Genomics Proteomics Bioinf. 2018, 16, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Karch, A.; Vogelmeier, C.; Welte, T.; Bals, R.; Kauczor, H.-U.; Biederer, J.; Heinrich, J.; Schulz, H.; Gläser, S.; Holle, R.; et al. The German COPD cohort COSYCONET: Aims, methods and descriptive analysis of the study population at baseline. Respir. Med. 2016, 114, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Miniati, M.; Monti, S.; Pavlickova, I.; Bottai, M. Survival in COPD: Impact of Lung Dysfunction and Comorbidities. Medicine 2014, 93, e76. [Google Scholar] [CrossRef] [PubMed]
- Backes, C.; Khaleeq, Q.T.; Meese, E.; Keller, A. miEAA: microRNA enrichment analysis and annotation. Nucleic Acids Res. 2016, 44, W110–W116. [Google Scholar] [CrossRef]
- Zheng, J.; Lin, Z.; Dong, P.; Lu, Z.; Gao, S.; Chen, X.; Wu, C.; Yu, F. Activation of hepatic stellate cells is suppressed by microRNA-150. Int. J. Mol. Med. 2013, 32, 17–24. [Google Scholar] [CrossRef]
- Yao, J.; Liang, L.H.; Zhang, Y.; Ding, J.; Tian, Q.; Li, J.J.; He, X.H. GNAI1 Suppresses Tumor Cell Migration and Invasion and is Post-Transcriptionally Regulated by Mir-320a/c/d in Hepatocellular Carcinoma. Cancer Boil. Med. 2012, 9, 234–241. [Google Scholar]
- Wang, B.; Jha, J.C.; Hagiwara, S.; McClelland, A.D.; Jandeleit-Dahm, K.; Thomas, M.C.; Cooper, M.E.; Kantharidis, P. Transforming growth factor-beta1-mediated renal fibrosis is dependent on the regulation of transforming growth factor receptor 1 expression by let-7b. Kidney Int. 2014, 85, 352–361. [Google Scholar] [CrossRef]
- Kimura, K.; Hohjoh, H.; Fukuoka, M.; Sato, W.; Oki, S.; Tomi, C.; Yamaguchi, H.; Kondo, T.; Takahashi, R.; Yamamura, T. Circulating exosomes suppress the induction of regulatory T cells via let-7i in multiple sclerosis. Nat. Commun. 2018, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Meinders, M.; Kulu, D.I.; van de Werken, H.J.; Hoogenboezem, M.; Janssen, H.; Brouwer, R.W.; van Ijcken, W.F.; Rijkers, E.J.; Demmers, J.A.; Kruger, I.; et al. Sp1/Sp3 transcription factors regulate hallmarks of megakaryocyte maturation and platelet formation and function. Blood 2015, 125, 1957–1967. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.F.; Singh, S.P.; Cantu, M.; Chen, L.; Zhang, H.H.; Farber, J.M. Differentiation of human T cells alters their repertoire of G protein alpha-subunits. J. Biol. Chem. 2010, 285, 35537–35550. [Google Scholar] [CrossRef] [PubMed]
- Backes, C.; Ludwig, N.; Leidinger, P.; Huwer, H.; Tenzer, S.; Fehlmann, T.; Franke, A.; Meese, E.; Lenhof, H.-P.; Keller, A. Paired proteomics, transcriptomics and miRNomics in non-small cell lung cancers: Known and novel signaling cascades. Oncotarget 2016, 7, 71514–71525. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zeng, K.; Xu, M.; Hu, X.; Liu, X.; Xu, T.; He, B.; Pan, Y.; Sun, H.; Wang, S. SP1-induced lncRNA-ZFAS1 contributes to colorectal cancer progression via the miR-150-5p/VEGFA axis. Cell Death Dis. 2018, 9, 982. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Katada, K.; Okato, A.; Arai, T.; Idichi, T.; Osako, Y.; Okamoto, Y.; Seki, N. Antitumor miR-150-5p and miR-150-3p inhibit cancer cell aggressiveness by targeting SPOCK1 in head and neck squamous cell carcinoma. Auris Nasus Larynx 2018, 45, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Zhang, H.; Niu, Y.; Wu, Y.; Sun, W.; Li, H.; Kong, J.; Ding, K.; Shen, H.M.; Wu, H.; et al. Long non-coding RNA linc00673 regulated non-small cell lung cancer proliferation, migration, invasion and epithelial mesenchymal transition by sponging miR-150-5p. Mol. Cancer 2017, 16, 118. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xie, J.; Shen, C.; Cheng, D.; Shi, Y.; Wu, Z.; Zhan, Q.; Deng, X.; Chen, H.; Shen, B.; et al. miR-150-5p inhibits hepatoma cell migration and invasion by targeting MMP14. PLoS ONE 2014, 9, e115577. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.L.; Du, H.; Liu, Y.L.; Huang, Y.T.; Wang, G.-H.; Zhang, X.; Chen, S.H.; Zhou, H.H. Low expression of microRNA-320b correlates with tumorigenesis and unfavorable prognosis in glioma. Oncol. Rep. 2017, 38, 959–966. [Google Scholar] [CrossRef]
- Tadano, T.; Kakuta, Y.; Hamada, S.; Shimodaira, Y.; Kuroha, M.; Kawakami, Y.; Kimura, T.; Shiga, H.; Endo, K.; Masamune, A.; et al. MicroRNA-320 family is downregulated in colorectal adenoma and affects tumor proliferation by targeting CDK6. World J. Gastrointest. Oncol. 2016, 8, 532–542. [Google Scholar] [CrossRef]
- Wang, H.; Cao, F.; Li, X.; Miao, H.; E, J.; Xing, J.; Fu, C.G. miR-320b suppresses cell proliferation by targeting c-Myc in human colorectal cancer cells. BMC Cancer 2015, 15, 748. [Google Scholar] [CrossRef] [PubMed]
- Sekine, Y.; Katsura, H.; Koh, E.; Hiroshima, K.; Fujisawa, T. Early detection of COPD is important for lung cancer surveillance. Eur. Respir. J. 2012, 39, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keller, A.; Ludwig, N.; Fehlmann, T.; Kahraman, M.; Backes, C.; Kern, F.; Vogelmeier, C.F.; Diener, C.; Fischer, U.; Biertz, F.; et al. Low miR-150-5p and miR-320b Expression Predicts Reduced Survival of COPD Patients. Cells 2019, 8, 1162. https://doi.org/10.3390/cells8101162
Keller A, Ludwig N, Fehlmann T, Kahraman M, Backes C, Kern F, Vogelmeier CF, Diener C, Fischer U, Biertz F, et al. Low miR-150-5p and miR-320b Expression Predicts Reduced Survival of COPD Patients. Cells. 2019; 8(10):1162. https://doi.org/10.3390/cells8101162
Chicago/Turabian StyleKeller, Andreas, Nicole Ludwig, Tobias Fehlmann, Mustafa Kahraman, Christina Backes, Fabian Kern, Claus F. Vogelmeier, Caroline Diener, Ulrike Fischer, Frank Biertz, and et al. 2019. "Low miR-150-5p and miR-320b Expression Predicts Reduced Survival of COPD Patients" Cells 8, no. 10: 1162. https://doi.org/10.3390/cells8101162
APA StyleKeller, A., Ludwig, N., Fehlmann, T., Kahraman, M., Backes, C., Kern, F., Vogelmeier, C. F., Diener, C., Fischer, U., Biertz, F., Herr, C., Jörres, R. A., Lenhof, H.-P., Bals, R., & Meese, E. (2019). Low miR-150-5p and miR-320b Expression Predicts Reduced Survival of COPD Patients. Cells, 8(10), 1162. https://doi.org/10.3390/cells8101162