Revisiting Centrioles in Nematodes—Historic Findings and Current Topics

 and
and
Abstract
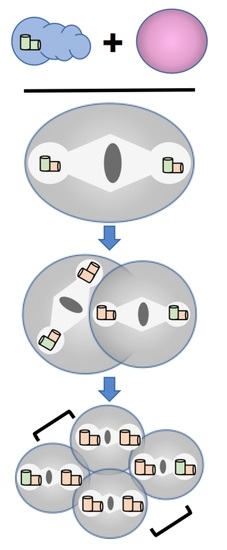
{kind=link}
{kind=link}
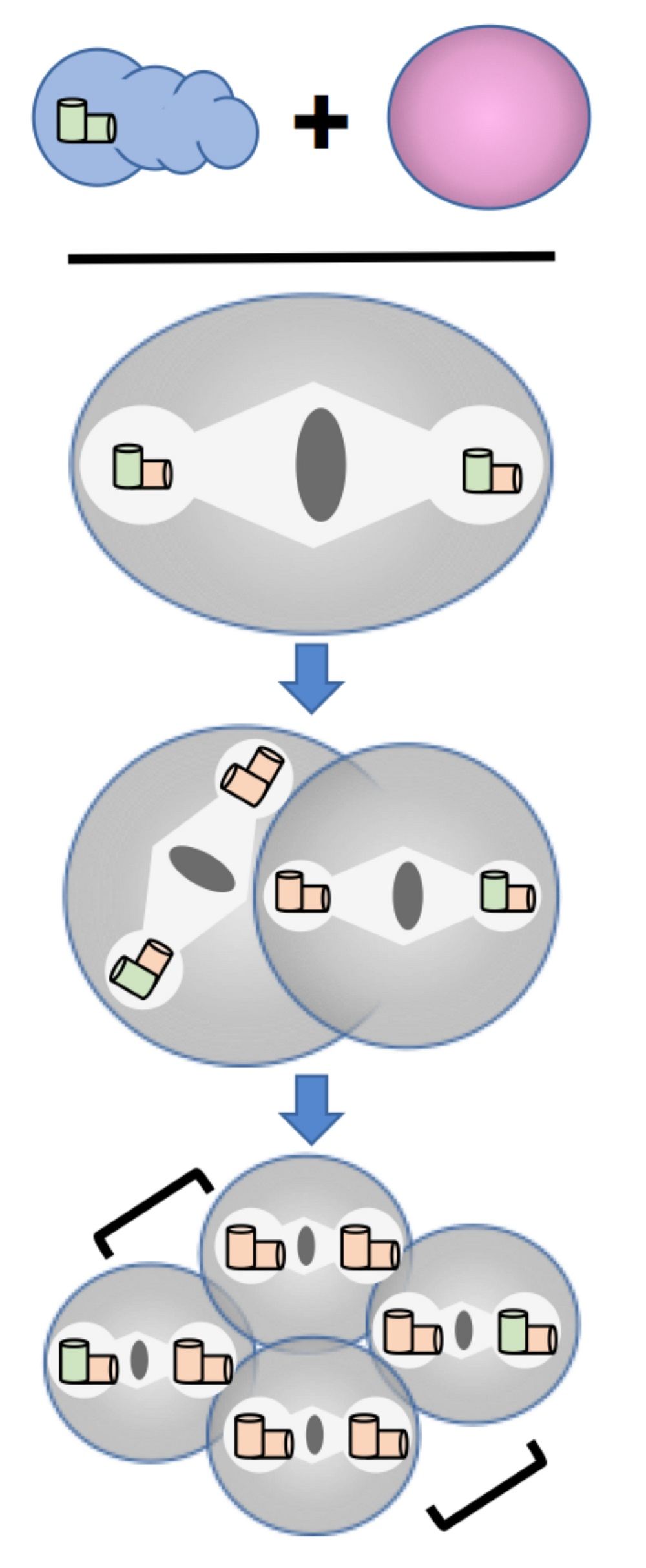
{kind=link}
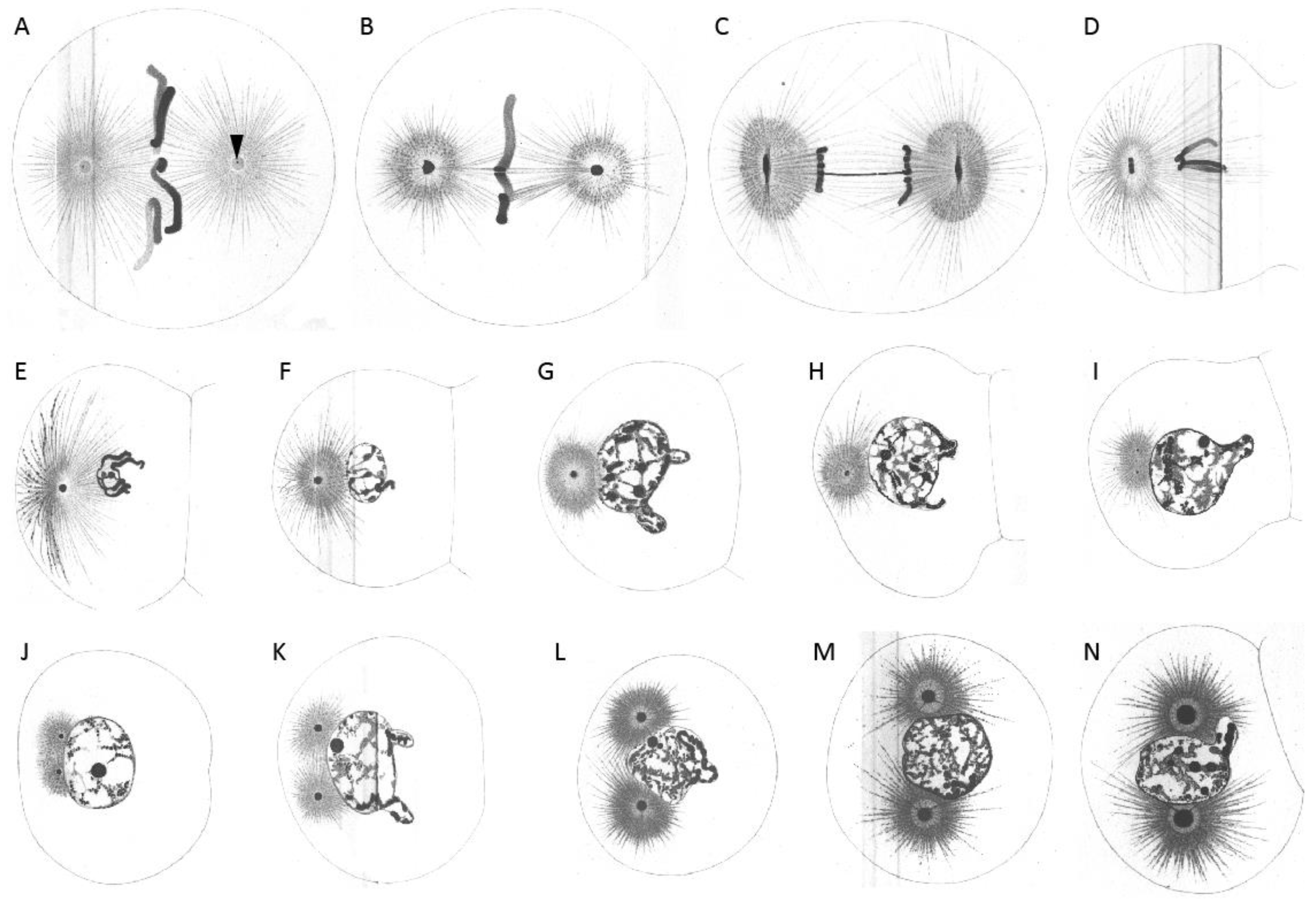
{kind=link}
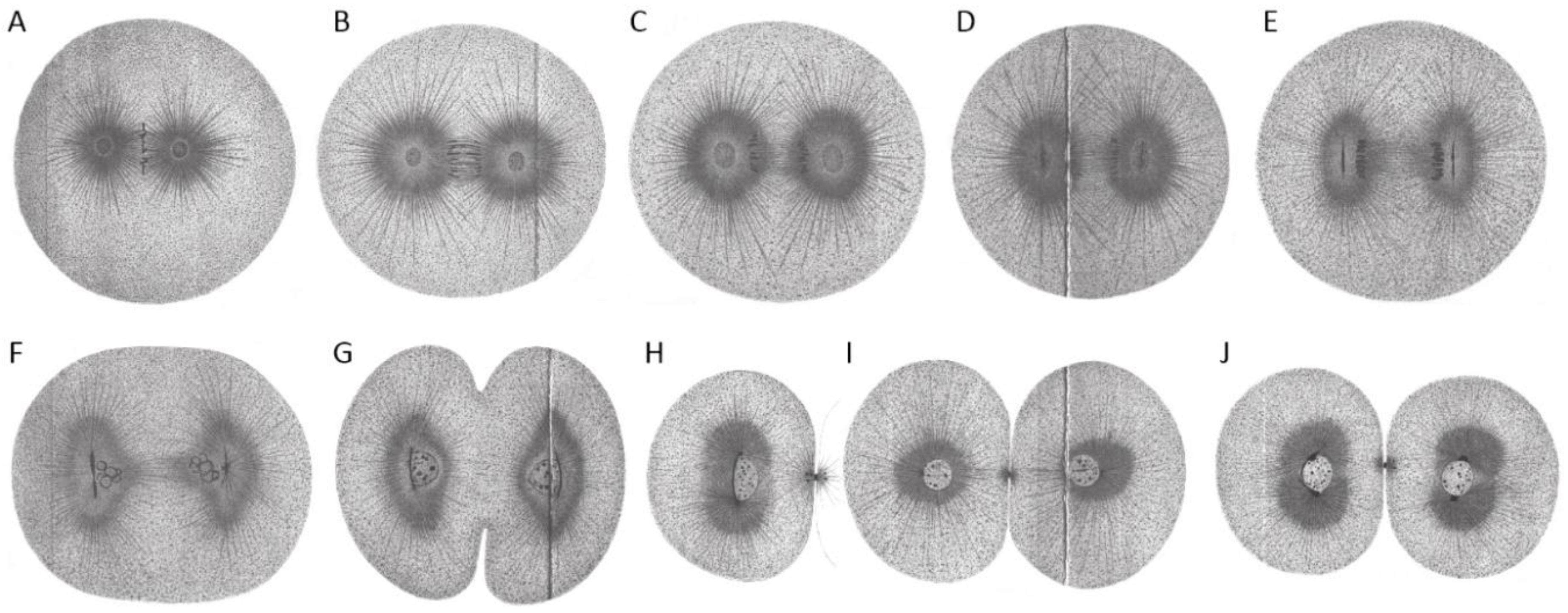
{kind=link}
{kind=link}
1. Introduction
“In der ruhenden Zelle besteht ausserhalb des Kerns in der Zellsubstanz ein specifisches Körperchen, das ich ››Centrosoma‹‹ […] nenne.”[1]
“In the resting cell outside of the nucleus there is a specific corpuscle, which I will term ››Centrosoma‹‹.”[1]
“Gleichwohl enthält nun dieses Gebilde […] noch ein viel kleineres Körperchen, das von mir zuerst im Ascaris-Ei aufgefundene ››Centralkorn‹‹. Ich gebrauche für dieses Gebilde fortan neben dem Wort ››Centralkorn‹‹ den […] Terminus ››Centriol‹‹ […].”[2]
“There is also a much smaller little corpucle, the ››Centralkorn‹‹, which I discovered in Ascaris eggs. In parallel to the word ››Centralkorn‹‹, I will use the term ››Centriol‹‹ […].”[2]
2. The Discovery of the Centrosome by Theodor Boveri—Early Observations
3. Return to Nematodes
4. The Structure of C. elegans Germ-Line Centrioles
5. The Core Centriole Assembly Pathway
6. Other Centriole Assembly Factors
7. PCM Recruitment
8. Concluding Remarks
Funding
Conflicts of Interest
References
- Boveri, T. Ueber den Antheil des Spermatozoon an der Theilung des Eies. Sitzungsber. Ges. Morph. Physiol. München 1887, 3, 151–164. [Google Scholar]
- Ueber die Natur der Centrosomen. Available online: https://www.biodiversitylibrary.org/page/6303288#page/7/mode/1up (accessed on 1 August 2018).
- Baltzer, F. Theodor Boveri—Leben und Werk eines grossen Biologen 1862–1915; Wissenschaftliche Verlagsgesellschaft m.b.H.: Stuttgart, Germany, 1962; pp. 1–194. [Google Scholar]
- Scheer, U. Historical roots of centrosome research: Discovery of boveri’s microscope slides in Würzburg. Philos. Trans. R. Soc. Lond B. Biol. Sci. 2014, 369, 20130469. [Google Scholar] [CrossRef] [PubMed]
- Boveri, T. Ueber die Bedeutung der Richtungskörper. Sitzungsber. Ges. Morph. Physiol. München 1886, 2, 101–106. [Google Scholar]
- Die Bildung der Richtungskörper bei Ascaris Megalocephala und Ascaris Lumbricoides. Available online: https://www.biodiversitylibrary.org/page/6562432#page/7/mode/1up (accessed on 1 August 2018).
- Boveri, T. Ueber die Befruchtung der Eier von Ascaris megalocephala. Sitzungsber. Ges. Morph. Physiol. München 1887, 3, 71–80. [Google Scholar]
- van Beneden, E.; Neyt, A. Nouvelle recherches sur la fécondation et la division mitosique chez l’Ascaride mégalocéphale. Bull. Acad. Royale Belgique 3éme sér 1887, 14, 215–295. [Google Scholar]
- Heidenhain, M. Ueber die Mikrocentren mehrkerniger Riesenzellen sowie über die Centralkörperfrage im Allgemeinen. Morphol. Arbeiten 1897, 1, 225–280. [Google Scholar]
- Boveri, T. Ueber das Verhalten der Centrosomen bei der Befruchtung des Seeigel-Eies, nebst allgemeinen Bemerkungen über Centrosomen und Verwandtes. Verhandl. Phys.-Med. Ges. Würzburg 1895, 29, 1–75. [Google Scholar]
- Boveri, T. Zur Physiologie der Kern- und Zellteilung; Verlag und Druck der Stahel’schen k. Hof- und Universitätsbuch- und Kunsthandlung: Würzburg, Germany, 1896; pp. 133–151. [Google Scholar]
- Sugioka, K.; Hamill, D.R.; Lowry, J.B.; McNeely, M.E.; Enrick, M.; Richter, A.C.; Kiebler, L.E.; Priess, J.R.; Bowerman, B. Centriolar SAS-7 acts upstream of SPD-2 to regulate centriole assembly and pericentriolar material formation. Elife 2017, 6, e20353. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, E.T.; McDonald, K.L.; Mantler, J.; McIntosh, J.R.; Hyman, A.A.; Müller-Reichert, T. Morphologically distinct microtubule ends in the mitotic centrosome of Caenorhabditis elegans. J. Cell Biol. 2003, 163, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, L.; O’Toole, E.; Schwager, A.; Hyman, A.A.; Müller-Reichert, T. Centriole assembly in Caenorhabditis elegans. Nature 2006, 444, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Kitagawa, D. Ultrastructural diversity between centrioles of eukaryotes. J. Biochem. 2018, 164, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, D.; Vakonakis, I.; Olieric, N.; Hilbert, M.; Keller, D.; Olieric, V.; Bortfeld, M.; Erat, M.C.; Fluckiger, I.; Gonczy, P.; et al. Structural basis of the 9-fold symmetry of centrioles. Cell 2011, 144, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Van Breugel, M.; Hirono, M.; Andreeva, A.; Yanagisawa, H.A.; Yamaguchi, S.; Nakazawa, Y.; Morgner, N.; Petrovich, M.; Ebong, I.O.; Robinson, C.V.; et al. Structures of SAS-6 suggest its organization in centrioles. Science 2011, 331, 1196–1199. [Google Scholar] [CrossRef] [PubMed]
- Guichard, P.; Desfosses, A.; Maheshwari, A.; Hachet, V.; Dietrich, C.; Brune, A.; Ishikawa, T.; Sachse, C.; Gonczy, P. Cartwheel architecture of Trichonympha basal body. Science 2012, 337, 553. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, M.; Noga, A.; Frey, D.; Hamel, V.; Guichard, P.; Kraatz, S.H.; Pfreundschuh, M.; Hosner, S.; Fluckiger, I.; Jaussi, R.; et al. SAS-6 engineering reveals interdependence between cartwheel and microtubules in determining centriole architecture. Nat. Cell Biol. 2016, 18, 393–403. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, K.F.; Caron, C.; Kopish, K.R.; Hurd, D.D.; Kemphues, K.J.; Li, Y.; White, J.G. The C. elegans zyg-1 gene encodes a regulator of centrosome duplication with distinct maternal and paternal roles in the embryo. Cell 2001, 105, 547–558. [Google Scholar] [CrossRef]
- Malone, C.J.; Misner, L.; Le Bot, N.; Tsai, M.C.; Campbell, J.M.; Ahringer, J.; White, J.G. The C. elegans hook protein, ZYG-12, mediates the essential attachment between the centrosome and nucleus. Cell 2003, 115, 825–836. [Google Scholar] [CrossRef]
- Wolf, N.; Hirsh, D.; McIntosh, J.R. Spermatogenesis in males of the free-living nematode, Caenorhabditis elegans. J. Ultrastruct. Res. 1978, 63, 155–169. [Google Scholar] [CrossRef]
- Goodenough, U.W.; StClair, H.S. Bald-2: A mutation affecting the formation of doublet and triplet sets of microtubules in Chlamydomonas reinhardtii. J. Cell Biol. 1975, 66, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Dutcher, S.K.; Trabuco, E.C. The UNI3 gene is required for assembly of basal bodies of Chlamydomonas and encodes delta-tubulin, a new member of the tubulin superfamily. Mol. Biol. Cell 1998, 9, 1293–1308. [Google Scholar] [CrossRef] [PubMed]
- Dupuis-Williams, P.; Fleury-Aubusson, A.; de Loubresse, N.G.; Geoffroy, H.; Vayssie, L.; Galvani, A.; Espigat, A.; Rossier, J. Functional role of epsilon-tubulin in the assembly of the centriolar microtubule scaffold. J. Cell Biol. 2002, 158, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Dutcher, S.K.; Morrissette, N.S.; Preble, A.M.; Rackley, C.; Stanga, J. Epsilon-tubulin is an essential component of the centriole. Mol. Biol. Cell 2002, 13, 3859–3869. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, C.; Wickstead, B.; McKean, P.G.; Gull, K. Basal body and flagellum mutants reveal a rotational constraint of the central pair microtubules in the axonemes of trypanosomes. J. Cell Sci. 2006, 119, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Ross, I.; Clarissa, C.; Giddings, T.H., Jr.; Winey, M. Epsilon-tubulin is essential in Tetrahymena thermophila for the assembly and stability of basal bodies. J. Cell Sci. 2013, 126, 3441–3451. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.T.; Kong, D.; Hoerner, C.R.; Loncarek, J.; Stearns, T. Centriole triplet microtubules are required for stable centriole formation and inheritance in human cells. Elife 2017, 6, e29061. [Google Scholar] [CrossRef] [PubMed]
- Serwas, D.; Su, T.Y.; Roessler, M.; Wang, S.; Dammermann, A. Centrioles initiate cilia assembly but are dispensable for maturation and maintenance in C. elegans. J. Cell. Biol. 2017, 216, 1659–1671. [Google Scholar] [CrossRef] [PubMed]
- Cell Division. Available online: http://www.wormbook.org/chapters/www_celldivision/celldivision.html (accessed on 01 August 2018).
- Redemann, S.; Baumgart, J.; Lindow, N.; Shelley, M.; Nazockdast, E.; Kratz, A.; Prohaska, S.; Brugues, J.; Furthauer, S.; Müller-Reichert, T. C. elegans chromosomes connect to centrosomes by anchoring into the spindle network. Nat. Commun. 2017, 8, 15288. [Google Scholar] [CrossRef] [PubMed]
- Tanos, B.E.; Yang, H.J.; Soni, R.; Wang, W.J.; Macaluso, F.P.; Asara, J.M.; Tsou, M.F. Centriole distal appendages promote membrane docking, leading to cilia initiation. Genes Dev. 2013, 27, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Xia, Y.; Zhang, D.; Wang, S.; Bao, Y.; He, R.; Teng, J.; Chen, J. Hierarchical assembly of centriole subdistal appendages via centrosome binding proteins CCDC120 and CCDC68. Nat. Commun. 2017, 8, 15057. [Google Scholar] [CrossRef] [PubMed]
- Sonnichsen, B.; Koski, L.B.; Walsh, A.; Marschall, P.; Neumann, B.; Brehm, M.; Alleaume, A.M.; Artelt, J.; Bettencourt, P.; Cassin, E.; et al. Full-genome RNAi profiling of early embryogenesis in Caenorhabditis elegans. Nature 2005, 434, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Dammermann, A.; Müller-Reichert, T.; Pelletier, L.; Habermann, B.; Desai, A.; Oegema, K. Centriole assembly requires both centriolar and pericentriolar material proteins. Dev. Cell 2004, 7, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Song, M.H.; Liu, Y.; Anderson, D.E.; Jahng, W.J.; O’Connell, K.F. Protein phosphatase 2A-SUR-6/B55 regulates centriole duplication in C. elegans by controlling the levels of centriole assembly factors. Dev. Cell 2011, 20, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Bettencourt-Dias, M.; Rodrigues-Martins, A.; Carpenter, L.; Riparbelli, M.; Lehmann, L.; Gatt, M.K.; Carmo, N.; Balloux, F.; Callaini, G.; Glover, D.M. SAK/PLK4 is required for centriole duplication and flagella development. Curr. Biol. 2005, 15, 2199–2207. [Google Scholar] [CrossRef] [PubMed]
- Habedanck, R.; Stierhof, Y.D.; Wilkinson, C.J.; Nigg, E.A. The polo kinase Plk4 functions in centriole duplication. Nat. Cell Biol. 2005, 7, 1140–1146. [Google Scholar] [CrossRef] [PubMed]
- Kleylein-Sohn, J.; Westendorf, J.; Le Clech, M.; Habedanck, R.; Stierhof, Y.D.; Nigg, E.A. Plk4-induced centriole biogenesis in human cells. Dev. Cell 2007, 13, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Santos, Z.; Machado, P.; Branco, P.; Tavares-Cadete, F.; Rodrigues-Martins, A.; Pereira-Leal, J.B.; Bettencourt-Dias, M. Stepwise evolution of the centriole-assembly pathway. J. Cell Sci. 2010, 123, 1414–1426. [Google Scholar] [CrossRef] [PubMed]
- Hodges, M.E.; Scheumann, N.; Wickstead, B.; Langdale, J.A.; Gull, K. Reconstructing the evolutionary history of the centriole from protein components. J. Cell Sci. 2010, 123, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Jana, S.C.; Bazan, J.F.; Bettencourt-Dias, M. Polo boxes come out of the crypt: A new view of PLK function and evolution. Structure 2012, 20, 1801–1804. [Google Scholar] [CrossRef] [PubMed]
- Shimanovskaya, E.; Viscardi, V.; Lesigang, J.; Lettman, M.M.; Qiao, R.; Svergun, D.I.; Round, A.; Oegema, K.; Dong, G. Structure of the C. elegans ZYG-1 cryptic polo box suggests a conserved mechanism for centriolar docking of Plk4 kinases. Structure 2014, 22, 1090–1104. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Park, J.E.; Kim, T.S.; Kim, J.H.; Kwak, M.J.; Ku, B.; Tian, L.; Murugan, R.N.; Ahn, M.; Komiya, S.; et al. Molecular basis for unidirectional scaffold switching of human Plk4 in centriole biogenesis. Nat. Struct. Mol. Biol. 2014, 21, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Delattre, M.; Canard, C.; Gonczy, P. Sequential protein recruitment in C. elegans centriole formation. Curr. Biol. 2006, 16, 1844–1849. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.A.; Kopish, K.R.; Zipperlen, P.; Ahringer, J.; O’Connell, K.F. Centrosome maturation and duplication in C. elegans require the coiled-coil protein SPD-2. Dev. Cell 2004, 6, 511–523. [Google Scholar] [CrossRef]
- Pelletier, L.; Ozlu, N.; Hannak, E.; Cowan, C.; Habermann, B.; Ruer, M.; Müller-Reichert, T.; Hyman, A.A. The Caenorhabditis elegans centrosomal protein SPD-2 is required for both pericentriolar material recruitment and centriole duplication. Curr. Biol. 2004, 14, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Dix, C.I.; Raff, J.W. Drosophila Spd-2 recruits PCM to the sperm centriole, but is dispensable for centriole duplication. Curr. Biol. 2007, 17, 1759–1764. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Ferreria, M.A.; Rath, U.; Buster, D.W.; Chanda, S.K.; Caldwell, J.S.; Rines, D.R.; Sharp, D.J. Human Cep192 is required for mitotic centrosome and spindle assembly. Curr. Biol. 2007, 17, 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, M.G.; Bucciarelli, E.; Bonaccorsi, S.; Gatti, M. Drosophila SPD-2 is an essential centriole component required for PCM recruitment and astral-microtubule nucleation. Curr. Biol. 2008, 18, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Lawo, S.; Bird, A.; Pinchev, D.; Ralph, A.; Richter, C.; Müller-Reichert, T.; Kittler, R.; Hyman, A.A.; Pelletier, L. The mammalian SPD-2 ortholog Cep192 regulates centrosome biogenesis. Curr. Biol. 2008, 18, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Sonnen, K.F.; Gabryjonczyk, A.M.; Anselm, E.; Stierhof, Y.D.; Nigg, E.A. Human Cep192 and Cep152 cooperate in Plk4 recruitment and centriole duplication. J. Cell Sci. 2013, 126, 3223–3233. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.S.; Wilkinson, C.J.; Mayor, T.; Mortensen, P.; Nigg, E.A.; Mann, M. Proteomic characterization of the human centrosome by protein correlation profiling. Nature 2003, 426, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Cizmecioglu, O.; Arnold, M.; Bahtz, R.; Settele, F.; Ehret, L.; Haselmann-Weiss, U.; Antony, C.; Hoffmann, I. Cep152 acts as a scaffold for recruitment of Plk4 and CPAP to the centrosome. J. Cell Biol. 2010, 191, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Park, J.E.; Shukla, A.; Choi, S.; Murugan, R.N.; Lee, J.H.; Ahn, M.; Rhee, K.; Bang, J.K.; Kim, B.Y.; et al. Hierarchical recruitment of Plk4 and regulation of centriole biogenesis by two centrosomal scaffolds, Cep192 and Cep152. Proc. Natl. Acad. Sci. USA 2013, 110, E4849–E4857. [Google Scholar] [CrossRef] [PubMed]
- Leidel, S.; Delattre, M.; Cerutti, L.; Baumer, K.; Gonczy, P. SAS-6 defines a protein family required for centrosome duplication in C. elegans and in human cells. Nat. Cell Biol. 2005, 7, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Delattre, M.; Leidel, S.; Wani, K.; Baumer, K.; Bamat, J.; Schnabel, H.; Feichtinger, R.; Schnabel, R.; Gonczy, P. Centriolar SAS-5 is required for centrosome duplication in C. elegans. Nat. Cell Biol. 2004, 6, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Stevens, N.R.; Dobbelaere, J.; Brunk, K.; Franz, A.; Raff, J.W. Drosophila Ana2 is a conserved centriole duplication factor. J. Cell Biol. 2010, 188, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Stevens, N.R.; Roque, H.; Raff, J.W. DSas-6 and Ana2 coassemble into tubules to promote centriole duplication and engagement. Dev. Cell 2010, 19, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Vulprecht, J.; David, A.; Tibelius, A.; Castiel, A.; Konotop, G.; Liu, F.; Bestvater, F.; Raab, M.S.; Zentgraf, H.; Izraeli, S.; et al. Stil is required for centriole duplication in human cells. J. Cell Sci. 2012, 125, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Cottee, M.A.; Muschalik, N.; Johnson, S.; Leveson, J.; Raff, J.W.; Lea, S.M. The homo-oligomerisation of both Sas-6 and Ana2 is required for efficient centriole assembly in flies. Elife 2015, 4, e07236. [Google Scholar] [CrossRef] [PubMed]
- Rogala, K.B.; Dynes, N.J.; Hatzopoulos, G.N.; Yan, J.; Pong, S.K.; Robinson, C.V.; Deane, C.M.; Gonczy, P.; Vakonakis, I. The Caenorhabditis elegans protein SAS-5 forms large oligomeric assemblies critical for centriole formation. Elife 2015, 4, e07410. [Google Scholar] [CrossRef] [PubMed]
- Lettman, M.M.; Wong, Y.L.; Viscardi, V.; Niessen, S.; Chen, S.H.; Shiau, A.K.; Zhou, H.; Desai, A.; Oegema, K. Direct binding of SAS-6 to ZYG-1 recruits SAS-6 to the mother centriole for cartwheel assembly. Dev. Cell 2013, 25, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Dzhindzhev, N.S.; Tzolovsky, G.; Lipinszki, Z.; Schneider, S.; Lattao, R.; Fu, J.; Debski, J.; Dadlez, M.; Glover, D.M. Plk4 phosphorylates Ana2 to trigger Sas6 recruitment and procentriole formation. Curr. Biol. 2014, 24, 2526–2532. [Google Scholar] [CrossRef] [PubMed]
- Ohta, M.; Ashikawa, T.; Nozaki, Y.; Kozuka-Hata, H.; Goto, H.; Inagaki, M.; Oyama, M.; Kitagawa, D. Direct interaction of Plk4 with STIL ensures formation of a single procentriole per parental centriole. Nat. Commun. 2014, 5, 5267. [Google Scholar] [CrossRef] [PubMed]
- Kratz, A.S.; Barenz, F.; Richter, K.T.; Hoffmann, I. Plk4-dependent phosphorylation of STIL is required for centriole duplication. Biol. Open 2015, 4, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Moyer, T.C.; Clutario, K.M.; Lambrus, B.G.; Daggubati, V.; Holland, A.J. Binding of STIL to Plk4 activates kinase activity to promote centriole assembly. J. Cell Biol. 2015, 209, 863–878. [Google Scholar] [CrossRef] [PubMed]
- McLamarrah, T.A.; Buster, D.W.; Galletta, B.J.; Boese, C.J.; Ryniawec, J.M.; Hollingsworth, N.A.; Byrnes, A.E.; Brownlee, C.W.; Slep, K.C.; Rusan, N.M.; et al. An ordered pattern of Ana2 phosphorylation by Plk4 is required for centriole assembly. J. Cell Biol. 2018, 217, 1217–1231. [Google Scholar] [CrossRef] [PubMed]
- Ohta, M.; Watanabe, K.; Ashikawa, T.; Nozaki, Y.; Yoshiba, S.; Kimura, A.; Kitagawa, D. Bimodal binding of STIL to Plk4 controls proper centriole copy number. Cell Rep. 2018, 23, 3160–3169. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, M.; Müller-Reichert, T.; Oegema, K.; Grill, S.; Hyman, A.A. SAS-4 is a C. elegans centriolar protein that controls centrosome size. Cell 2003, 112, 575–587. [Google Scholar] [CrossRef]
- Leidel, S.; Gonczy, P. SAS-4 is essential for centrosome duplication in C. elegans and is recruited to daughter centrioles once per cell cycle. Dev. Cell 2003, 4, 431–439. [Google Scholar] [CrossRef]
- Basto, R.; Lau, J.; Vinogradova, T.; Gardiol, A.; Woods, C.G.; Khodjakov, A.; Raff, J.W. Flies without centrioles. Cell 2006, 125, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, J.; Mennella, V.; Blachon, S.; Zhai, B.; Smith, A.H.; Megraw, T.L.; Nicastro, D.; Gygi, S.P.; Agard, D.A.; Avidor-Reiss, T. Sas-4 provides a scaffold for cytoplasmic complexes and tethers them in a centrosome. Nat. Commun. 2011, 2, 359. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Gooi, L.M.; Wason, A.; Gabriel, E.; Mehrjardi, N.Z.; Yang, Q.; Zhang, X.; Debec, A.; Basiri, M.L.; Avidor-Reiss, T.; et al. Conserved TCP domain of Sas-4/CPAP is essential for pericentriolar material tethering during centrosome biogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, E354–363. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Jin, C.; Chu, Y.; Varghese, C.; Hua, S.; Yan, F.; Miao, Y.; Liu, J.; Mann, D.; Ding, X.; et al. Dimerization of CPAP orchestrates centrosome cohesion plasticity. J. Biol. Chem. 2010, 285, 2488–2497. [Google Scholar] [CrossRef] [PubMed]
- Cottee, M.A.; Muschalik, N.; Wong, Y.L.; Johnson, C.M.; Johnson, S.; Andreeva, A.; Oegema, K.; Lea, S.M.; Raff, J.W.; van Breugel, M. Crystal structures of the CPAP/STIL complex reveal its role in centriole assembly and human microcephaly. Elife 2013, 2, e01071. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.Y.; Chen, H.L.; Chang, C.W.; Li, B.R.; Tang, T.K. Identification of a novel microtubule-destabilizing motif in CPAP that binds to tubulin heterodimers and inhibits microtubule assembly. Mol. Biol. Cell 2004, 15, 2697–2706. [Google Scholar] [CrossRef] [PubMed]
- Cormier, A.; Clement, M.J.; Knossow, M.; Lachkar, S.; Savarin, P.; Toma, F.; Sobel, A.; Gigant, B.; Curmi, P.A. The PN2-3 domain of centrosomal P4.1-associated protein implements a novel mechanism for tubulin sequestration. J. Biol. Chem. 2009, 284, 6909–6917. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, S.; Rogala, K.B.; Dynes, N.J.; Hilbert, M.; Leidel, S.A.; Steinmetz, M.O.; Gonczy, P.; Vakonakis, I. Interaction between the Caenorhabditis elegans centriolar protein SAS-5 and microtubules facilitates organelle assembly. Mol. Biol. Cell 2018, 29, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Hatzopoulos, G.N.; Erat, M.C.; Cutts, E.; Rogala, K.B.; Slater, L.M.; Stansfeld, P.J.; Vakonakis, I. Structural analysis of the G-box domain of the microcephaly protein CPAP suggests a role in centriole architecture. Structure 2013, 21, 2069–2077. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Farmer, V.; Shukla, A.; James, J.; Gruskin, R.; Kiriyama, S.; Loncarek, J. Centriole maturation requires regulated Plk1 activity during two consecutive cell cycles. J. Cell Biol. 2014, 206, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Novak, Z.A.; Conduit, P.T.; Wainman, A.; Raff, J.W. Asterless licenses daughter centrioles to duplicate for the first time in Drosophila embryos. Curr. Biol. 2014, 24, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Loncarek, J.; Hergert, P.; Magidson, V.; Khodjakov, A. Control of daughter centriole formation by the pericentriolar material. Nat. Cell Biol. 2008, 10, 322–328. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, E.; Greenan, G.; Lange, K.I.; Srayko, M.; Müller-Reichert, T. The role of γ-tubulin in centrosomal microtubule organization. PLoS ONE 2012, 7, e29795. [Google Scholar] [CrossRef] [PubMed]
- Peel, N.; Stevens, N.R.; Basto, R.; Raff, J.W. Overexpressing centriole-replication proteins in vivo induces centriole overduplication and de novo formation. Curr. Biol. 2007, 17, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, D.; Fluckiger, I.; Polanowska, J.; Keller, D.; Reboul, J.; Gonczy, P. PP2a phosphatase acts upon SAS-5 to ensure centriole formation in C. elegans embryos. Dev. Cell 2011, 20, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Ferreira, I.; Rodrigues-Martins, A.; Bento, I.; Riparbelli, M.; Zhang, W.; Laue, E.; Callaini, G.; Glover, D.M.; Bettencourt-Dias, M. The SCF/Slimb ubiquitin ligase limits centrosome amplification through degradation of SAK/PLK4. Curr. Biol. 2009, 19, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.C.; Rusan, N.M.; Roberts, D.M.; Peifer, M.; Rogers, S.L. The SCF slimb ubiquitin ligase regulates Plk4/Sak levels to block centriole reduplication. J. Cell Biol. 2009, 184, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Holland, A.J.; Lan, W.; Niessen, S.; Hoover, H.; Cleveland, D.W. Polo-like kinase 4 kinase activity limits centrosome overduplication by autoregulating its own stability. J. Cell Biol. 2010, 188, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, C.W.; Klebba, J.E.; Buster, D.W.; Rogers, G.C. The protein phosphatase 2A regulatory subunit twins stabilizes Plk4 to induce centriole amplification. J. Cell Biol. 2011, 195, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Ferreira, I.; Bento, I.; Pimenta-Marques, A.; Jana, S.C.; Lince-Faria, M.; Duarte, P.; Borrego-Pinto, J.; Gilberto, S.; Amado, T.; Brito, D.; et al. Regulation of autophosphorylation controls PLK4 self-destruction and centriole number. Curr. Biol. 2013, 23, 2245–2254. [Google Scholar] [CrossRef] [PubMed]
- Klebba, J.E.; Buster, D.W.; Nguyen, A.L.; Swatkoski, S.; Gucek, M.; Rusan, N.M.; Rogers, G.C. Polo-like kinase 4 autodestructs by generating its Slimb-binding phosphodegron. Curr. Biol. 2013, 23, 2255–2261. [Google Scholar] [CrossRef] [PubMed]
- Arquint, C.; Cubizolles, F.; Morand, A.; Schmidt, A.; Nigg, E.A. The SKP1-Cullin-F-box E3 ligase βTrCP and CDK2 cooperate to control STIL abundance and centriole number. Open Biol. 2018, 8, 170253. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, J.B.; Wueseke, O.; Viscardi, V.; Mahamid, J.; Ochoa, S.D.; Bunkenborg, J.; Widlund, P.O.; Pozniakovsky, A.; Zanin, E.; Bahmanyar, S.; et al. Centrosomes. Regulated assembly of a supramolecular centrosome scaffold in vitro. Science 2015, 348, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Zwicker, D.; Decker, M.; Jaensch, S.; Hyman, A.A.; Julicher, F. Centrosomes are autocatalytic droplets of pericentriolar material organized by centrioles. Proc. Natl. Acad. Sci. USA 2014, 111, E2636–E2645. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, J.B.; Ferreira Gomes, B.; Widlund, P.O.; Mahamid, J.; Honigmann, A.; Hyman, A.A. The centrosome is a selective condensate that nucleates microtubules by concentrating tubulin. Cell 2017, 169, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Mahamid, J.; Pfeffer, S.; Schaffer, M.; Villa, E.; Danev, R.; Cuellar, L.K.; Forster, F.; Hyman, A.A.; Plitzko, J.M.; Baumeister, W. Visualizing the molecular sociology at the HeLa cell nuclear periphery. Science 2016, 351, 969–972. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarz, A.; Sankaralingam, P.; O’Connell, K.F.; Müller-Reichert, T. Revisiting Centrioles in Nematodes—Historic Findings and Current Topics. Cells 2018, 7, 101. https://doi.org/10.3390/cells7080101
Schwarz A, Sankaralingam P, O’Connell KF, Müller-Reichert T. Revisiting Centrioles in Nematodes—Historic Findings and Current Topics. Cells. 2018; 7(8):101. https://doi.org/10.3390/cells7080101
Chicago/Turabian StyleSchwarz, Anna, Prabhu Sankaralingam, Kevin F. O’Connell, and Thomas Müller-Reichert. 2018. "Revisiting Centrioles in Nematodes—Historic Findings and Current Topics" Cells 7, no. 8: 101. https://doi.org/10.3390/cells7080101
APA StyleSchwarz, A., Sankaralingam, P., O’Connell, K. F., & Müller-Reichert, T. (2018). Revisiting Centrioles in Nematodes—Historic Findings and Current Topics. Cells, 7(8), 101. https://doi.org/10.3390/cells7080101