Lack of LTβR Increases Susceptibility of IPEC-J2 Cells to Porcine Epidemic Diarrhea Virus



Abstract
1. Introduction
2. Materials and Methods
2.1. Porcine Intestine Samples
2.2. Cell Culture
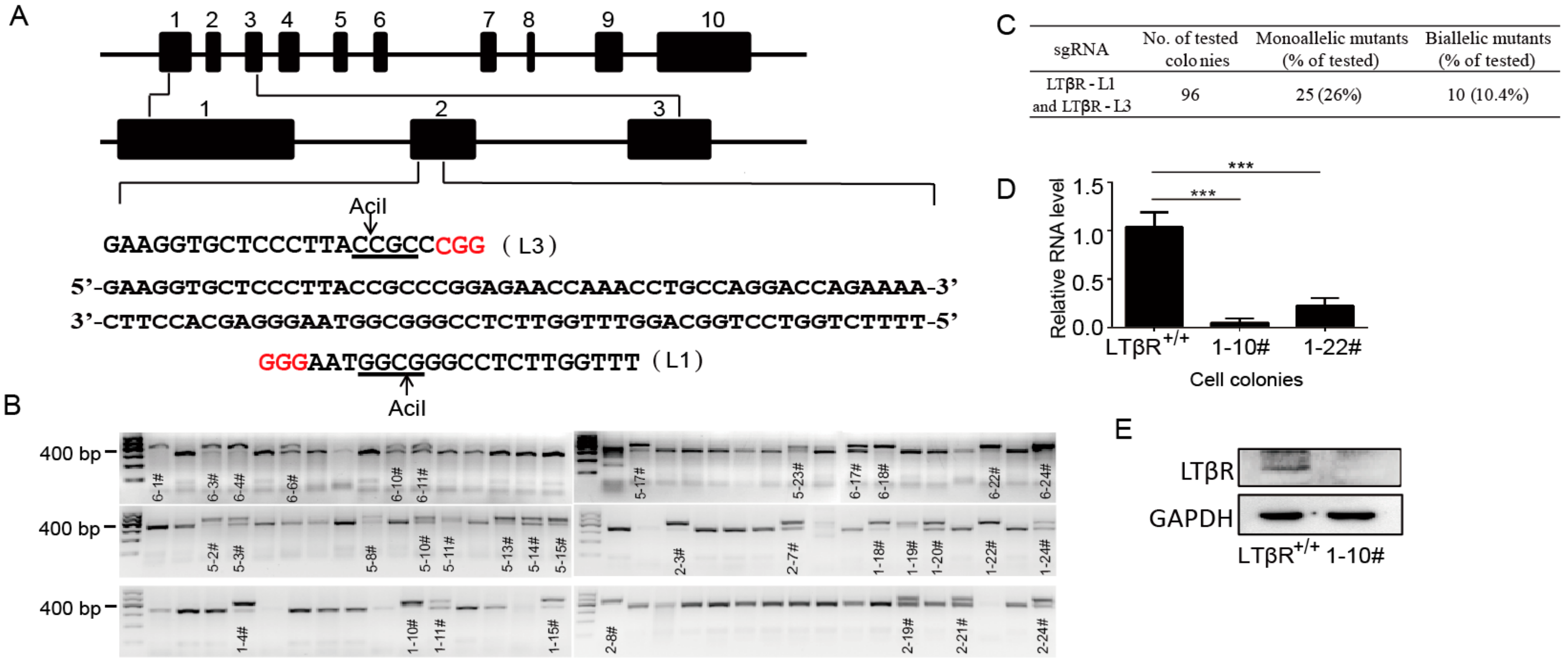
2.3. Gene Targeting by the CRISPR/CAS9 System
2.4. Reverse Transcription PCR (RT-PCR)
2.5. Western Blotting
2.6. Cell Proliferation
2.7. Cell Cycle Analysis
2.8. Digital HoloMonitor Microscopy
2.9. Statistical Analysis
3. Results
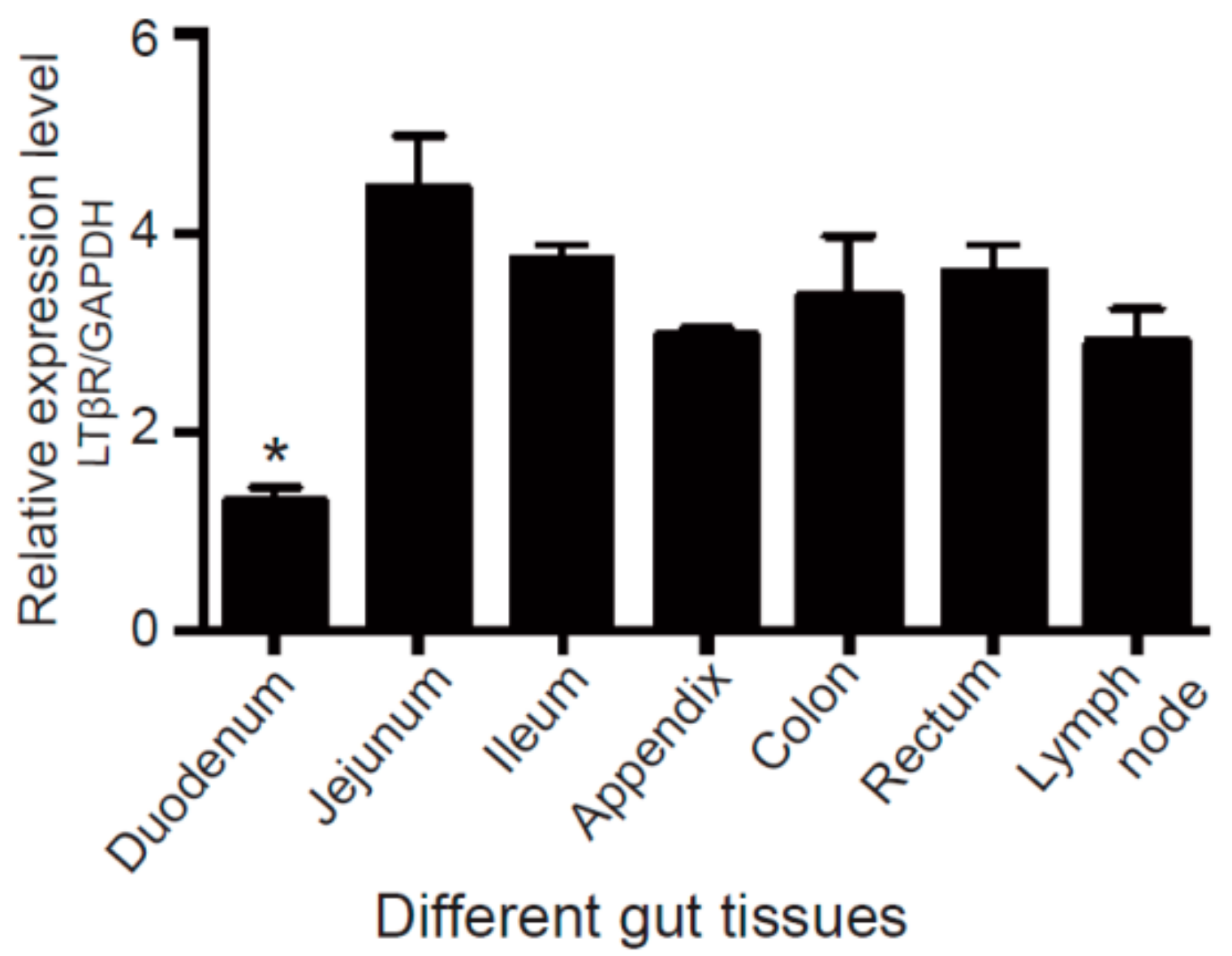
3.1. LTβR Is Highly Expressed in Porcine Gut Tissues
3.2. Generation of LTβR−/− Cells Using CRISPR/Cas9
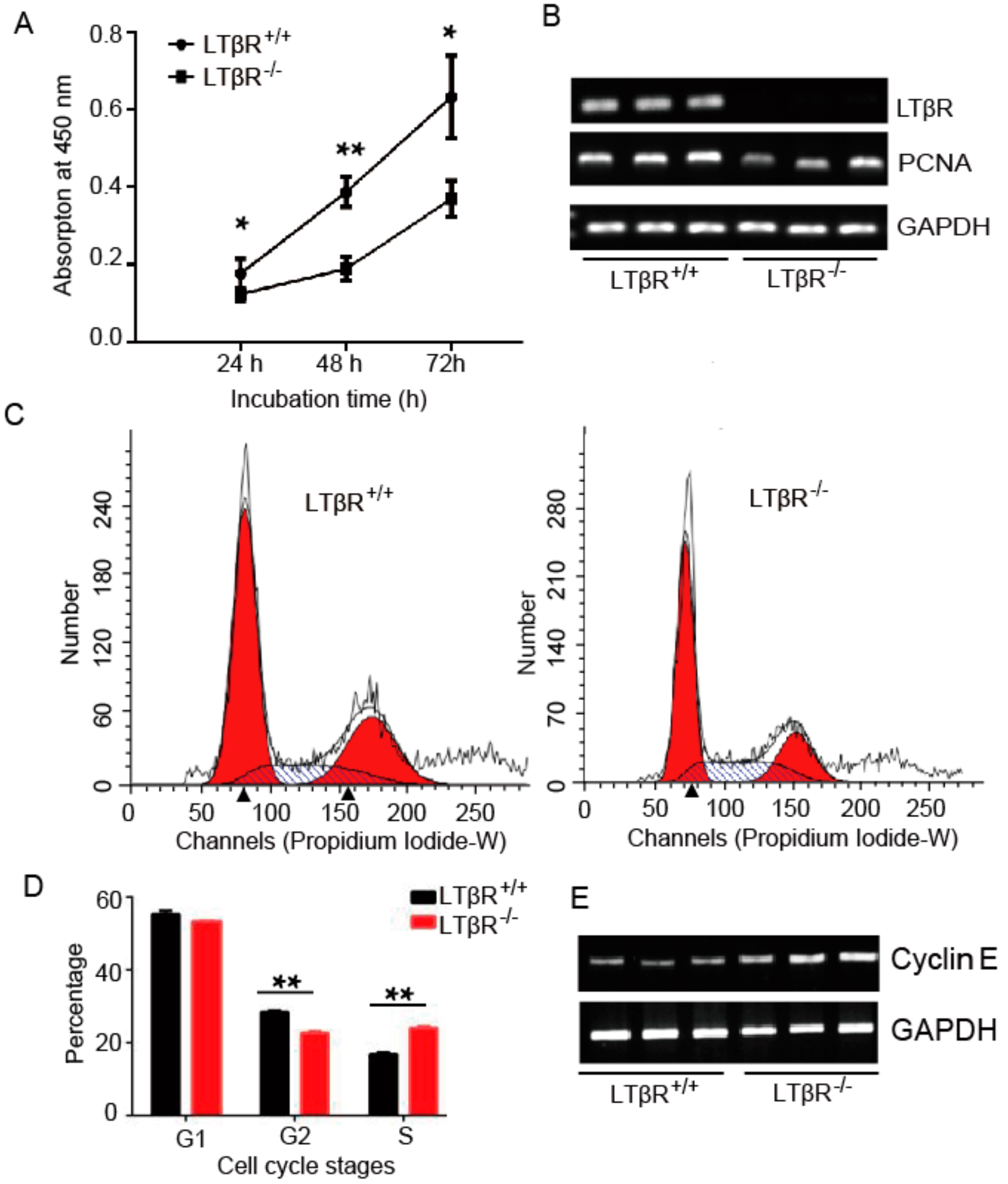
3.3. LTβR Knockout Inhibits IPEC-J2 Cell Proliferation
3.4. LTβR Knockout Induces IPEC-J2 Cell Apoptosis
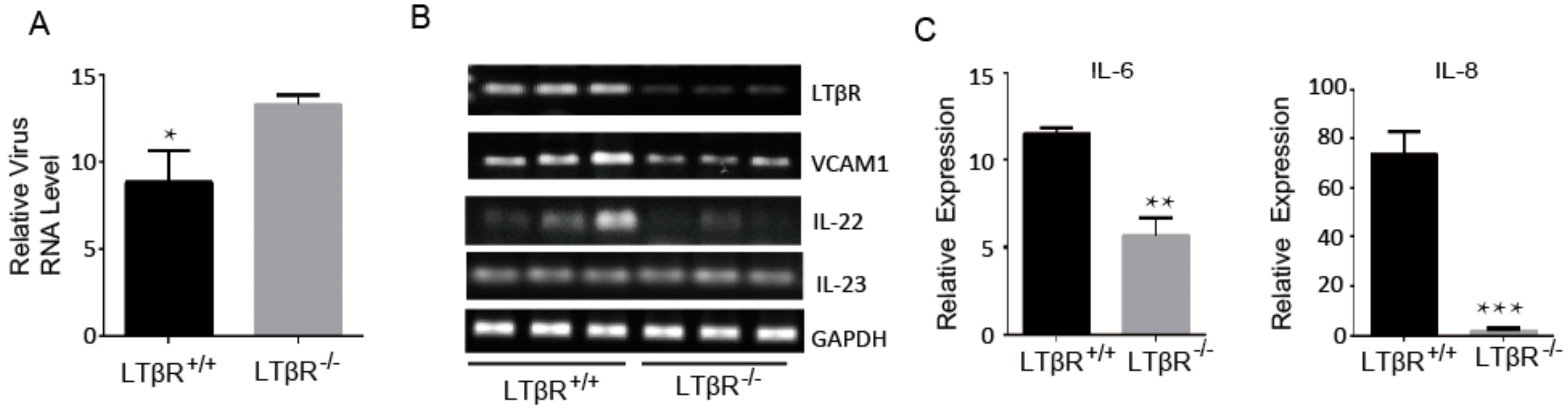
3.5. LTβR Knockout IPEC-J2 Cells Are Susceptible to PEDV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Force, W.R.; Walter, B.N.; Hession, C.; Tizard, R.; Kozak, C.A.; Browning, J.L.; Ware, C.F. Mouse lymphotoxin-beta receptor. Molecular genetics, ligand binding, and expression. J. Immunol. 1995, 155, 5280–5288. [Google Scholar] [PubMed]
- Browning, J.L.; French, L.E. Visualization of lymphotoxin-beta and lymphotoxin-beta receptor expression in mouse embryos. J. Immunol. 2002, 168, 5079–5087. [Google Scholar] [CrossRef] [PubMed]
- Futterer, A.; Mink, K.; Luz, A.; Kosco-Vilbois, M.H.; Pfeffer, K. The lymphotoxin beta receptor controls organogenesis and affinity maturation in peripheral lymphoid tissues. Immunity 1998, 9, 59–70. [Google Scholar] [CrossRef]
- Hehlgans, T.; Muller, P.; Stopfer, P.; Mannel, D.N. Activation of the lymphotoxin-beta receptor induces NFkappaB-dependent interleukin-6 and MIP-2 secretion in mouse fibrosarcoma cells. Eur. Cytokine Netw. 2003, 14, 103–107. [Google Scholar] [PubMed]
- Ehlers, S.; Hölscher, C.; Scheu, S.; Tertilt, C.; Hehlgans, T.; Suwinski, J.; Endres, R.; Pfeffer, K. The lymphotoxin beta receptor is critically involved in controlling infections with the intracellular pathogens Mycobacterium tuberculosis and Listeria monocytogenes. J. Immunol. 2003, 170, 5210–5218. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, V.; Poroyko, V.; Kim, T.-J.; Devkota, S.; Fu, S.; Liu, D.; Tumanov, A.V.; Koroleva, E.P.; Deng, L.; Nagler, C.; et al. Lymphotoxin regulates commensal responses to enable diet-induced obesity. Nat. Immunol. 2012, 13, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Scarzello, A.J.; Jiang, Q.; Back, T.; Dang, H.; Hodge, D.; Hanson, C.; Subleski, J.; Weiss, J.M.; Stauffer, J.K.; Chaisaingmongkol, J.; et al. LTbetaR signalling preferentially accelerates oncogenic AKT-initiated liver tumours. Gut 2015, 65, 1765–1775. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Li, N.; Li, F.; Sang, J.; Deng, H.; Han, Q.; Lv, Y.; Li, C.; Liu, Z. Association of LTBR polymorphisms with chronic hepatitis B virus infection and hepatitis B virus-related hepatocellular carcinoma. Int. Immunopharmacol. 2017, 49, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Onder, L.; Danuser, R.; Scandella, E.; Firner, S.; Chai, Q.; Hehlgans, T.; Stein, J.V.; Ludewig, B. Endothelial cell-specific lymphotoxin-beta receptor signaling is critical for lymph node and high endothelial venule formation. J. Exp. Med. 2013, 210, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Macho-Fernandez, E.; Koroleva, E.P.; Spencer, C.M.; Tighe, M.; Torrado, E.; Cooper, A.M.; Fu, Y.-X.; Tumanov, A.V. Lymphotoxin beta receptor signaling limits mucosal damage through driving IL-23 production by epithelial cells. Mucosal Immunol. 2015, 8, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, N.; Huber, B.; Barabas, N.; Röhrl, J.; Pfeffer, K.; Hehlgans, T. Lymphotoxin beta receptor activation on macrophages induces cross-tolerance to TLR4 and TLR9 ligands. J. Immunol. 2012, 188, 3426–3433. [Google Scholar] [CrossRef] [PubMed]
- Debouck, P.; Pensaert, M. Experimental infection of pigs with a new porcine enteric coronavirus, CV 777. Am. J. Vet. Res. 1980, 41, 219–223. [Google Scholar] [PubMed]
- Sun, R.Q.; Cai, R.J.; Chen, Y.Q.; Liang, P.S.; Chen, D.K.; Song, C.X. Outbreak of porcine epidemic diarrhea in suckling piglets, China. Emerg. Infect. Dis. 2012, 18, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Cima, G. Viral disease affects U.S. pigs: Porcine epidemic diarrhea found in at least 11 states. J. Am. Vet. Med. Assoc. 2013, 243, 30–31. [Google Scholar] [PubMed]
- Wang, Y.; Koroleva, E.P.; Kruglov, A.A.; Kuprash, D.V.; Nedospasov, S.A.; Fu, Y.X.; Tumanov, A.V. Lymphotoxin beta receptor signaling in intestinal epithelial cells orchestrates innate immune responses against mucosal bacterial infection. Immunity 2010, 32, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Shi, H.; Guo, D.; Chen, J.; Shi, D.; Zhu, Q.; Zhang, X.; Feng, L. Analysis of protein expression changes of the Vero E6 cells infected with classic PEDV strain CV777 by using quantitative proteomic technique. J. Virol. Methods 2015, 218, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, J.; Cao, C.; Huang, J.; Hai, T.; Wang, Y.; Zheng, Q.; Zhang, H.; Qin, G.; Miao, X.; et al. Efficient CRISPR/Cas9-mediated biallelic gene disruption and site-specific knockin after rapid selection of highly active sgRNAs in pigs. Sci. Rep. 2015, 5, 13348. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sengel, C.; Emerson, M.M.; Cepko, C.L. A gene regulatory network controls the binary fate decision of rod and bipolar cells in the vertebrate retina. Dev. Cell 2014, 30, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.; Sicinski, P.; Hinds, P.W. Cyclins and cdks in development and cancer: A perspective. Oncogene 2005, 24, 2909–2915. [Google Scholar] [CrossRef] [PubMed]
- Mir, T.A.; Shinohara, H. Label-free observation of three-dimensional morphology change of a single PC12 cell by digital holographic microscopy. Anal. Biochem. 2012, 429, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Xia, R.W.; Sun, L.; Qin, W.Y.; Gan, L.N.; Bao, W.B.; Wu, S.L. Developmental expression of LTbetaR and differential expression in Escherichia coli F18 resistant/sensitive piglets. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Gao, J.; Zhu, L.; Yang, Q. Transmissible gastroenteritis virus and porcine epidemic diarrhoea virus infection induces dramatic changes in the tight junctions and microfilaments of polarized IPEC-J2 cells. Virus Res. 2014, 192, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Hsieh, S.L.; Chen, M.C.; Lin, W.W. Lymphotoxin beta receptor induces interleukin 8 gene expression via NF-kappaB and AP-1 activation. Exp. Cell Res. 2002, 278, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.K.; Noh, E.-K.; Gwon, G.-D.; Kim, J.Y.; Jo, J.-C.; Choi, Y.; Koh, S.; Baek, J.H.; Min, Y.J.; Kim, H. LIGHT (TNFSF14) Increases the Survival and Proliferation of Human Bone Marrow-Derived Mesenchymal Stem Cells. PLoS ONE 2016, 11, e0166589. [Google Scholar] [CrossRef] [PubMed]
- Rooney, I.A.; Butrovich, K.D.; Glass, A.A.; Borboroglu, S.; Benedict, C.A.; Whitbeck, J.C.; Cohen, G.H.; Eisenberg, R.J.; Ware, C.F. The lymphotoxin-beta receptor is necessary and sufficient for LIGHT-mediated apoptosis of tumor cells. J. Biol. Chem. 2000, 275, 14307–14315. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.Y.; Wang, P.Y.; Han, S.H.; Hsieh, S.L. The cytoplasmic domain of the lymphotoxin-beta receptor mediates cell death in HeLa cells. J. Biol. Chem. 1999, 274, 11868–11873. [Google Scholar] [CrossRef] [PubMed]
- Dejardin, E.; Droin, N.M.; Delhase, M.; Haas, E.; Cao, Y.; Makris, C.; Li, Z.-W.; Karin, M.; Ware, C.F.; Green, D.R. The lymphotoxin-beta receptor induces different patterns of gene expression via two NF-kappaB pathways. Immunity 2002, 17, 525–535. [Google Scholar] [CrossRef]
- Wroblewska, J.A.; Zhang, Y.; Tang, H.; Guo, X.; Nagler, C.; Fu, Y.X. Cutting Edge: Lymphotoxin Signaling Is Essential for Clearance of Salmonella from the Gut Lumen and Generation of Anti-Salmonella Protective Immunity. J. Immunol. 2017, 198, 55–60. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| Lymphotoxin beta receptor (LTβR) | CACTCATGCTGGGCCTCT | GAGCAGCAGACGTGATGTTT |
| Vascular cell adhesion molecule 1 (VCAM1) | ATCCAAGCTGCTCCAAAAGA | GGCCCTGTGGATGGTATATG |
| Interleukin-22 (IL-22) | TTGCTCAAGTTCGTGTCGTC | GGTCAAGCTTGCAGTGATGA |
| Interleukin-23 (IL-23) | TAGGGGTCGAGTCAGAGGTG | GAGTGCCATCCTTGAGCTGT |
| Interleukin-6 (IL-6) | CCACCGGTCTTGTGGAGTTT | AGTCGGGTTGTCTAGGCTGA |
| Interleukin-8 (IL-8) | TGCAAGCTTTGTTATGCAGTG | GCCTGGTGAATTTTTGCTGT |
| Proliferating cell nuclear antigen (PCNA) | GATTCCACCACCATGTTCGAG | GATTCCACCACCATGTTCGAG |
| Caspase 3 (CASP3) | GCCATGGTGAAGAAGGAAAA | GTCCGTCTCAATCCCACAGT |
| Tumor necrosis factor Superfamily member 10 (TNFSF10) | ACCCAAAGGCTCAACAC | CCCACCTGAGATGGATCACT |
| Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) | GTGAAGGTCGGAGTGAACG | CTCGCTCCTGGAAGATGGTG |
| Porcine epidemic diarrhea virus (PEDV) | GCACTTATTGGCAGGCTTTGT | CCATTGAGAAAAGAAAGTGTCGTAG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altawaty, T.; Liu, L.; Zhang, H.; Tao, C.; Hou, S.; Li, K.; Wang, Y. Lack of LTβR Increases Susceptibility of IPEC-J2 Cells to Porcine Epidemic Diarrhea Virus. Cells 2018, 7, 222. https://doi.org/10.3390/cells7110222
Altawaty T, Liu L, Zhang H, Tao C, Hou S, Li K, Wang Y. Lack of LTβR Increases Susceptibility of IPEC-J2 Cells to Porcine Epidemic Diarrhea Virus. Cells. 2018; 7(11):222. https://doi.org/10.3390/cells7110222
Chicago/Turabian StyleAltawaty, Tawfeek, Lulu Liu, Hongyong Zhang, Cong Tao, Shaohua Hou, Kui Li, and Yanfang Wang. 2018. "Lack of LTβR Increases Susceptibility of IPEC-J2 Cells to Porcine Epidemic Diarrhea Virus" Cells 7, no. 11: 222. https://doi.org/10.3390/cells7110222
APA StyleAltawaty, T., Liu, L., Zhang, H., Tao, C., Hou, S., Li, K., & Wang, Y. (2018). Lack of LTβR Increases Susceptibility of IPEC-J2 Cells to Porcine Epidemic Diarrhea Virus. Cells, 7(11), 222. https://doi.org/10.3390/cells7110222