Assessing Autophagy in Sciatic Nerves of a Rat Model that Develops Inflammatory Autoimmune Peripheral Neuropathies

 ,
, 
Abstract
1. Introduction
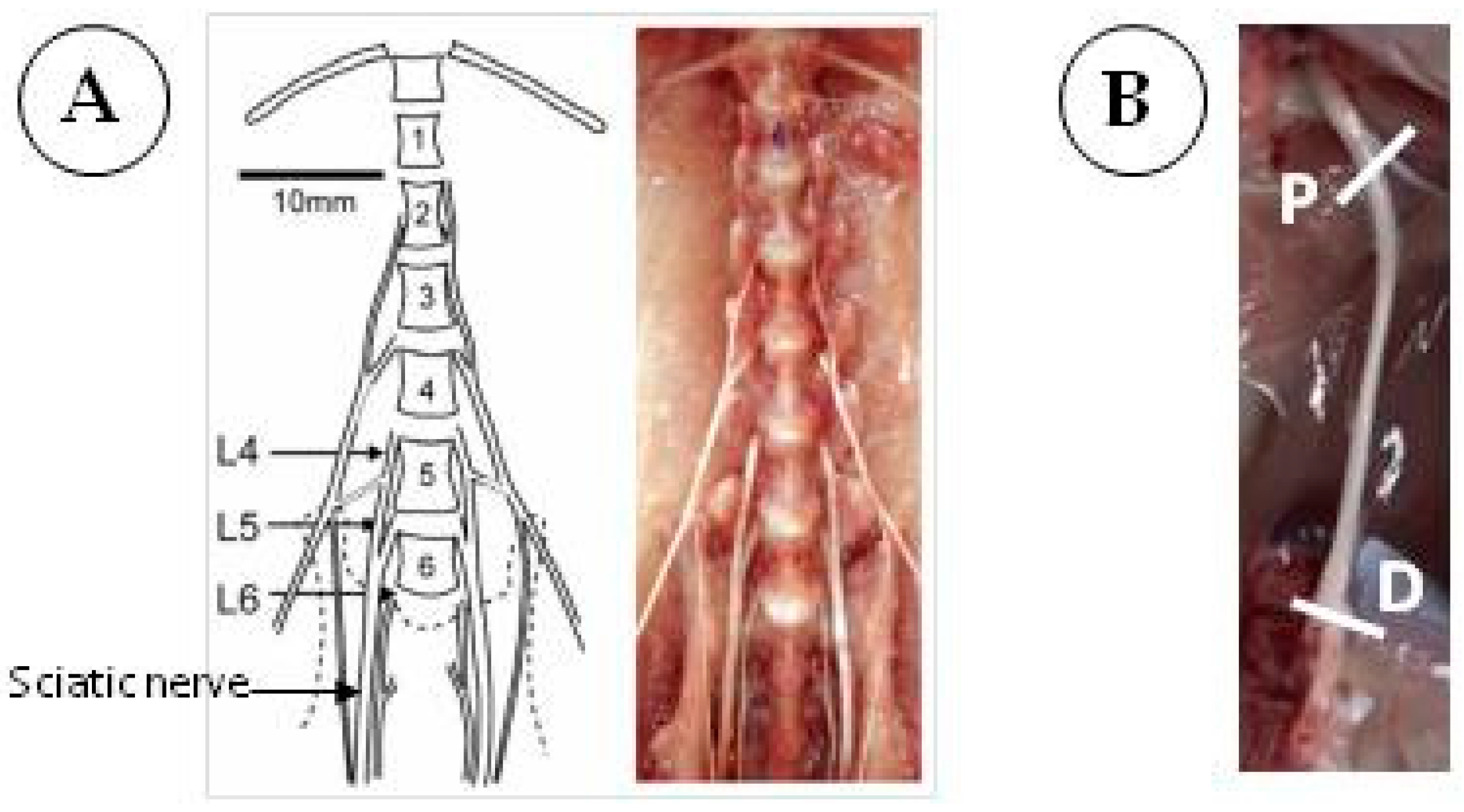
2. Rat Sciatic Nerve as a Valuable Model to Study Autophagy
2.1. Induction of CIDP in Rats
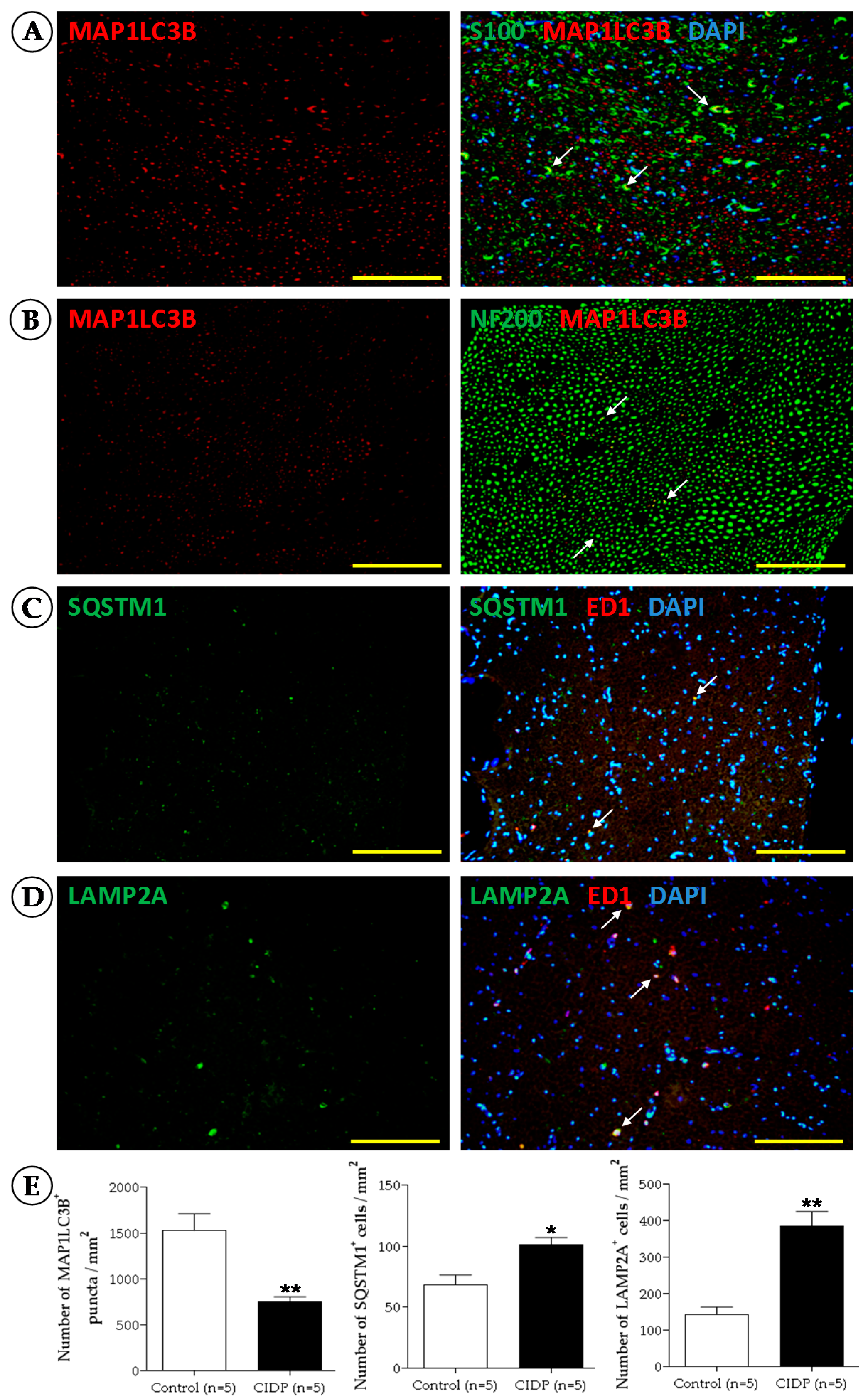
2.2. Immunofluorescence
2.2.1. Procedure
2.2.2. Notes
- (1)
- to use normal horse serum or normal donkey serum as blocking agent in PBS-Tween (PBS-T) rather than bovine serum albumin or fetal bovine serum;
- (2)
- to use the primary antibodies in blocking solution;
- (3)
- to use the secondary antibody in PBS-T and not in blocking solution;
- (4)
- to wash in PBS and not in PBS-T.
2.2.3. Advantages
2.2.4. Limitations
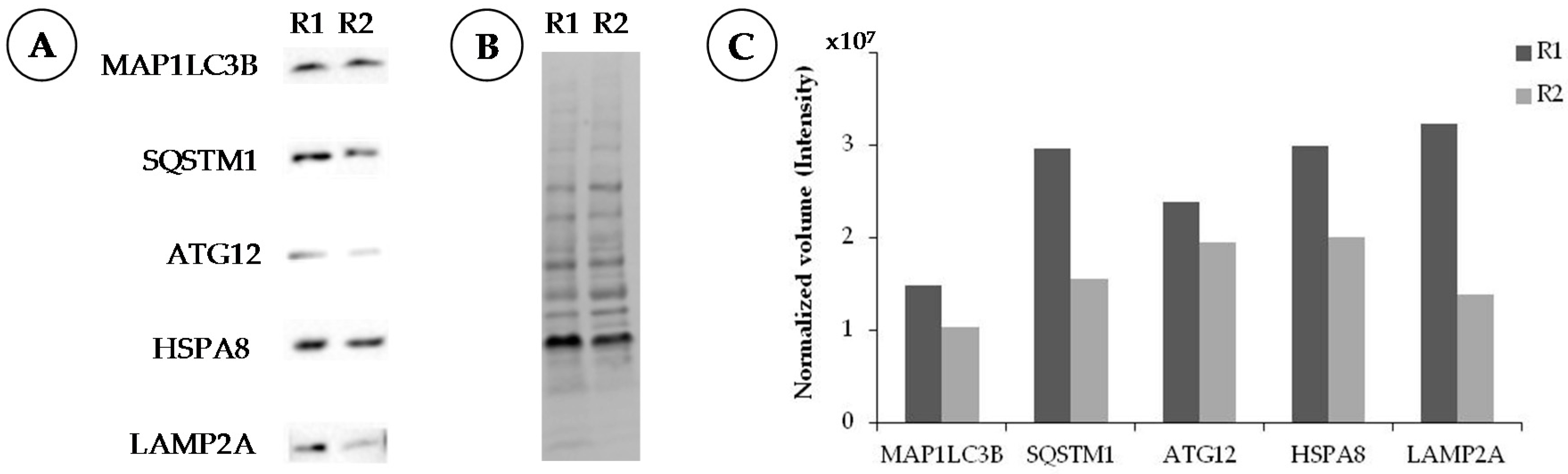
2.3. Western Blotting
2.3.1. Procedure
2.3.2. Notes
2.3.3. Advantages and Limitations
- sensitivity (detection of small protein amounts);
- quantification of the amount of protein.
- high cost, time, and technical demand;
- requirement of numerous experimental optimizations (protein extraction, lysis buffer, blocking buffer, etc.);
- unexpected results: no bands, weak signal, unexpected or unusual bands, high background, etc.
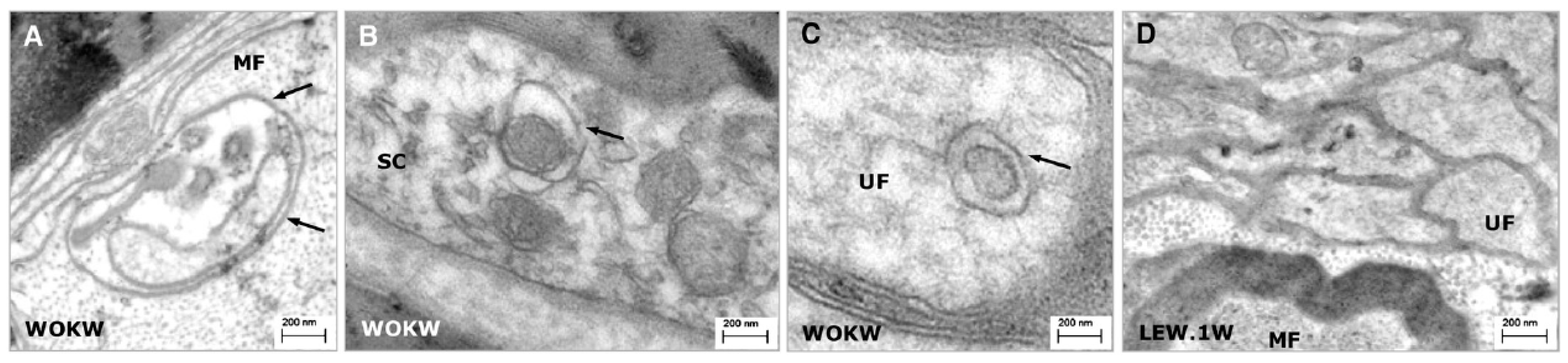
2.4. TEM
2.4.1. Procedure
2.4.2. Note
2.4.3. Advantages and Limitations
3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Ethics Statement
References
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Muller, S. Manipulating autophagic processes in autoimmune diseases: A special focus on modulating chaperone-mediated autophagy, an emerging therapeutic target. Front. Immunol. 2015, 6, 252. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Klionsky, D.J. Eaten alive: A history of macroautophagy. Nat. Cell Biol. 2010, 12, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Weidberg, H.; Shvets, E.; Elazar, Z. Biogenesis and cargo selectivity of autophagosomes. Annu. Rev. Biochem. 2011, 80, 125–156. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, S.; Cuervo, A.M. Chaperone-mediated autophagy: A unique way to enter the lysosome world. Trends Cell Biol. 2012, 22, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, A.M.; Wong, E. Chaperone-mediated autophagy: Roles in disease and aging. Cell Res. 2014, 24, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A.; Kwon, Y.T. Degradation of misfolded proteins in neurodegenerative diseases: Therapeutic targets and strategies. Exp. Mol. Med. 2015, 47, e147. [Google Scholar] [CrossRef] [PubMed]
- Ahlberg, J.; Glaumann, H. Uptake–microautophagy–and degradation of exogenous proteins by isolated rat liver lysosomes. Effects of pH, ATP, and inhibitors of proteolysis. Exp. Mol. Pathol. 1985, 42, 78–88. [Google Scholar] [CrossRef]
- Mijaljica, D.; Prescott, M.; Devenish, R.J. Microautophagy in mammalian cells: Revisiting a 40-year-old conundrum. Autophagy 2011, 7, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.J.; Xavier, R.J. Autophagy, immunity and human disease. Curr. Opin. Gastroenterol. 2009, 25, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Nagata, E.; Sawa, A.; Ross, C.A.; Snyder, S.H. Autophagosome-like vacuole formation in Huntington’s disease lymphoblasts. Neuroreport 2004, 15, 1325–1328. [Google Scholar] [CrossRef] [PubMed]
- Gros, F.; Arnold, J.; Page, N.; Décossas, M.; Korganow, A.-S.; Martin, T.; Muller, S. Macroautophagy is deregulated in murine and human lupus T lymphocytes. Autophagy 2012, 8, 1113–1123. [Google Scholar] [CrossRef] [PubMed]
- Gianchecchi, E.; Delfino, D.V.; Fierabracci, A. Recent insights on the putative role of autophagy in autoimmune diseases. Autoimmun. Rev. 2014, 13, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Macri, C.; Wang, F.; Tasset, I.; Schall, N.; Page, N.; Briand, J.-P.; Cuervo, A.M.; Muller, S. Modulation of deregulated chaperone-mediated autophagy by a phosphopeptide. Autophagy 2015, 11, 472–486. [Google Scholar] [CrossRef] [PubMed]
- Sasazawa, Y.; Sato, N.; Umezawa, K.; Simizu, S. Conophylline protects cells in cellular models of neurodegenerative diseases by inducing mammalian target of rapamycin (mTOR)-independent autophagy. J. Biol. Chem. 2015, 290, 6168–6178. [Google Scholar] [CrossRef] [PubMed]
- Mintern, J.D.; Harris, J. Autophagy and immunity. Immunol. Cell Biol. 2015, 93, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wang, H.; Wu, Y.; He, Z.; Qin, Y.; Shen, Q. The autophagy level is increased in the synovial tissues of patients with active rheumatoid arthritis and is correlated with disease severity. Mediators Inflamm. 2017, 2017, 7623145. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, B.; Schall, N.; Wilhelm, M.; Muller, S. Assessing autophagy in mouse models and patients with systemic autoimmune diseases. Cells 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Brun, S.; René, F.; de Sèze, J.; Loeffler, J.P.; Jeltsch-David, H. Autophagy in neuroinflammatory diseases. Autoimmun. Rev. 2017, 16, 856–874. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.A.C.; Allen, D.; Makowska, A.; Gregson, N.A. Pathogenesis of chronic inflammatory demyelinating polyradiculoneuropathy. J. Peripher. Nerv. Syst. 2006, 11, 30–46. [Google Scholar] [CrossRef] [PubMed]
- Vucic, S.; Kiernan, M.C.; Cornblath, D.R. Guillain–Barré syndrome: An update. J. Clin. Neurosci. 2009, 16, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Chen, X.; Xue, R.; Zhou, Q.; Hu, P.; Ouyang, X.; Dai, T.; Zhu, W.; Tian, S. Autophagy is involved in the pathogenesis of experimental autoimmune neuritis in rats. Neuroreport 2016, 27, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Alirezaei, M.; Fox, H.S.; Flynn, C.T.; Moore, C.S.; Hebb, A.L.O.; Frausto, R.F.; Bhan, V.; Kiosses, W.B.; Whitton, J.L.; Robertson, G.S.; et al. Elevated ATG5 expression in autoimmune demyelination and multiple sclerosis. Autophagy 2009, 5, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Igci, M.; Baysan, M.; Yigiter, R.; Ulasli, M.; Geyik, S.; Bayraktar, R.; Bozgeyik, İ.; Bozgeyik, E.; Bayram, A.; Cakmak, E.A. Gene expression profiles of autophagy-related genes in multiple sclerosis. Gene 2016, 588, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.-P.; Wang, H.-X.; Zhuang, J.-C.; Liu, Q.-B.; Zhao, G.-X.; Li, Z.-X.; Wu, Z.-Y. Variants of autophagy-related gene 5 are associated with neuromyelitis optica in the Southern Han Chinese population. Autoimmunity 2014, 47, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Beaino, W.; Trifilieff, E. Thiopalmitoylated peptides from the peripheral nervous system myelin p0 protein: Synthesis, characterization, and neuritogenic properties. Bioconjugate Chem. 2010, 21, 1439–1447. [Google Scholar] [CrossRef] [PubMed]
- Brun, S.; Beaino, W.; Kremer, L.; Taleb, O.; Mensah-Nyagan, A.G.; Lam, C.D.; Greer, J.M.; de Seze, J.; Trifilieff, E. Characterization of a new rat model for chronic inflammatory demyelinating polyneuropathies. J. Neuroimmunol. 2015, 278, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Schmalbruch, H. Fiber composition of the rat sciatic nerve. Anat. Rec. 1986, 215, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Shehab, S.A.; Atkinson, M.E.; Payne, J.N. The origins of the sciatic nerve and changes in neuropeptides after axotomy: A double labelling study using retrograde transport of true blue and vasoactive intestinal polypeptide immunohistochemistry. Brain Res. 1986, 376, 180–185. [Google Scholar] [CrossRef]
- Rigaud, M.; Gemes, G.; Barabas, M.-E.; Chernoff, D.I.; Abram, S.E.; Stucky, C.L.; Hogan, Q.H. Species and strain differences in rodent sciatic nerve anatomy: Implications for studies of neuropathic pain. Pain 2008, 136, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Asato, F.; Butler, M.; Blomberg, H.; Gordh, T. Variation in rat sciatic nerve anatomy: Implications for a rat model of neuropathic pain. J. Peripher. Nerv. Syst. 2000, 5, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in mammalian autophagy research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, E.; Galluzzi, L.; Maiuri, M.C.; Criollo, A.; Vitale, I.; Hangen, E.; Modjtahedi, N.; Kroemer, G. Methods for assessing autophagy and autophagic cell death. Methods Mol. Biol. 2008, 445, 29–76. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Arozena, A.A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Cuervo, A.M.; Ravikumar, B.; Sarkar, S.; Korolchuk, V.; Kaushik, S.; Klionsky, D.J. In search of an “autophagomometer”. Autophagy 2009, 5, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Yerra, V.G.; Kumar, A. Adenosine monophosphate-activated protein kinase abates hyperglycaemia-induced neuronal injury in experimental models of diabetic neuropathy: Effects on mitochondrial biogenesis, autophagy and neuroinflammation. Mol. Neurobiol. 2017, 54, 2301–2312. [Google Scholar] [CrossRef] [PubMed]
- Kosacka, J.; Nowicki, M.; Blüher, M.; Baum, P.; Stockinger, M.; Toyka, K.V.; Klöting, I.; Stumvoll, M.; Serke, H.; Bechmann, I.; et al. Increased autophagy in peripheral nerves may protect Wistar Ottawa Karlsburg W rats against neuropathy. Exp. Neurol. 2013, 250, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Zhang, H.; Gu, B.; Dai, W.; Wu, Q.; Sun, L.; Zhao, L.; Shi, Y.; Liang, X. Jinmaitong alleviates the diabetic peripheral neuropathy by inducing autophagy. Chin. J. Integr. Med. 2016, 22, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Chen, L.; Zhang, H.-X.; Li, S.-F.; Liu, P.; Zhao, T.-Y.; Li, C.-X. Autophagy promotes peripheral nerve regeneration and motor recovery following sciatic nerve crush injury in rats. J. Mol. Neurosci. 2016, 58, 416–423. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neurological Autoimmune Diseases | Autophagy Abnormalities | Methods | Model Systems or Patient Samples Tested | Ref. |
|---|---|---|---|---|
| Guillain–Barré syndrome (GBS) | Increased Beclin-1 and MAP1LC3B expression levels; | WB; TEM | EAN rat (sciatic nerves) | [22] |
| Increased MAP1LC3BII/I ratio; Decreased expression of SQSTM1; | ||||
| Increased formation of autophagosomes | ||||
| Chronic inflammatory demyelinating polyneuropathy (CIDP) | Increased Beclin-1 and MAP1LC3B-II expression levels; | TEM; WB | EAN rat (sciatic nerves) | [19] |
| Increased ratio MAP1LC3B-II/I; | ||||
| Decreased expression of SQSTM1 | ||||
| Multiple sclerosis (MS) | Increased mRNA and protein level of ATG5; | qPCR; WB | EAE mice (blood) and patient (blood and brain) | [23] |
| Decreased expression of ATG16L2 and ATG9A genes; | qPCR | Patient (blood) | [24] | |
| Increased expression of ULK1 gene | ||||
| Neuromyelitis optica (NMO) | Increased ATG5 variants | Mass array system | Patient (blood) | [25] |
| Antibody | Supplier; References | Technique (Concentration) |
|---|---|---|
| MAP1LC3B | MBL; M186-3 | IF (5 μg/mL) |
| WB (0.5 μg/mL) | ||
| SQSTM1 | Abcam; ab109012 | IF (5 μg/mL) |
| WB (0.5 μg/mL) | ||
| HSPA8 | Abcam; ab21052 | WB (0.5 μg/mL) |
| ATG12 | Abcam; ab155589 | WB (0.5 μg/mL) |
| LAMP2A | Abcam; ab125068 | IF (5 μg/mL) |
| WB (0.5 μg/mL) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brun, S.; Schall, N.; Jeltsch-David, H.; Sèze, J.d.; Muller, S. Assessing Autophagy in Sciatic Nerves of a Rat Model that Develops Inflammatory Autoimmune Peripheral Neuropathies. Cells 2017, 6, 30. https://doi.org/10.3390/cells6030030
Brun S, Schall N, Jeltsch-David H, Sèze Jd, Muller S. Assessing Autophagy in Sciatic Nerves of a Rat Model that Develops Inflammatory Autoimmune Peripheral Neuropathies. Cells. 2017; 6(3):30. https://doi.org/10.3390/cells6030030
Chicago/Turabian StyleBrun, Susana, Nicolas Schall, Hélène Jeltsch-David, Jérôme de Sèze, and Sylviane Muller. 2017. "Assessing Autophagy in Sciatic Nerves of a Rat Model that Develops Inflammatory Autoimmune Peripheral Neuropathies" Cells 6, no. 3: 30. https://doi.org/10.3390/cells6030030
APA StyleBrun, S., Schall, N., Jeltsch-David, H., Sèze, J. d., & Muller, S. (2017). Assessing Autophagy in Sciatic Nerves of a Rat Model that Develops Inflammatory Autoimmune Peripheral Neuropathies. Cells, 6(3), 30. https://doi.org/10.3390/cells6030030