
Cyclic GMP and Cilia Motility

{kind=link}
{kind=link}
Abstract
:1. Introduction
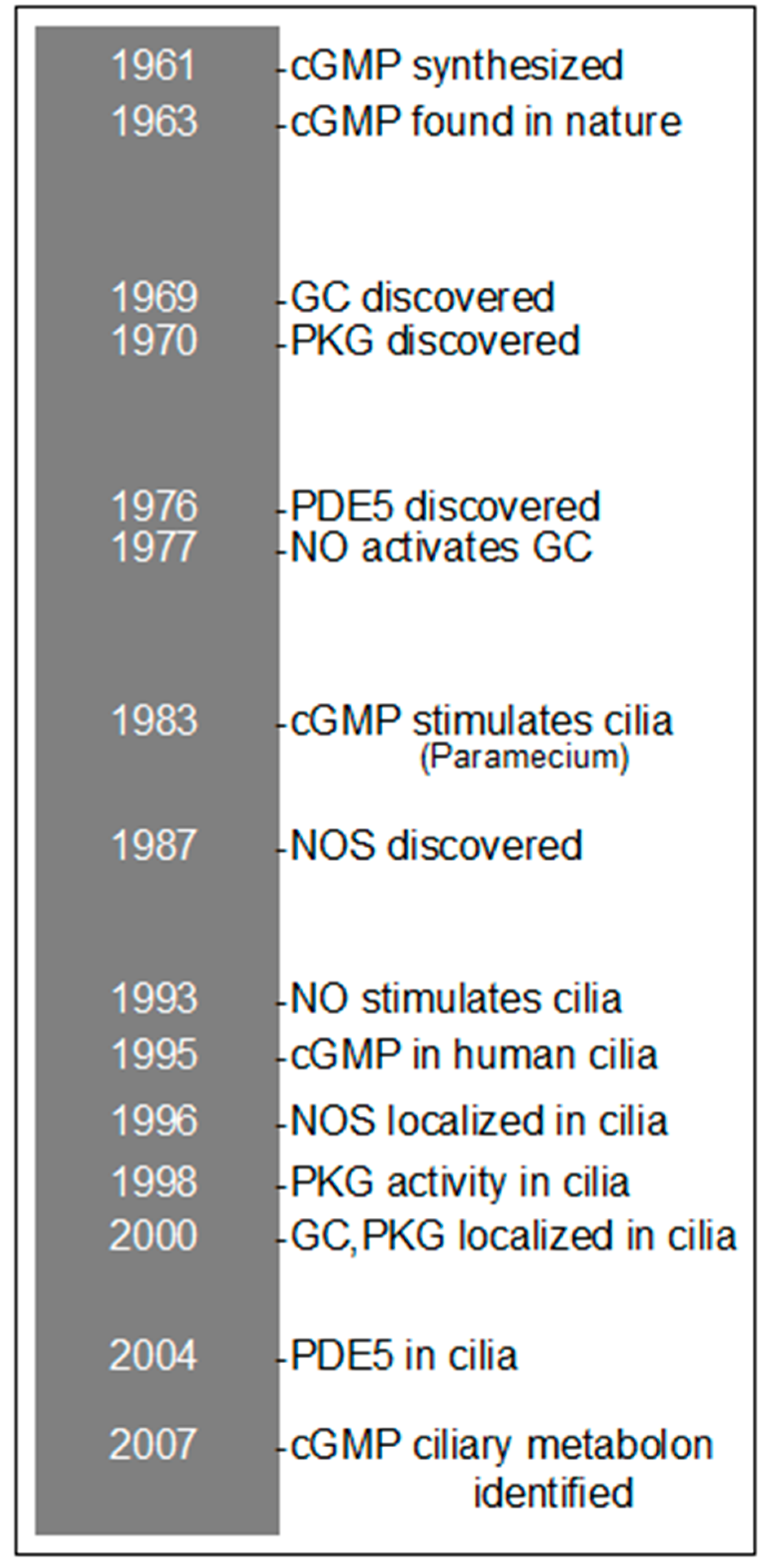
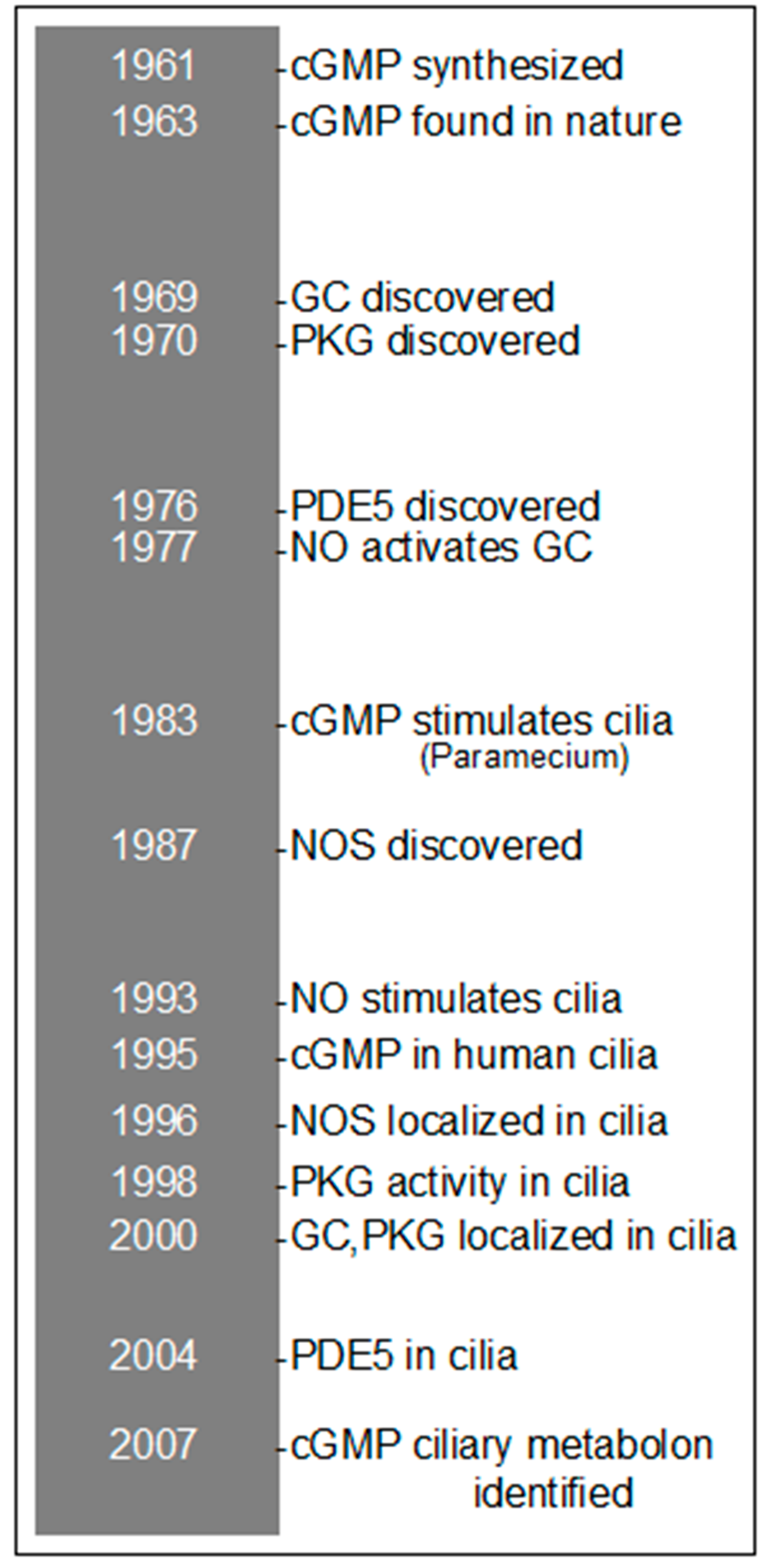
2. History of cGMP
3. Model Systems for Studying cGMP Regulation of Cilia
3.1. Photoreceptor
3.2. Olfactory Receptor Cilia
4. cGMP Regulation of Protozoan Cilia
4.1. Paramecium
4.2. Tetrahymena
5. Other Models of cGMP and Cilia Action
5.1. Animal Models of cGMP-Mediated Cilia Motility
5.1.1. Frog
5.1.2. Rat
5.1.3. Rabbit
5.1.4. Porcine
5.1.5. Bovine
5.1.6. Human
5.1.7. Mouse
6. Nitric Oxide and Cilia
7. Alcohol and the NO-cGMP Ciliary Response
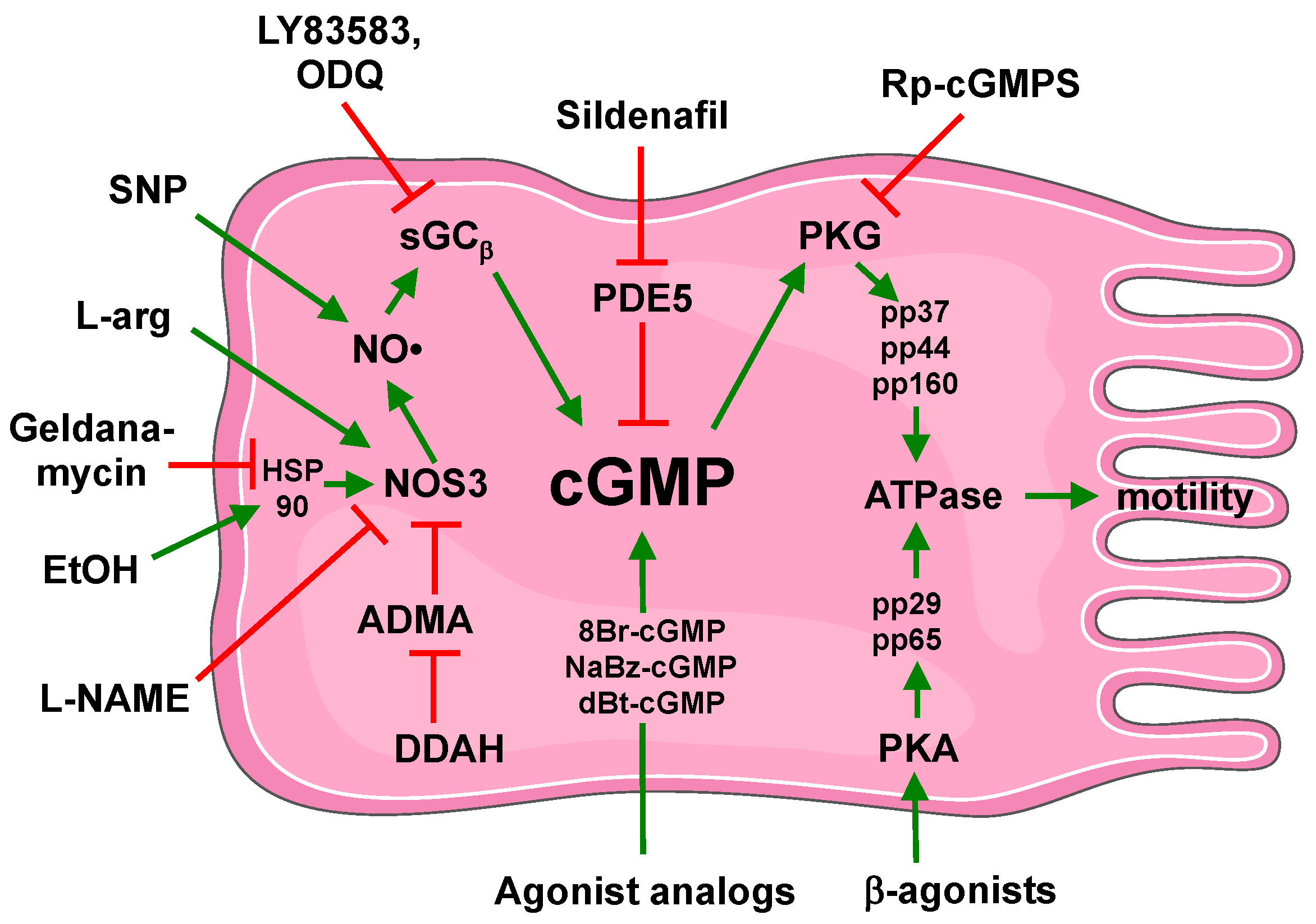
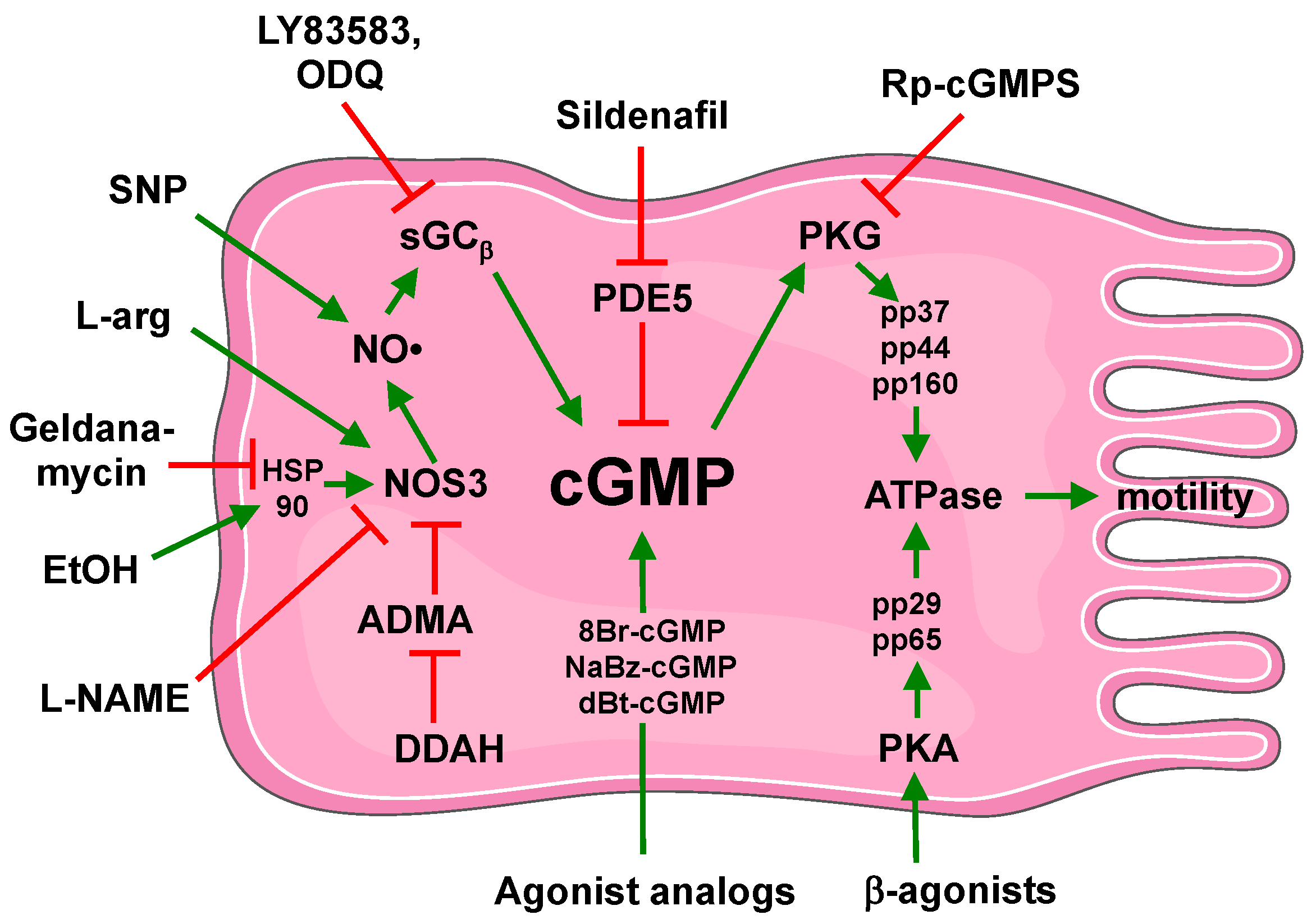
8. Conclusion
Acknowledgments
Conflicts of Interest
References
- Elferink, J.G.; VanUffelen, B.E. The role of cyclic nucleotides in neutrophil migration. Gen. Pharmacol. 1996, 27, 387–393. [Google Scholar] [CrossRef]
- Van Haastert, P.J.; Kuwayama, H. cGMP as second messenger during dictyostelium chemotaxis. FEBS Lett. 1997, 410, 25–28. [Google Scholar] [CrossRef]
- Bicker, G. STOP and GO with NO: Nitric oxide as a regulator of cell motility in simple brains. Bioessays 2005, 27, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, T.M.; Cornwell, T.L. Towards an understanding of the mechanism of action of cyclic AMP and cyclic GMP in smooth muscle relaxation. Blood Vessels 1991, 28, 129–137. [Google Scholar] [PubMed]
- Dubovskaya, L.V.; Bakakina, Y.S.; Kolesneva, E.V.; Sodel, D.L.; McAinsh, M.R.; Hetherington, A.M.; et al. cGMP-dependent ABA-induced stomatal closure in the ABA-insensitive arabidopsis mutant abi1-1. New Phytol. 2011, 191, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Michael Smith-Nobel lecture: Synthetic DNA and biology. Available online: http://www.nobelprize.org/nobel_prizes/chemistry/laureates/1993/smith-lecture.html (accessed on 8 May 2015).
- Ashman, D.F.; Lipton, R.; Melicow, M.M.; Price, T.D. Isolation of adenosine 3′,5′-monophosphate and guanosine 3′,5′-monophosphate from rat urine. Biochem. Biophys. Res. Commun. 1963, 11, 330–334. [Google Scholar] [CrossRef]
- Hardman, J.G.; Davis, J.W.; Sutherland, E.W. Measurement of guanosine 3′,5′-monophosphate and other cyclic nucleotides. variations in urinary excretion with hormonal state of the rat. J. Biol. Chem. 1966, 241, 4812–4815. [Google Scholar] [PubMed]
- Hardman, J.G.; Sutherland, E.W. Guanyl cyclase: An enzyme catalyzing the formation of guanosine 3′,5′-monophosphate from guanosine triphosphate. J. Biol. Chem. 1969, 244, 6363–6370. [Google Scholar] [PubMed]
- Rall, T.W.; Sutherland, E.W.; Berthet, J. The relationship of epinephrine and glucagon to liver phosphorylase. IV. Effect of epinephrine and glucagon on the reactivation of phosphorylase in liver homogenates. J. Biol. Chem. 1957, 224, 463–475. [Google Scholar]
- Francis, S.H.; Corbin, J.D. Structure and function of cyclic nucleotide-dependent protein kinases. Annu. Rev. Physiol. 1994, 56, 237–272. [Google Scholar] [CrossRef] [PubMed]
- Kuo, J.F.; Greengard, P. Cyclic nucleotide-dependent protein kinases VI. Isolation and partial purification of a protein kinase activated by guanosine 3′,5′-monophosphate. J. Biol. Chem. 1970, 245, 2493–2498. [Google Scholar] [PubMed]
- Lincoln, T.M.; Corbin, J.D. Characterization and biological role of the cGMP-dependent protein kinase. Adv. Cyclic Nucleotide Res. 1983, 15, 139–192. [Google Scholar]
- Stout, S.L.; Wyatt, T.A.; Adams, J.J.; Sisson, J.H. Nitric oxide-dependent cilia regulatory enzyme localization in bovine bronchial epithelial cells. J. Histochem. Cytochem. 2007, 55, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.H.; Thomas, M.K.; Corbin, J.D. Cyclic GMP-Binding Cyclic GMP-Specific Phosphodiesterase from Lung; John Wiley and Sons Ltd: New York, NY, USA, 1990. [Google Scholar]
- Lincoln, T.M.; Hall, C.L.; Park, C.R.; Corbin, J.D. Guanosine 3′,5′-monophosphate binding proteins in rat tissues. Proc. Natl. Acad. Sci. USA 1976, 73, 2559–2563. [Google Scholar] [CrossRef] [PubMed]
- Sisson, J.H.; Wyatt, T.A. Inhibition of PDE5 delays the onset of ethanol-mediated ciliary desensitization in bronchial epithelial cells. Alcohol. Clin. Exp. Res. 2004, 28, 13A. [Google Scholar] [CrossRef]
- Juilfs, D.M.; Fulle, H.J.; Zhao, A.Z.; Houslay, M.D.; Garbers, D.L.; Beavo, J.A. A subset of olfactory neurons that selectively express cGMP-stimulated phosphodiesterase (PDE2) and guanylyl cyclase-D define a unique olfactory signal transduction pathway. Proc. Natl. Acad. Sci. USA 1997, 94, 3388–3395. [Google Scholar] [CrossRef] [PubMed]
- Forget, M.A.; Sisson, J.H.; Spurzem, J.R.; Wyatt, T.A. Ethanol increases phosphodiesterase 4 activity in bovine bronchial epithelial cells. Alcohol 2003, 31, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Schraermeyer, U.; Esser, P.; Grisanti, S.; Rack, M.; Heimann, K. Cytochemical localization of guanylate cyclase in photoreceptor cells of the mouse. Graefes Arch. Clin. Exp. Ophthalmol. 1997, 235, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Giessl, A.; Pulvermuller, A.; Trojan, P.; Park, J.H.; Choe, H.W.; Ernst, O.P.; Hofmann, K.P.; Wolfrum, U. Differential expression and interaction with the visual G-protein transducin of centrin isoforms in mammalian photoreceptor cells. J. Biol. Chem. 2004, 279, 51472–51481. [Google Scholar] [CrossRef] [PubMed]
- Ortego, J.; Coca-Prados, M. Functional expression of components of the natriuretic peptide system in human ocular nonpigmented ciliary epithelial cells. Biochem. Biophys. Res. Commun. 1999, 258, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Del Pilar Gomez, M.; Nasi, E. Calcium-independent, cGMP-mediated light adaptation in invertebrate ciliary photoreceptors. J. Neurosci. 2005, 25, 2042–2049. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Gold, G.H. A cyclic nucleotide-gated conductance in olfactory receptor cilia. Nature 1987, 325, 442–444. [Google Scholar] [CrossRef] [PubMed]
- Dhallan, R.S.; Yau, K.W.; Schrader, K.A.; Reed, R.R. Primary structure and functional expression of a cyclic nucleotide-activated channel from olfactory neurons. Nature 1990, 347, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Breer, H.; Klemm, T.; Boekhoff, I. Nitric oxide mediated formation of cyclic GMP in the olfactory system. Neuroreport 1992, 3, 1030–1032. [Google Scholar] [CrossRef] [PubMed]
- Kurahashi, T.; Kaneko, A. Gating properties of the cAMP-gated channel in toad olfactory receptor cells. J. Physiol. 1993, 466, 287–302. [Google Scholar] [PubMed]
- Kroner, C.; Boekhoff, I.; Lohmann, S.M.; Genieser, H.G.; Breer, H. Regulation of olfactory signalling via cGMP-dependent protein kinase. Eur. J. Biochem. 1996, 236, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Leinders-Zufall, T.; Rand, M.N.; Shepherd, G.M.; Greer, C.A.; Zufall, F. Calcium entry through cyclic nucleotide-gated channels in individual cilia of olfactory receptor cells: Spatiotemporal dynamics. J. Neurosci. 1997, 17, 4136–4148. [Google Scholar] [PubMed]
- Dittman, A.H.; Quinn, T.P.; Nevitt, G.A.; Hacker, B.; Storm, D.R. Sensitization of olfactory guanylyl cyclase to a specific imprinted odorant in coho salmon. Neuron 1997, 19, 381–389. [Google Scholar] [CrossRef]
- Moon, C.; Jaberi, P.; Otto-Bruc, A.; Baehr, W.; Palczewski, K.; Ronnett, G.V. Calcium-sensitive particulate guanylyl cyclase as a modulator of cAMP in olfactory receptor neurons. J. Neurosci. 1998, 18, 3195–3205. [Google Scholar] [PubMed]
- Wyatt, T.A.; Forget, M.A.; Sisson, J.H. Ethanol stimulates ciliary beating by dual cyclic nucleotide kinase activation in bovine bronchial epithelial cells. Am. J. Pathol. 2003, 163, 1157–1166. [Google Scholar] [CrossRef]
- Pietrobon, M.; Zamparo, I.; Maritan, M.; Franchi, S.A.; Pozzan, T.; Lodovichi, C. Interplay among cGMP, cAMP, and Ca2+ in living olfactory sensory neurons in vitro and in vivo. J. Neurosci. 2011, 31, 8395–8405. [Google Scholar] [CrossRef] [PubMed]
- Bargmann, C.I. Chemosensation in C. elegans. WormBook 2006, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Bonini, N.M.; Nelson, D.L. Differential regulation of paramecium ciliary motility by cAMP and cGMP. J. Cell Biol. 1988, 106, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.E.; Guo, Y.; Kleefeld, G.; Volkel, H. Hyperpolarization- and depolarization-activated Ca2+ currents in paramecium trigger behavioral changes and cGMP formation independently. J. Membr. Biol. 1997, 156, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Andrivon, C. Membrane control of ciliary movement in ciliates. Biol. Cell 1988, 63, 133–142. [Google Scholar] [CrossRef]
- Walerczyk, M.; Fabczak, S. Additional evidence for the cyclic GMP signaling pathway resulting in the photophobic behavior of stentor coeruleus. Photochem. Photobiol. 2001, 74, 829–836. [Google Scholar] [CrossRef]
- Eistetter, H.; Seckler, B.; Bryniok, D.; Schultz, E. Phosphorylation of endogenous proteins of cilia from paramaceium tetraurelia in vitro. Eur. J. Cell Biol. 1983, 31, 220–226. [Google Scholar] [PubMed]
- Miglietta, L.A.; Nelson, D.L. A novel cGMP-dependent protein kinase from paramecium. J. Biol. Chem. 1988, 263, 16096–16105. [Google Scholar] [PubMed]
- Klumpp, S.; Steiner, A.L.; Schultz, J.E. Immunocytochemical localization of cyclic GMP, cGMP-dependent protein kinase, calmodulin and calcineurin in paramecium tetraurelia. Eur. J. Cell Biol. 1983, 32, 164–170. [Google Scholar] [PubMed]
- Kissmehl, R.; Kruger, T.P.; Treptau, T.; Froissard, M.; Plattner, H. Multigene family encoding 3′,5′-cyclic-GMP-dependent protein kinases in paramecium tetraurelia cells. Eukaryot. Cell 2006, 5, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Bell, W.E.; Hallworth, R.; Wyatt, T.A.; Sisson, J.H. Use of a novel cell adhesion method and digital measurement to show stimulus-dependent variation in somatic and oral ciliary beat frequency in paramecium. J. Eukaryot. Microbiol. 2015, 62, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Travis, S.M.; Nelson, D.L. Regulation of axonemal Mg2+-ATPase from paramecium cilia: Effects of Ca2+ and cyclic nucleotides. Biochim. Biophys. Acta 1988, 966, 84–93. [Google Scholar] [CrossRef]
- Walczak, C.E.; Nelson, D.L. In vitro phosphorylation of ciliary dyneins by protein kinases from paramecium. J. Cell Sci. 1993, 106, 1369–1376. [Google Scholar] [PubMed]
- Ann, K.S.; Nelson, D.L. Protein substrates for cGMP-dependent protein phosphorylation in cilia of wild type and atalanta mutants of paramecium. Cell Motil. Cytoskeleton. 1995, 30, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, M.; Kurahashi, S.; Kamachi, H.; Inoue, H. Control of the ciliary beat by cyclic nucleotides in intact cortical sheets from paramecium. Zoolog. Sci. 2004, 21, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Valeyev, N.V.; Heslop-Harrison, P.; Postlethwaite, I.; Gizatullina, A.N.; Kotov, N.V.; Bates, D.G. Crosstalk between G-protein and Ca2+ pathways switches intracellular cAMP levels. Mol. Biosyst. 2009, 5, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Joglar-Ramirez, J.A.; Renaud, F.L. Calcium regulates the regeneration of cilia in tetrahymena thermophila. J. Protozool. 1988, 35, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Kohidai, L.; Barsony, J.; Roth, J.; Marx, S.J. Rapid effects of insulin on cyclic GMP location in an intact protozoan. Experientia 1992, 48, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Doran, S.A.; Tran, C.H.; Eskicioglu, C.; Stachniak, T.; Ahn, K.C.; Goldberg, J.I. Constitutive and permissive roles of nitric oxide activity in embryonic ciliary cells. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R348–R355. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S. Pharmacological control of ciliary activity in the young sea urchin larva. Studies on the role of Ca2+ and cyclic nucleotides. Comp. Biochem. Physiol. C 1984, 78, 183–191. [Google Scholar] [CrossRef]
- Zagoory, O.; Braiman, A.; Priel, Z. The mechanism of ciliary stimulation by acetylcholine: Roles of calcium, PKA, and PKG. J. Gen. Physiol. 2002, 119, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Shirakami, G.; Zhan, X.; Johns, R.A. Regulation of ciliary beat frequency by the nitric oxide-cyclic guanosine monophosphate signaling pathway in rat airway epithelial cells. Am. J. Respir. Cell Mol. Biol. 2000, 23, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Shirakami, G.; Li, D.; Zhan, X.; Johns, R.A. Propofol stimulates ciliary motility via the nitric oxide-cyclic GMP pathway in cultured rat tracheal epithelial cells. Anesthesiology 2000, 93, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.A.; Pryzwansky, K.B.; Lincoln, T.M. KT5823 activates human neutrophils and fails to inhibit cGMP-dependent protein kinase phosphorylation of vimentin. Res. Commun. Chem. Pathol. Pharm. 1991, 74, 3–14. [Google Scholar]
- Tamaoki, J.; Kobayashi, K.; Sakai, N.; Kanemura, T.; Horii, S.; Isono, K.; Takeuchi, S.; Chiyotani, A.; Yamawaki, I.; Takizawa, T. Atrial natriuretic factor inhibits ciliary motility in cultured rabbit epithelium. Am. J. Physiol. 1991, 260, C201–C205. [Google Scholar] [PubMed]
- Uzlaner, N.; Priel, Z. Interplay between the NO pathway and elevated [Ca2+]i enhances ciliary activity in rabbit trachea. J. Physiol. 1999, 516, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sanderson, M.J. The role of cGMP in the regulation of rabbit airway ciliary beat frequency. J. Physiol. 2003, 551, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Gertsberg, I.; Hellman, V.; Fainshtein, M.; Weil, S.; Silberberg, S.D.; Danilenko, M.; Priel, Z. Intracellular Ca2+ regulates the phosphorylation and the dephosphorylation of ciliary proteins via the NO pathway. J. Gen. Physiol. 2004, 124, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.A.; Spurzem, J.R.; May, K.; Sisson, J.H. Regulation of ciliary beat frequency by both PKA and PKG in bovine airway epithelial cells. Am. J. Physiol. 1998, 275, L827–L835. [Google Scholar] [PubMed]
- Wyatt, T.A.; Forget, M.A.; Adams, J.M.; Sisson, J.H. Both cAMP and cGMP are required for maximal ciliary beat stimulation in a cell-free model of bovine ciliary axonemes. Am. J. Physiol. Lung. Cell Mol. Physiol. 2005, 288, L546–L551. [Google Scholar] [CrossRef] [PubMed]
- Sisson, J.H.; Pavlik, J.A.; Wyatt, T.A. Alcohol stimulates ciliary motility of isolated airway axonemes through a nitric oxide, cyclase, and cyclic nucleotide-dependent kinase mechanism. Alcohol. Clin. Exp. Res. 2009, 33, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Geary, C.A.; Davis, C.W.; Paradiso, A.M.; Boucher, R.C. Role of CNP in human airways: cGMP-mediated stimulation of ciliary beat frequency. Am. J. Physiol. 1995, 268, L1021–L1028. [Google Scholar] [PubMed]
- Yang, B.; Schlosser, R.J.; McCaffrey, T.V. Dual signal transduction mechanisms modulate ciliary beat frequency in upper airway epithelium. Am. J. Physiol. 1996, 270, L745–L751. [Google Scholar] [PubMed]
- Yang, B.; Schlosser, R.J.; McCaffrey, T.V. Signal transduction pathways in modulation of ciliary beat frequency by methacholine. Ann. Otol. Rhinol. Laryngol. 1997, 106, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Wang, H.; Lou, W.; Jin, S.; Fan, E.; Li, Y.; Han, D.; Zhang, L. Regulation of ciliary beat frequency by the nitric oxide signaling pathway in mouse nasal and tracheal epithelial cells. Exp. Cell Res. 2011, 317, 2548–2553. [Google Scholar] [CrossRef] [PubMed]
- Jain, B.; Rubinstein, I.; Robbins, R.A.; Leise, K.L.; Sisson, J.H. Modulation of airway epithelial cell ciliary beat frequency by nitric oxide. Biochem. Biophys. Res. Commun. 1993, 191, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Li, D.; Johns, R.A. Immunohistochemical evidence for the NO cGMP signaling pathway in respiratory ciliated epithelia of rat. J. Histochem. Cytochem. 1999, 47, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Simet, S.M.; Pavlik, J.A.; Sisson, J.H. Proteomic analysis of bovine axonemes exposed to acute alcohol: Role of endothelial nitric oxide synthase and heat shock protein 90 in cilia stimulation. Alcohol. Clin. Exp. Res. 2013, 37, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Simet, S.M.; Pavlik, J.A.; Sisson, J.H. Dietary antioxidants prevent alcohol-induced ciliary dysfunction. Alcohol. 2013, 47, 629–635. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wyatt, T.A.; Wells, S.M.; Alsaidi, Z.A.; DeVasure, J.M.; Klein, E.B.; Bailey, K.L.; Sisson, J.H. Asymmetric dimethylarginine blocks nitric oxide-mediated alcohol-stimulated cilia beating. Mediators Inflamm. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- De Sanctis, G.T.; MacLean, J.A.; Hamada, K.; Mehta, S.; Scott, J.A.; Jiao, A.; Yandava, C.N.; Kobzik, L.; Wolyniec, W.W.; Fabian, A.J.; et al. Contribution of nitric oxide synthases 1, 2, and 3 to airway hyperresponsiveness and inflammation in a murine model of asthma. J. Exp. Med. 1999, 189, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Sisson, J.H. Ethanol stimulates apparent nitric oxide-dependent ciliary beat frequency in bovine airway epithelial cells. Am. J. Physiol. 1995, 268, L596–L600. [Google Scholar] [PubMed]
- Tamaoki, J.; Chiyotani, A.; Kondo, M.; Konno, K. Role of NO generation in beta-adrenoceptor-mediated stimulation of rabbit airway ciliary motility. Am. J. Physiol. 1995, 268, C1342–C1347. [Google Scholar] [PubMed]
- Xue, C.; Botkin, S.J.; Johns, R.A. Localization of endothelial NOS at the basal microtubule membrane in ciliated epithelium of rat lung. J. Histochem. Cytochem. 1996, 44, 463–471. [Google Scholar] [PubMed]
- Runer, T.; Lindberg, S. Ciliostimulatory effects mediated by nitric oxide. Acta Otolaryngol. 1999, 119, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Sisson, J.H.; May, K.; Wyatt, T.A. Nitric oxide-dependent ethanol stimulation of ciliary motility is linked to cAMP-dependent protein kinase (PKA) activation in bovine bronchial epithelium. Alcohol. Clin. Exp. Res. 1999, 23, 1528–1533. [Google Scholar] [PubMed]
- Piggott, L.A.; Hassell, K.A.; Berkova, Z.; Morris, A.P.; Silberbach, M.; Rich, T.C. Natriuretic peptides and nitric oxide stimulate cGMP synthesis in different cellular compartments. J. Gen. Physiol. 2006, 128, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Sherman, T.S.; Chen, Z.; Yuhanna, I.S.; Lau, K.S.; Margraf, L.R.; Shaul, P.W. Nitric oxide synthase isoform expression in the developing lung epithelium. Am. J. Physiol. 1999, 276, L383–L390. [Google Scholar] [PubMed]
- Chen, J.H.; Takeno, S.; Osada, R.; Ueda, T.; Yajin, K. Modulation of ciliary activity by tumor necrosis factor-alpha in cultured sinus epithelial cells. possible roles of nitric oxide. Hiroshima. J. Med. Sci. 2000, 49, 49–55. [Google Scholar] [PubMed]
- Deja, M.; Busch, T.; Bachmann, S.; Riskowski, K.; Campean, V.; Wiedmann, B.; Schwabe, M.; Hell, B.; Pfeilschifter, J.; Falke, K.J.; et al. Reduced nitric oxide in sinus epithelium of patients with radiologic maxillary sinusitis and sepsis. Am. J. Respir. Crit. Care Med. 2003, 168, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Pifferi, M.; Bush, A.; Maggi, F.; Michelucci, A.; Ricci, V.; Conidi, M.E.; Cangiotti, A.M.; Bodini, A.; Simi, P.; Macchia, P.; et al. Nasal nitric oxide and nitric oxide synthase expression in primary ciliary dyskinesia. Eur. Respir. J. 2011, 37, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.L.; Lucas, J.S.; Walker, W.T.; Owen, H.; Premadeva, I.; Lackie, P.M. Neuronal NOS localises to human airway cilia. Nitric Oxide 2015, 44, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Sisson, J.H. Alcohol and airways function in health and disease. Alcohol 2007, 41, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.A.; Noll, E.C.; Rodney, W.M. Survival after a serum ethanol concentration of 1 1/2%. Lancet 1982, 320. [Google Scholar] [CrossRef]
- Wyatt, T.A.; Gentry-Nielsen, M.J.; Pavlik, J.A.; Sisson, J.H. Desensitization of PKA-stimulated ciliary beat frequency in an ethanol-fed rat model of cigarette smoke exposure. Alcohol. Clin. Exp. Res. 2004, 28, 998–1004. [Google Scholar] [CrossRef] [PubMed]
- Elliott, M.K.; Sisson, J.H.; Wyatt, T.A. Effects of cigarette smoke and alcohol on ciliated tracheal epithelium and inflammatory cell recruitment. Am. J. Respir. Cell Mol. Biol. 2007, 36, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Lazic, T.; Wyatt, T.A.; Matic, M.; Meyerholz, D.K.; Grubor, B.; Gallup, J.M.; Kersting, K.W.; Imerman, P.M.; Almeida-De-Macedo, M.; Ackermann, M.R. Maternal alcohol ingestion reduces surfactant protein A expression by preterm fetal lung epithelia. Alcohol 2007, 41, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.A.; Sisson, J.H. Chronic ethanol downregulates PKA activation and ciliary beating in bovine bronchial epithelial cells. Am. J. Physiol. Lung. Cell Mol. Physiol. 2001, 281, L575–L581. [Google Scholar] [PubMed]
- Wyatt, T.A.; Nicolarsen, J.; Jerrells, T.R.; Sisson, J.H. Localization and enzyme activity of protein kinase C epsilon (PKCe) in ciliated bovine bronchial epithelial cells. FASEB J. 2004, 18, A948. [Google Scholar]
- Price, M.E.; Pavlik, J.A.; Sisson, J.H.; Wyatt, T.A. Inhibition of protein phosphatase 1 reverses alcohol-induced ciliary dysfunction. Am. J. Physiol. Lung. Cell Mol. Physiol. 2015, 308, L577–L585. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Pavlik, J.; Fox, L.; Scarbrough, C.; Sale, W.S.; Sisson, J.H.; Wirschell, M. Alcohol-induced ciliary dysfunction targets the outer dynein arm. Am. J. Physiol. Lung. Cell Mol. Physiol. 2015, 308, L569–L576. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wyatt, T.A. Cyclic GMP and Cilia Motility. Cells 2015, 4, 315-330. https://doi.org/10.3390/cells4030315
Wyatt TA. Cyclic GMP and Cilia Motility. Cells. 2015; 4(3):315-330. https://doi.org/10.3390/cells4030315
Chicago/Turabian StyleWyatt, Todd A. 2015. "Cyclic GMP and Cilia Motility" Cells 4, no. 3: 315-330. https://doi.org/10.3390/cells4030315
APA StyleWyatt, T. A. (2015). Cyclic GMP and Cilia Motility. Cells, 4(3), 315-330. https://doi.org/10.3390/cells4030315