



Combined Transplantation of Mesenchymal Progenitor and Neural Stem Cells to Repair Cervical Spinal Cord Injury
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Mesenchymal Progenitor Cell Culture
2.3. Neural Stem Cell Culture
2.4. Surgeries
2.5. MPCs Preparation for IV Injections
2.6. Intravenous Injections of MPCs
2.7. NSCs Preparation for Intraspinal Injections
2.8. Intraspinal Injections of NSCs
2.9. Group Designation
2.10. Bioluminescence
2.11. Behavior—Mouse Cylinder
2.12. NPCs and MSCs Culture
2.13. MSC-Driven NSC Differentiation
2.14. Immunocytochemistry
2.15. Histology
2.16. Immunostaining (Tissues)
2.17. Intraspinal Injection NSC Counts
2.18. Statistics
2.18.1. In Vivo Analysis
2.18.2. In Vitro Analysis
3. Results
3.1. Bioluminescence Imaging Is an Effective Tracker of Intraspinally Injected NSCs
3.2. Dual-IV and IS Injection of MPCs and NSCs in Unilateral SCI Shows Limited Capacity to Improve Forelimb Function
3.3. GFP+-NSCs Isolated from the Subventricular Zone Can Survive and Differentiate in the Cervically Injured Spinal Cord
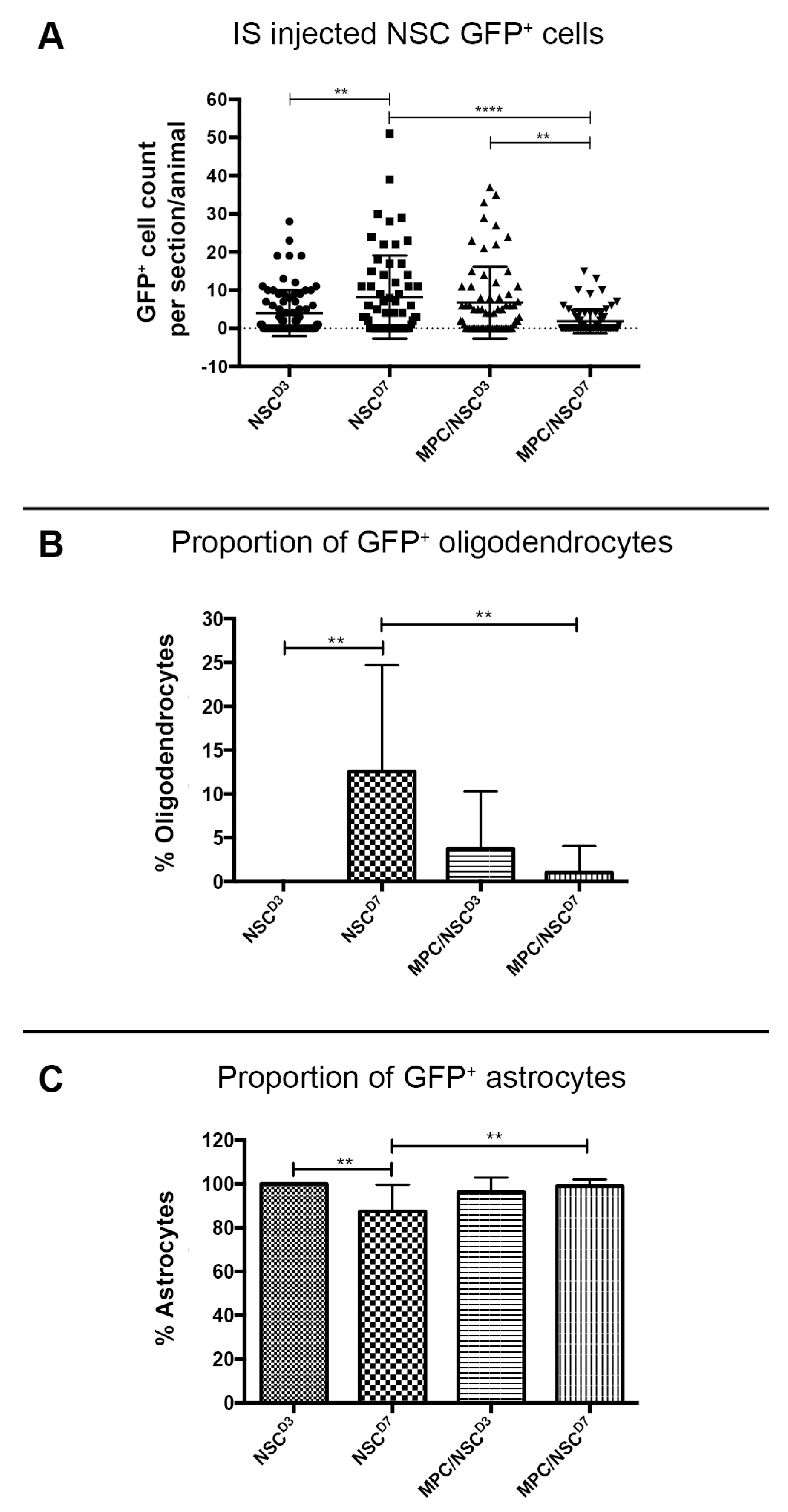
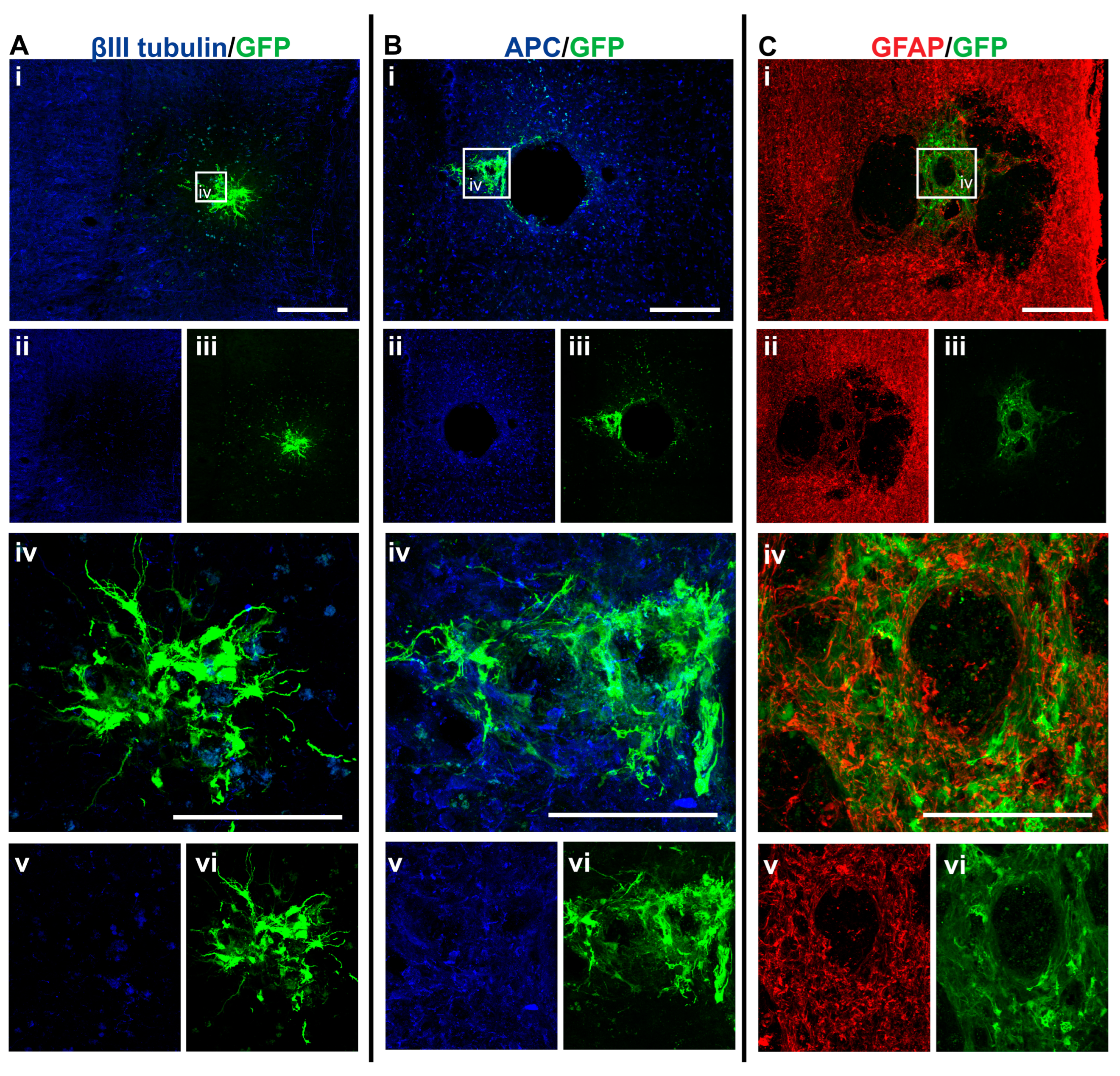

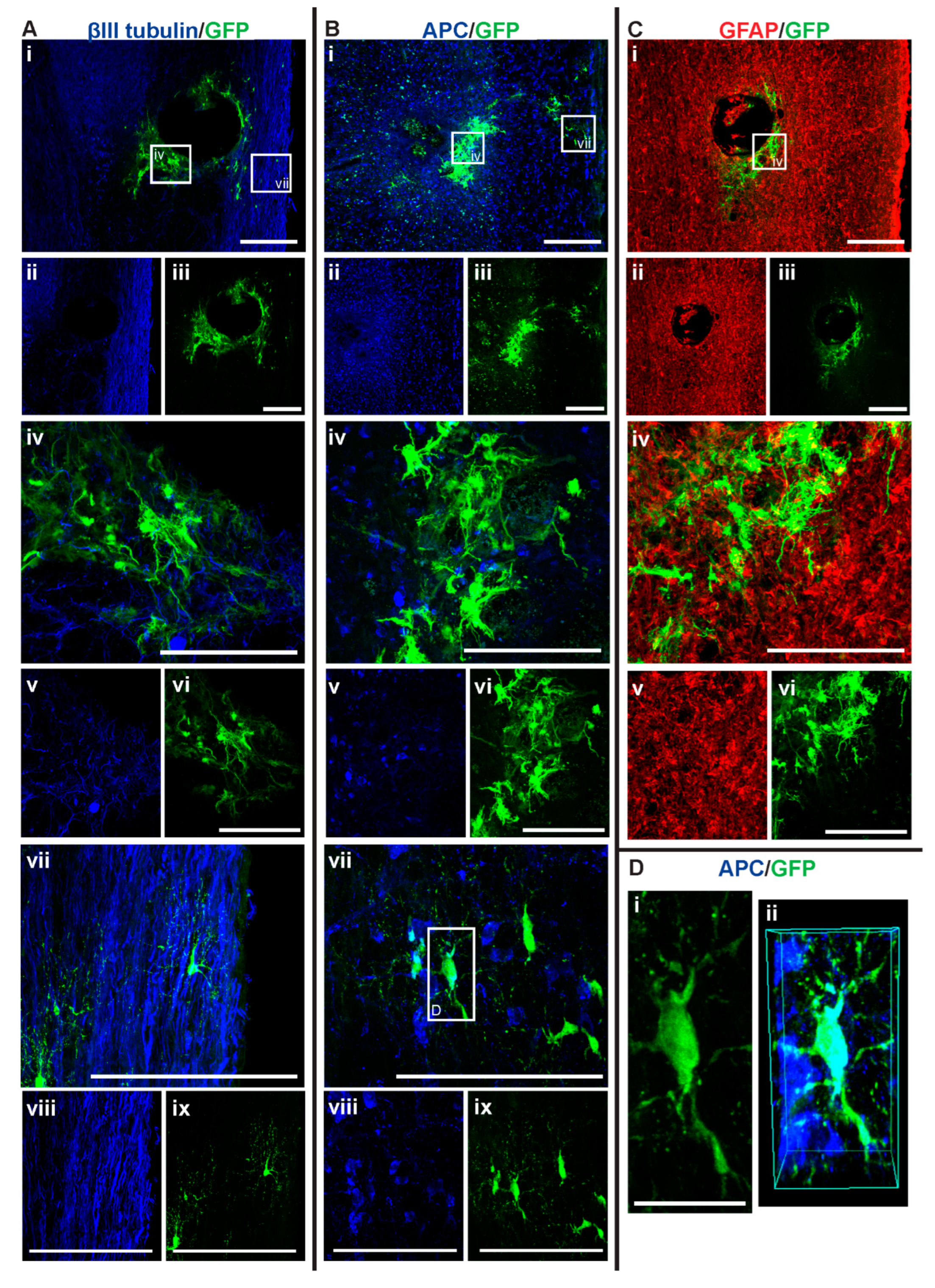
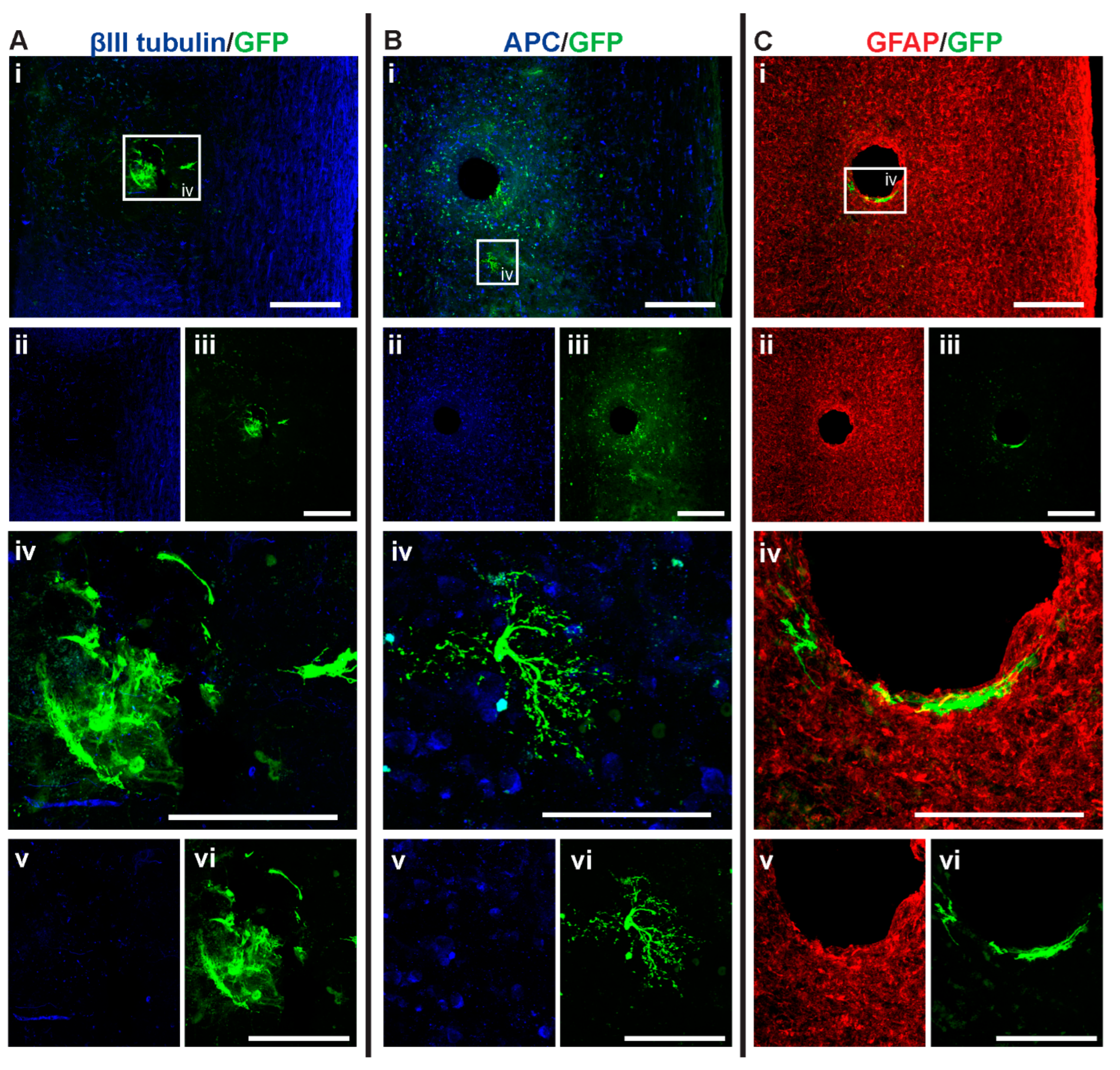
3.4. Intraspinal Injection of NSCs Results in Cavity Formation at the Injection Site
3.5. Intraspinal Injection at Day Seven Increased Survival of NSCs and Promoted Oligodendroglia Differentiation
3.6. Injection of IV MPCs Increased NSC Differentiation into Oligodendrocytes at Day Three Injection Timepoint
3.7. Combination of IV Injection of MPCs with Intraspinal Injection of NSCs at D7 Resulted in Reduced NSC Survival and Differentiation Capacity
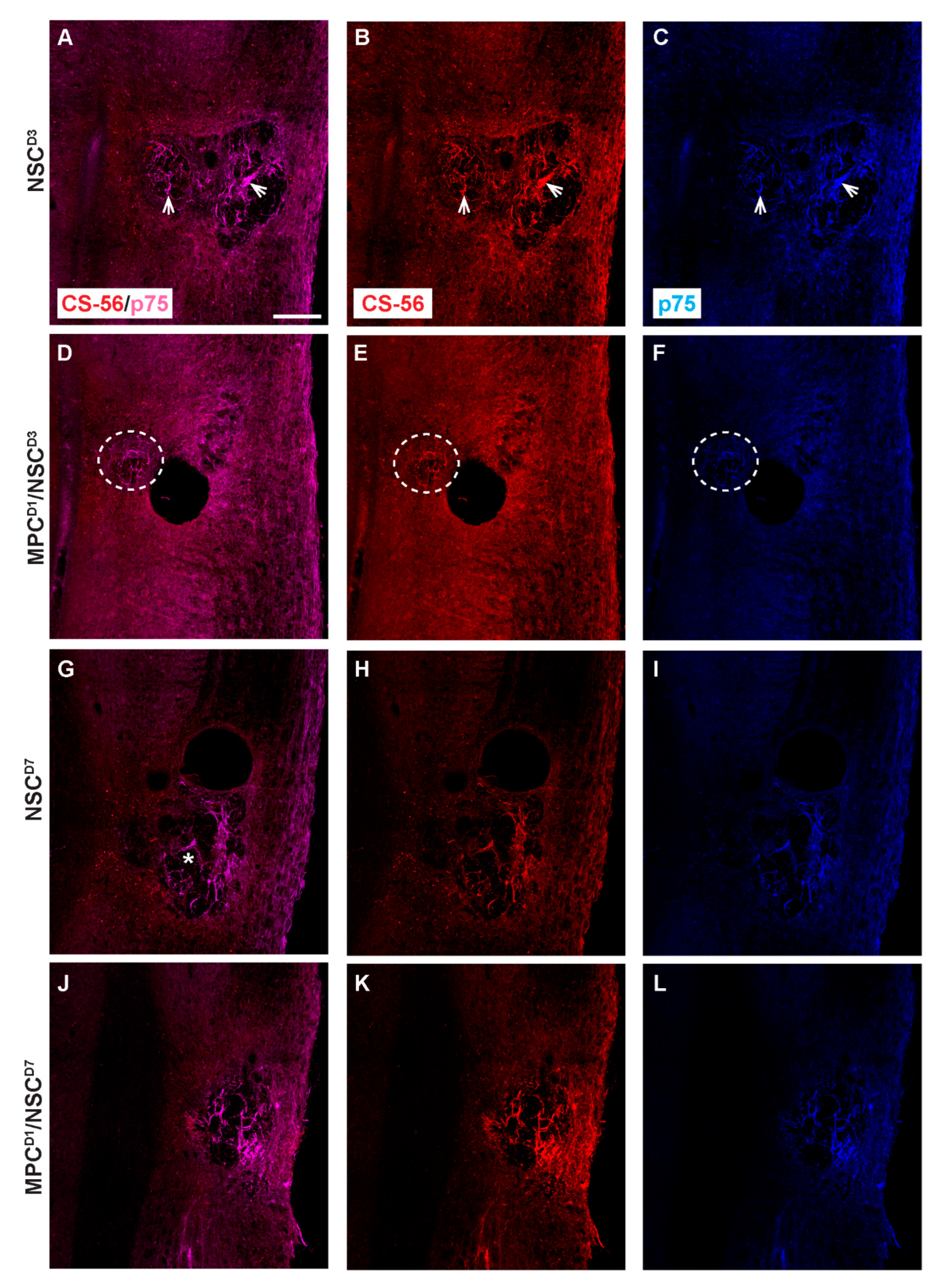
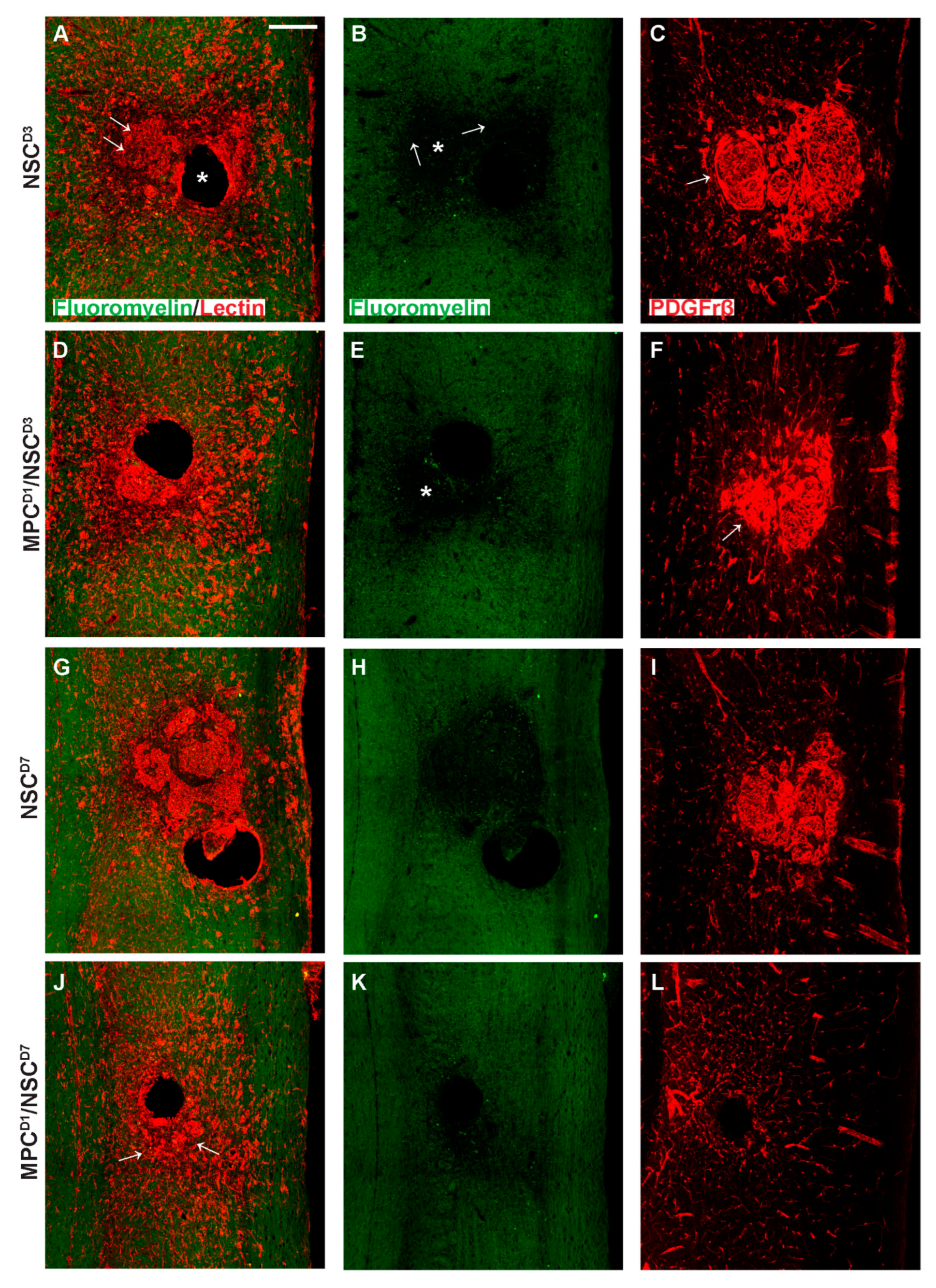
3.8. Distribution of Proteoglycan Deposition and Schwann Cell Ingression
3.9. Extracellular Matrix, Deposition, and Pericyte Infiltration at the Site of NSC Transplantation Is Limited by Prior Injection of MPCs at Day One
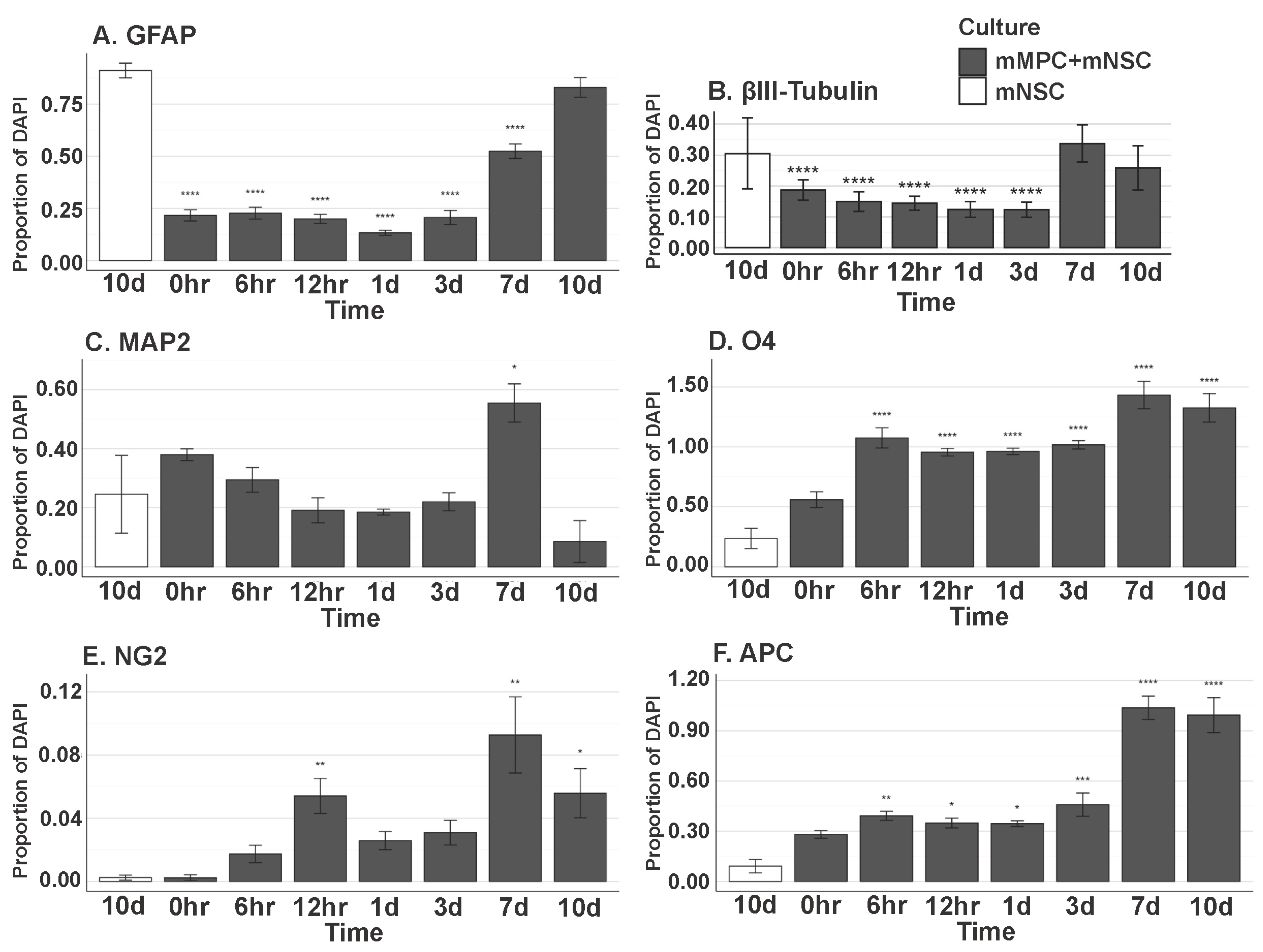
3.10. In Vitro MPC/NSC Co-Cultures Show the Increased Oligodendrocyte Cell Fate for NSCs
4. Discussion
4.1. NSC Differentiation into Oligodendroglia Increased in the Presence of MPCs
4.2. NSCs Isolated from the Subventricular Zone Can Survive and Differentiate in the Injured Cervical Spinal Cord
4.3. Intraspinal Injection at Day Seven Increased Survival of NSCs and Promoted Oligodendrocyte Differentiation
4.4. Injection of IV MPCs Altered NSC Differentiation in Mice Receiving Intraspinal Injection of NSCs at D3
4.5. Combination of IV Injection of MPCs with Intraspinal Injection of NSCs at D7 Resulted in Reduced NSC Survival and Differentiation Capacity
4.6. Changes in Proteoglycan Deposition, Vascular, and Pericyte/Schwann Cell Ingression-Role in Tissue Repair
4.7. Dual-Injection of MPCs and NSCs in Unilateral SCI Shows Limited Capacity to Improve Forelimb Function
4.8. Off the Shelf Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Abbreviations | n |
|---|---|---|
| Intraspinal HBSS D3 | HBSSD3 | 10 |
| Intraspinal NSC D3 | NSC_D3 | 8 |
| Intravenous HBSS D1 with intraspinal HBSS D3 | HBSSD1/HBSSD3 | 9 |
| Intravenous MPC D1 with intraspinal NSC D3 | MPCD1/NSC_D3 | 9 |
| Intraspinal HBSS D7 | HBSSD7 | 9 |
| Intraspinal NSC D7 | NSC_D7 | 8 |
| Intravenous HBSS D1 with intraspinal HBSS D7 | HBSSD1/HBSSD7 | 10 |
| Intravenous MPC D1 with intraspinal NSC D7 | MPCD1/NSC_D7 | 9 |
References
- Ruitenberg, M.J.; Vukovic, J.; Sarich, J.; Busfield, S.J.; Plant, G.W. Olfactory ensheathing cells: Characteristics, genetic engineering, and therapeutic potential. J. Neurotrauma 2006, 23, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Tetzlaff, W.; Okon, E.B.; Karimi-Abdolrezaee, S.; Hill, C.E.; Sparling, J.S.; Plemel, J.R.; Plunet, W.T.; Tsai, E.C.; Baptiste, D.; Smithson, L.J.; et al. A systematic review of cellular transplantation therapies for spinal cord injury. J. Neurotrauma 2011, 28, 1611–1682. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.S.; Harvey, A.R.; Plant, G.W.; Hodgetts, S.I. Systematic review of induced pluripotent stem cell technology as a potential clinical therapy for spinal cord injury. Cell Transplant. 2013, 22, 571–617. [Google Scholar] [CrossRef]
- Silver, J.; Schwab, M.E.; Popovich, P.G. Central nervous system regenerative failure: Role of oligodendrocytes, astrocytes, and microglia. Cold Spring Harb. Perspect. Biol. 2015, 7, a020602. [Google Scholar] [CrossRef]
- Forostyak, S.; Jendelova, P.; Sykova, E. The role of mesenchymal stromal cells in spinal cord injury, regenerative medicine and possible clinical applications. Biochimie 2013, 95, 2257–2270. [Google Scholar] [CrossRef]
- Dasari, V.R.; Veeravalli, K.K.; Dinh, D.H. Mesenchymal stem cells in the treatment of spinal cord injuries: A review. World J. Stem Cells 2014, 6, 120–133. [Google Scholar] [CrossRef]
- Hodgetts, S.I.; Simmons, P.J.; Plant, G.W. Human mesenchymal precursor cells (Stro-1(+)) from spinal cord injury patients improve functional recovery and tissue sparing in an acute spinal cord injury rat model. Cell Transplant. 2013, 22, 393–412. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Dunne, C.; Hewson, J.; Wohl, C.; Wheatley, M.; Peterson, A.C.; Reynolds, B.A. Multipotent CNS stem cells are present in the adult mammalian spinal cord and ventricular neuroaxis. J. Neurosci. 1996, 16, 7599–7609. [Google Scholar] [CrossRef]
- Wilcox, J.T.; Satkunendrarajah, K.; Zuccato, J.A.; Nassiri, F.; Fehlings, M.G. Neural precursor cell transplantation enhances functional recovery and reduces astrogliosis in bilateral compressive/contusive cervical spinal cord injury. Stem Cells Transl. Med. 2014, 3, 1148–1159. [Google Scholar] [CrossRef]
- Bonner, J.F.; Steward, O. Repair of spinal cord injury with neuronal relays: From fetal grafts to neural stem cells. Brain Res. 2015, 1619, 115–123. [Google Scholar] [CrossRef]
- Grégoire, C.; Goldenstein, B.L.; Floriddia, E.M.; Barnabé-Heider, F.; Fernandes, K.J.L. Endogenous neural stem cell responses to stroke and spinal cord injury. Glia 2015, 63, 1469–1482. [Google Scholar] [CrossRef] [PubMed]
- Piltti, K.M.; Avakian, S.N.; Funes, G.M.; Hu, A.; Uchida, N.; Anderson, A.J.; Cummings, B.J. Transplantation dose alters the dynamics of human neural stem cell engraftment, proliferation and migration after spinal cord injury. Stem Cell Res. 2015, 15, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Salewski, R.P.; Mitchell, R.A.; Shen, C.; Fehlings, M.G. Transplantation of neural stem cells clonally derived from embryonic stem cells promotes recovery after murine spinal cord injury. Stem Cells Dev. 2015, 24, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Stenudd, M.; Sabelström, H.; Frisén, J. Role of endogenous neural stem cells in spinal cord injury and repair. JAMA Neurol. 2015, 72, 235–237. [Google Scholar] [CrossRef]
- Bonner, J.F.; Blesch, A.; Neuhuber, B.; Fischer, I. Promoting directional axon growth from neural progenitors grafted into the injured spinal cord. J. Neurosci. Res. 2010, 88, 1182–1192. [Google Scholar] [CrossRef]
- Plant, G.W.; Christensen, C.L.; Oudega, M.; Bunge, M.B. Delayed transplantation of olfactory ensheathing glia promotes sparing/regeneration of supraspinal axons in the contused adult rat spinal cord. J. Neurotrauma 2003, 20, 1–16. [Google Scholar] [CrossRef]
- Cummings, B.J.; Uchida, N.; Tamaki, S.J.; Salazar, D.L.; Hooshmand, M.; Summers, R.; Gage, F.H.; Anderson, A.J. Human neural stem cells differentiate and promote locomotor recovery in spinal cord-injured mice. Proc. Natl. Acad. Sci. USA 2005, 102, 14069–14074. [Google Scholar] [CrossRef]
- Lee, S.V.; Czisch, C.E.; Huang, Y.; Han, M.H.; Harvey, A.R.; Plant, G.W. Early intravenous delivery of mesenchymal progenitor cells leads to behavioral and pathological amelioration after cervical spinal cord injury. In Proceedings of the Society for Neuroscience 44th Annual Meeting: Abstract No. 4812, Washington, DC, USA, 15 November 2014. [Google Scholar]
- White, S.V.; Czisch, C.E.; Han, M.H.; Plant, C.D.; Harvey, A.R.; Plant, G.W. Intravenous Transplantation of Mesenchymal Progenitors Distribute Solely to the Lungs and Improve Outcomes in Cervical Spinal Cord Injury. Stem Cells 2016, 34, 1812–1825. [Google Scholar] [CrossRef]
- Oh, J.S.; Kim, K.N.; An, S.S.; Pennant, W.A.; Kim, H.J.; Gwak, S.J.; Yoon, D.H.; Lim, M.H.; Choi, B.H.; Ha, Y. Cotransplantation of mouse neural stem cells (mNSCs) with adipose tissue-derived mesenchymal stem cells improves mNSC survival in a rat spinal cord injury model. Cell Transplant. 2011, 20, 837–849. [Google Scholar] [CrossRef]
- Short, B.J.; Brouard, N.; Simmons, P.J. Prospective isolation of mesenchymal stem cells from mouse compact bone. Methods Mol. Biol. 2009, 482, 259–268. [Google Scholar]
- Popovich, P.G.; Wei, P.; Stokes, B.T. Cellular inflammatory response after spinal cord injury in Sprague-Dawley and Lewis rats. J. Comp. Neurol. 1997, 377, 443–464. [Google Scholar] [CrossRef]
- Cao, Y.A.; Wagers, A.J.; Beilhack, A.; Dusich, J.; Bachmann, M.H.; Negrin, R.S.; Weissman, I.L.; Contag, C.H. Shifting foci of hematopoiesis during reconstitution from single stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 221–226. [Google Scholar] [CrossRef] [PubMed]
- van der Bogt, K.E.; Hellingman, A.A.; Lijkwan, M.A.; Bos, E.J.; de Vries, M.R.; van Rappard, J.R.; Fischbein, M.P.; Quax, P.H.; Robbins, R.C.; Hamming, J.F.; et al. Molecular imaging of bone marrow mononuclear cell survival and homing in murine peripheral artery disease. JACC Cardiovasc. Imaging 2012, 5, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Azari, H.; Rahman, M.; Sharififar, S.; Reynolds, B.A. Isolation and expansion of the adult mouse neural stem cells using the neurosphere assay. J. Vis. Exp. 2010, e2393. [Google Scholar] [CrossRef] [PubMed]
- Schallert, T.; Fleming, S.M.; Leasure, J.L.; Tillerson, J.L.; Bland, S.T. CNS plasticity and assessment of forelimb sensorimotor outcome in unilateral rat models of stroke, cortical ablation, parkinsonism and spinal cord injury. Neuropharmacology 2000, 39, 777–787. [Google Scholar] [CrossRef]
- Plant, G.W.; Bates, M.L.; Bunge, M.B. Inhibitory proteoglycan immunoreactivity is higher at the caudal than the rostral Schwann cell graft-transected spinal cord interface. Mol. Cell Neurosci. 2001, 17, 471–487. [Google Scholar] [CrossRef]
- Lankford, K.L.; Arroyo, E.J.; Nazimek, K.; Bryniarski, K.; Askenase, P.W.; Kocsis, J.D. Intravenously delivered mesenchymal stem cell-derived exosomes target M2-type macrophages in the injured spinal cord. PLoS ONE 2018, 13, e0190358. [Google Scholar] [CrossRef]
- Nakazaki, M.; Morita, T.; Lankford, K.L.; Askenase, P.W.; Kocsis, J.D. Small extracellular vesicles released by infused mesenchymal stromal cells target M2 macrophages and promote TGF-β upregulation, microvascular stabilization and functional recovery in a rodent model of severe spinal cord injury. J. Extracell. Vesicles 2021, 10, e12137. [Google Scholar] [CrossRef]
- Martin, D.; Robe, P.; Franzen, R.; Delree, P.; Schoenen, J.; Stevenaert, A.; Moonen, G. Effects of Schwann cell transplantation in a contusion model of rat spinal cord injury. J. Neurosci. Res. 1996, 45, 588–597. [Google Scholar] [CrossRef]
- Barbour, H.R.; Plant, C.D.; Harvey, A.R.; Plant, G.W. Tissue sparing, behavioral recovery, supraspinal axonal sparing/regeneration following sub-acute glial transplantation in a model of spinal cord contusion. BMC Neurosci. 2013, 14, 106. [Google Scholar] [CrossRef]
- Wichterle, H.; Lieberam, I.; Porter, J.A.; Jessell, T.M. Directed differentiation of embryonic stem cells into motor neurons. Cell 2002, 110, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Sigurjonsson, O.E.; Perreault, M.C.; Egeland, T.; Glover, J.C. Adult human hematopoietic stem cells produce neurons efficiently in the regenerating chicken embryo spinal cord. Proc. Natl. Acad. Sci. USA 2005, 102, 5227–5232. [Google Scholar] [CrossRef] [PubMed]
- Bible, E.; Qutachi, O.; Chau, D.Y.; Alexander, M.R.; Shakesheff, K.M.; Modo, M. Neo-vascularization of the stroke cavity by implantation of human neural stem cells on VEGF-releasing PLGA microparticles. Biomaterials 2012, 33, 7435–7446. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Wang, Y.; Graham, L.; McHale, K.; Gao, M.; Wu, D.; Brock, J.; Blesch, A.; Rosenzweig, E.S.; Havton, L.A.; et al. Long-distance growth and connectivity of neural stem cells after severe spinal cord injury. Cell 2012, 150, 1264–1273. [Google Scholar] [CrossRef]
- Gao, M.; Lu, P.; Bednark, B.; Lynam, D.; Conner, J.M.; Sakamoto, J.; Tuszynski, M.H. Templated agarose scaffolds for the support of motor axon regeneration into sites of complete spinal cord transection. Biomaterials 2013, 34, 1529–1536. [Google Scholar] [CrossRef]
- Cao, Q.L.; Howard, R.M.; Dennison, J.B.; Whittemore, S.R. Differentiation of engrafted neuronal-restricted precursor cells is inhibited in the traumatically injured spinal cord. Exp. Neurol. 2002, 177, 349–359. [Google Scholar] [CrossRef]
- Sontag, C.J.; Uchida, N.; Cummings, B.J.; Anderson, A.J. Injury to the spinal cord niche alters the engraftment dynamics of human neural stem cells. Stem Cell Rep. 2014, 2, 620–632. [Google Scholar] [CrossRef]
- Nishida, A.; Takahashi, M.; Tanihara, H.; Nakano, I.; Takahashi, J.B.; Mizoguchi, A.; Ide, C.; Honda, Y. Incorporation and differentiation of hippocampus-derived neural stem cells transplanted in injured adult rat retina. Invest. Ophthalmol. Vis. Sci. 2000, 41, 4268–4274. [Google Scholar]
- Ma, Y.H.E.; Putta, A.R.; Chan, C.H.H.; Vidman, S.R.; Monje, P.; Plant, G.W. Efficacy of Deferoxamine Mesylate in Serum and Serum-Free Media: Adult Ventral Root Schwann Cell Survival Following Hydrogen Peroxide-Induced Cell Death. Cells 2025, 14, 461. [Google Scholar] [CrossRef]
- Shihabuddin, L.S.; Horner, P.J.; Ray, J.; Gage, F.H. Adult spinal cord stem cells generate neurons after transplantation in the adult dentate gyrus. J. Neurosci. 2000, 20, 8727–8735. [Google Scholar] [CrossRef]
- Suh, H.; Zhou, Q.-G.; Fernandez-Carasa, I.; Clemenson, G.D.; Pons-Espinal, M.; Ro, E.J.; Marti, M.; Raya, A.; Gage, F.H.; Consiglio, A. Long-Term Labeling of Hippocampal Neural Stem Cells by a Lentiviral Vector. Front. Mol. Neurosci. 2018, 11, 415. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Graham, L.; Wang, Y.; Wu, D.; Tuszynski, M. Promotion of survival and differentiation of neural stem cells with fibrin and growth factor cocktails after severe spinal cord injury. J. Vis. Exp. 2014, e50641. [Google Scholar] [CrossRef]
- Wilems, T.S.; Pardieck, J.; Iyer, N.; Sakiyama-Elbert, S.E. Combination therapy of stem cell derived neural progenitors and drug delivery of anti-inhibitory molecules for spinal cord injury. Acta Biomater. 2015, 28, 23–32. [Google Scholar] [CrossRef] [PubMed]



| Primary Antibody | Supplier | Host | Dilution |
|---|---|---|---|
| Beta-III Tubulin | Aves Lab, Inc. (Davis, CA, USA) | Chicken | 1:1000 |
| O4 | EMD Millipore (Burlington, MA, USA) | Mouse | 1:50 |
| GFAP | Dako (Santa Clara, CA, USA) | Rabbit | 1:500 |
| MAP2 | Aves Lab, Inc. (Davis, CA, USA) | Chicken | 1:1000 |
| APC | CalBiochem (San Diego, CA, USA) | Mouse | 1:200 |
| GFAP | Dako (Santa Clara, MA, USA) | Rabbit | 1:500 |
| TuJ1 | Biolegends (San Diego, CA, USA) | Mouse | 1:500 |
| NG2 Chondroitin Sulfate | EMD Millipore (Burlington, MA, USA) | Rabbit | 1:500 |
| GFAP | Aves Lab, Inc. (Davis, CA, USA) | Chicken | 1:500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, S.V.; Ma, Y.H.E.; Plant, C.D.; Harvey, A.R.; Plant, G.W. Combined Transplantation of Mesenchymal Progenitor and Neural Stem Cells to Repair Cervical Spinal Cord Injury. Cells 2025, 14, 630. https://doi.org/10.3390/cells14090630
White SV, Ma YHE, Plant CD, Harvey AR, Plant GW. Combined Transplantation of Mesenchymal Progenitor and Neural Stem Cells to Repair Cervical Spinal Cord Injury. Cells. 2025; 14(9):630. https://doi.org/10.3390/cells14090630
Chicago/Turabian StyleWhite, Seok Voon, Yee Hang Ethan Ma, Christine D. Plant, Alan R. Harvey, and Giles W. Plant. 2025. "Combined Transplantation of Mesenchymal Progenitor and Neural Stem Cells to Repair Cervical Spinal Cord Injury" Cells 14, no. 9: 630. https://doi.org/10.3390/cells14090630
APA StyleWhite, S. V., Ma, Y. H. E., Plant, C. D., Harvey, A. R., & Plant, G. W. (2025). Combined Transplantation of Mesenchymal Progenitor and Neural Stem Cells to Repair Cervical Spinal Cord Injury. Cells, 14(9), 630. https://doi.org/10.3390/cells14090630