
Pharmacological Inhibition of Microglial Proliferation Supports Blood–Brain Barrier Integrity in Experimental Autoimmune Encephalomyelitis

 ,
,  ,
,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. EAE Induction and GW2580 Treatment
2.3. Spinal Cord mRNA Analysis
2.4. Histology
2.5. Immunohistochemistry and Quantification
2.6. Gelatin Gel Zymography
2.7. Functional Enrichment Analysis
2.8. Statistical Analysis
3. Results
3.1. GW2580 Decreased EAE Clinical Severity and Prevents the Relapse Phase
3.2. Rats Treated with GW2580 Have Enlarged but Reduced Number of Penetrated Perivascular Cuffs in EAE
3.3. Blocking CSF1R Decreased Cerebellum Glial Activation
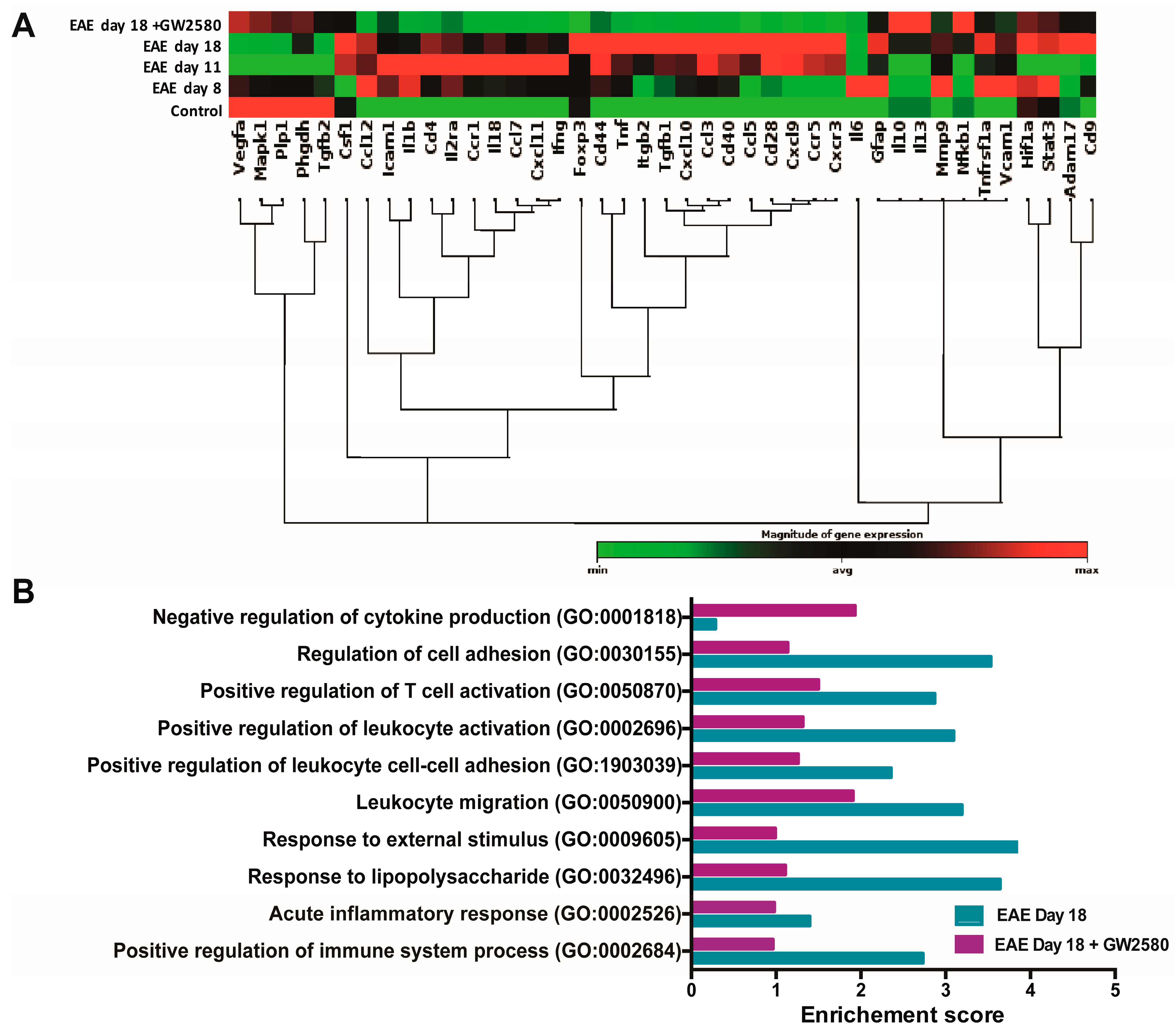
3.4. Altered Expression of BBB-Related Genes in Spinal Cord During Early EAE
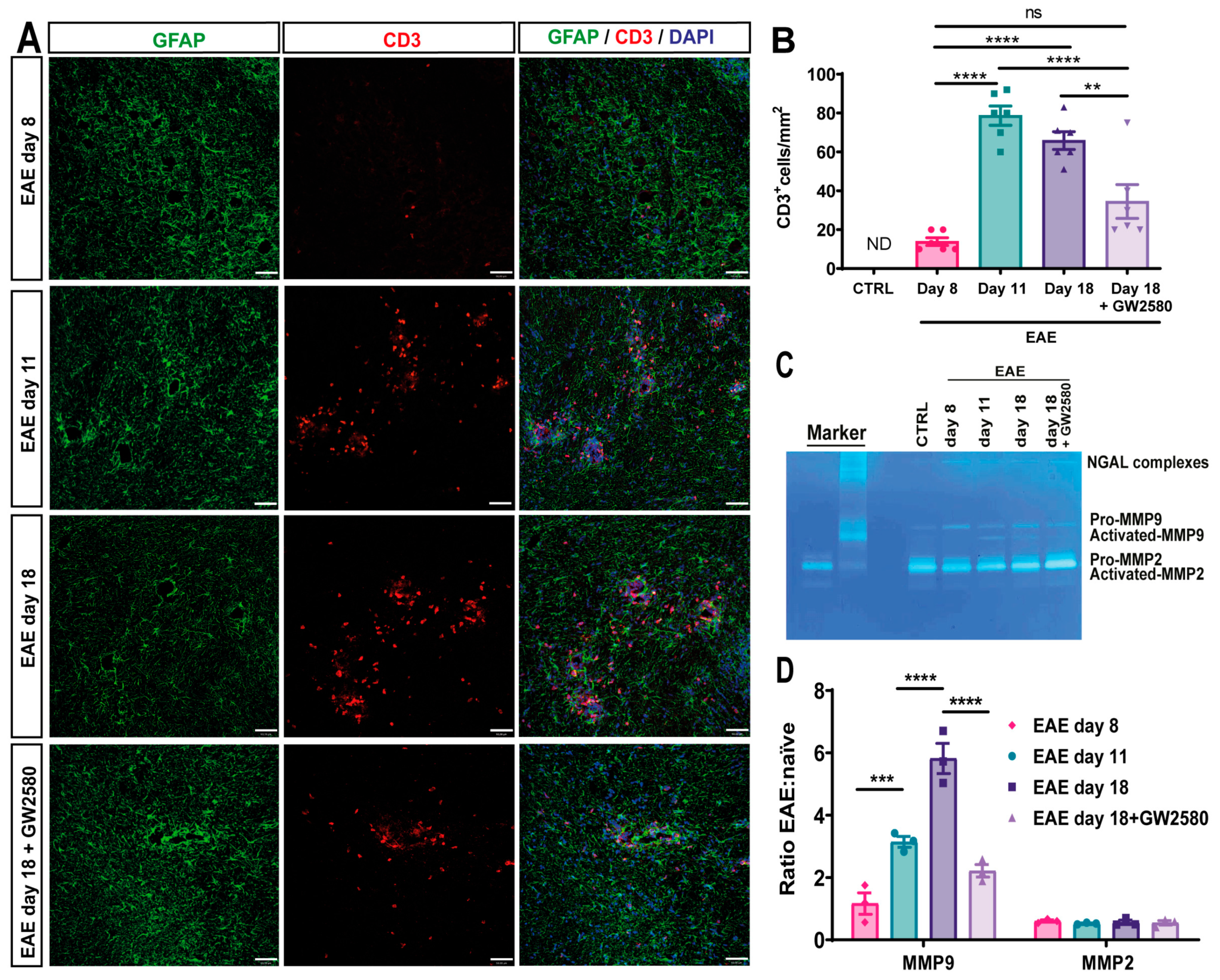
3.5. GW2580 Attenuates T-Cell Infiltration into EAE
3.6. GW2580 Treatment Correlates with Reduced MMP-2/9 Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lassmann, H.; Brück, W.; Lucchinetti, C. Heterogeneity of Multiple Sclerosis Pathogenesis: Implications for Diagnosis and Therapy. Trends Mol. Med. 2001, 7, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Kermode, A.G.; Thompson, A.J.; Tofts, P.; MacManus, D.G.; Kendall, B.E.; Kingsley, D.P.; Moseley, I.F.; Rudge, P.; McDonald, W.I. Breakdown of the Blood-Brain Barrier Precedes Symptoms and Other MRI Signs of New Lesions in Multiple Sclerosis. Pathogenetic and Clinical Implications. Brain J. Neurol. 1990, 113 Pt 5, 1477–1489. [Google Scholar] [CrossRef] [PubMed]
- Gay, D.; Esiri, M. Blood-Brain Barrier Damage in Acute Multiple Sclerosis Plaques. An Immunocytological Study. Brain J. Neurol. 1991, 114 Pt 1B, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Gay, F.W.; Drye, T.J.; Dick, G.W.; Esiri, M.M. The Application of Multifactorial Cluster Analysis in the Staging of Plaques in Early Multiple Sclerosis. Identification and Characterization of the Primary Demyelinating Lesion. Brain J. Neurol. 1997, 120 Pt 8, 1461–1483. [Google Scholar] [CrossRef]
- Trapp, B.D.; Peterson, J.; Ransohoff, R.M.; Rudick, R.; Mörk, S.; Bö, L. Axonal Transection in the Lesions of Multiple Sclerosis. N. Engl. J. Med. 1998, 338, 278–285. [Google Scholar] [CrossRef]
- McConnell, H.L.; Kersch, C.N.; Woltjer, R.L.; Neuwelt, E.A. The Translational Significance of the Neurovascular Unit. J. Biol. Chem. 2017, 292, 762–770. [Google Scholar] [CrossRef]
- Thurgur, H.; Pinteaux, E. Microglia in the Neurovascular Unit: Blood–Brain Barrier–Microglia Interactions After Central Nervous System Disorders. Neuroscience 2019, 405, 55–67. [Google Scholar] [CrossRef]
- Agrawal, S.; Anderson, P.; Durbeej, M.; van Rooijen, N.; Ivars, F.; Opdenakker, G.; Sorokin, L.M. Dystroglycan Is Selectively Cleaved at the Parenchymal Basement Membrane at Sites of Leukocyte Extravasation in Experimental Autoimmune Encephalomyelitis. J. Exp. Med. 2006, 203, 1007–1019. [Google Scholar] [CrossRef]
- Lakhan, S.E.; Kirchgessner, A.; Tepper, D.; Leonard, A. Matrix Metalloproteinases and Blood-Brain Barrier Disruption in Acute Ischemic Stroke. Front. Neurol. 2013, 4. [Google Scholar] [CrossRef]
- Song, J.; Wu, C.; Korpos, E.; Zhang, X.; Agrawal, S.M.; Wang, Y.; Faber, C.; Schäfers, M.; Körner, H.; Opdenakker, G.; et al. Focal MMP-2 and MMP-9 Activity at the Blood-Brain Barrier Promotes Chemokine-Induced Leukocyte Migration. Cell Rep. 2015, 10, 1040–1054. [Google Scholar] [CrossRef]
- Zhang, S.; Kan, Q.-C.; Xu, Y.; Zhang, G.-X.; Zhu, L. Inhibitory Effect of Matrine on Blood-Brain Barrier Disruption for the Treatment of Experimental Autoimmune Encephalomyelitis. Mediators Inflamm. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular Matrix Degradation and Remodeling in Development and Disease. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- McQuibban, G.A.; Butler, G.S.; Gong, J.H.; Bendall, L.; Power, C.; Clark-Lewis, I.; Overall, C.M. Matrix Metalloproteinase Activity Inactivates the CXC Chemokine Stromal Cell-Derived Factor-1. J. Biol. Chem. 2001, 276, 43503–43508. [Google Scholar] [CrossRef]
- Tietz, S.; Engelhardt, B. Brain Barriers: Crosstalk between Complex Tight Junctions and Adherens Junctions. J Cell Biol 2015, 209, 493–506. [Google Scholar] [CrossRef]
- Hannocks, M.-J.; Zhang, X.; Gerwien, H.; Chashchina, A.; Burmeister, M.; Korpos, E.; Song, J.; Sorokin, L. The Gelatinases, MMP-2 and MMP-9, as Fine Tuners of Neuroinflammatory Processes. Matrix Biol. 2019, 75–76, 102–113. [Google Scholar] [CrossRef]
- Császár, E.; Lénárt, N.; Cserép, C.; Környei, Z.; Fekete, R.; Pósfai, B.; Balázsfi, D.; Hangya, B.; Schwarcz, A.D.; Szabadits, E.; et al. Microglia Modulate Blood Flow, Neurovascular Coupling, and Hypoperfusion via Purinergic Actions. J. Exp. Med. 2022, 219, e20211071. [Google Scholar] [CrossRef]
- Cserép, C.; Pósfai, B.; Lénárt, N.; Fekete, R.; László, Z.I.; Lele, Z.; Orsolits, B.; Molnár, G.; Heindl, S.; Schwarcz, A.D.; et al. Microglia Monitor and Protect Neuronal Function through Specialized Somatic Purinergic Junctions. Science 2020, 367, 528–537. [Google Scholar] [CrossRef]
- Joost, E.; Jordão, M.J.C.; Mages, B.; Prinz, M.; Bechmann, I.; Krueger, M. Microglia Contribute to the Glia Limitans around Arteries, Capillaries and Veins under Physiological Conditions, in a Model of Neuroinflammation and in Human Brain Tissue. Brain Struct. Funct. 2019, 224, 1301–1314. [Google Scholar] [CrossRef]
- Salamero-Boix, A.; Schulz, M.; Sevenich, L. Activation of Microglia and Macrophages in Neurodegenerative Diseases. In Neurodegenerative Diseases Biomarkers: Towards Translating Research to Clinical Practice; Peplow, P.V., Martinez, B., Gennarelli, T.A., Eds.; Neuromethods; Springer: New York, NY, USA, 2022; pp. 11–38. ISBN 978-1-07-161712-0. [Google Scholar]
- Sosna, J.; Philipp, S.; Albay, R.; Reyes-Ruiz, J.M.; Baglietto-Vargas, D.; LaFerla, F.M.; Glabe, C.G. Early Long-Term Administration of the CSF1R Inhibitor PLX3397 Ablates Microglia and Reduces Accumulation of Intraneuronal Amyloid, Neuritic Plaque Deposition and Pre-Fibrillar Oligomers in 5XFAD Mouse Model of Alzheimer’s Disease. Mol. Neurodegener. 2018, 13, 11. [Google Scholar] [CrossRef]
- Spangenberg, E.E.; Lee, R.J.; Najafi, A.R.; Rice, R.A.; Elmore, M.R.P.; Blurton-Jones, M.; West, B.L.; Green, K.N. Eliminating Microglia in Alzheimer’s Mice Prevents Neuronal Loss without Modulating Amyloid-β Pathology. Brain J. Neurol. 2016, 139, 1265–1281. [Google Scholar] [CrossRef]
- Tarale, P.; Alam, M.M. Colony-Stimulating Factor 1 Receptor Signaling in the Central Nervous System and the Potential of Its Pharmacological Inhibitors to Halt the Progression of Neurological Disorders. Inflammopharmacology 2022. [Google Scholar] [CrossRef] [PubMed]
- Nissen, J.C.; Thompson, K.K.; West, B.L.; Tsirka, S.E. Csf1R Inhibition Attenuates Experimental Autoimmune Encephalomyelitis and Promotes Recovery. Exp. Neurol. 2018, 307, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ren, H.; Wood, K.; Li, M.; Qiu, S.; Shi, F.-D.; Ma, C.; Liu, Q. Depletion of Microglia Augments the Dopaminergic Neurotoxicity of MPTP. FASEB J. 2018, 32, 3336–3345. [Google Scholar] [CrossRef] [PubMed]
- Lou, N.; Takano, T.; Pei, Y.; Xavier, A.L.; Goldman, S.A.; Nedergaard, M. Purinergic Receptor P2RY12-Dependent Microglial Closure of the Injured Blood–Brain Barrier. Proc. Natl. Acad. Sci. USA 2016, 113, 1074–1079. [Google Scholar] [CrossRef]
- Conway, J.G.; McDonald, B.; Parham, J.; Keith, B.; Rusnak, D.W.; Shaw, E.; Jansen, M.; Lin, P.; Payne, A.; Crosby, R.M.; et al. Inhibition of Colony-Stimulating-Factor-1 Signaling in Vivo with the Orally Bioavailable cFMS Kinase Inhibitor GW2580. Proc. Natl. Acad. Sci. USA 2005, 102, 16078–16083. [Google Scholar] [CrossRef]
- Neal, M.L.; Fleming, S.M.; Budge, K.M.; Boyle, A.M.; Kim, C.; Alam, G.; Beier, E.E.; Wu, L.-J.; Richardson, J.R. Pharmacological Inhibition of CSF1R by GW2580 Reduces Microglial Proliferation and Is Protective against Neuroinflammation and Dopaminergic Neurodegeneration. FASEB J. 2020, 34, 1679–1694. [Google Scholar] [CrossRef]
- Gómez-Nicola, D.; Fransen, N.L.; Suzzi, S.; Perry, V.H. Regulation of Microglial Proliferation during Chronic Neurodegeneration. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 2481–2493. [Google Scholar] [CrossRef]
- De Lucia, C.; Rinchon, A.; Olmos-Alonso, A.; Riecken, K.; Fehse, B.; Boche, D.; Perry, V.H.; Gomez-Nicola, D. Microglia Regulate Hippocampal Neurogenesis during Chronic Neurodegeneration. Brain. Behav. Immun. 2016, 55, 179–190. [Google Scholar] [CrossRef]
- Martínez-Muriana, A.; Mancuso, R.; Francos-Quijorna, I.; Olmos-Alonso, A.; Osta, R.; Perry, V.H.; Navarro, X.; Gomez-Nicola, D.; López-Vales, R. CSF1R Blockade Slows the Progression of Amyotrophic Lateral Sclerosis by Reducing Microgliosis and Invasion of Macrophages into Peripheral Nerves. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Olmos-Alonso, A.; Schetters, S.T.T.; Sri, S.; Askew, K.; Mancuso, R.; Vargas-Caballero, M.; Holscher, C.; Perry, V.H.; Gomez-Nicola, D. Pharmacological Targeting of CSF1R Inhibits Microglial Proliferation and Prevents the Progression of Alzheimer’s-like Pathology. Brain 2016, 139, 891–907. [Google Scholar] [CrossRef]
- Crespo, O.; Kang, S.C.; Daneman, R.; Lindstrom, T.M.; Ho, P.P.; Sobel, R.A.; Steinman, L.; Robinson, W.H. Tyrosine Kinase Inhibitors Ameliorate Autoimmune Encephalomyelitis in a Mouse Model of Multiple Sclerosis. J. Clin. Immunol. 2011, 31, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Borjini, N.; Fernández, M.; Giardino, L.; Calzà, L. Cytokine and Chemokine Alterations in Tissue, CSF, and Plasma in Early Presymptomatic Phase of Experimental Allergic Encephalomyelitis (EAE), in a Rat Model of Multiple Sclerosis. J. Neuroinflammation 2016, 13, 291. [Google Scholar] [CrossRef] [PubMed]
- Louhimies, S. Directive 86/609/EEC on the Protection of Animals Used for Experimental and Other Scientific Purposes. Altern. Lab. Anim. ATLA 2002, 30 (Suppl. 2), 217–219. [Google Scholar] [CrossRef]
- Leblond, A.-L.; Klinkert, K.; Martin, K.; Turner, E.C.; Kumar, A.H.; Browne, T.; Caplice, N.M. Systemic and Cardiac Depletion of M2 Macrophage through CSF-1R Signaling Inhibition Alters Cardiac Function Post Myocardial Infarction. PLoS ONE 2015, 10, e0137515. [Google Scholar] [CrossRef]
- Wang, W.; Sun, W.; Gao, X.; Peng, L.; Lin, L.; Xiao, K.; Liu, Y.; Di, X.; Zhu, S.; Chen, H.; et al. The Preventive Effects of Colony-Stimulating Factor 1 Receptor (CSF-1R) Inhibition on Bladder Outlet Obstruction Induced Remodeling. Neurourol. Urodyn. 2022, 41, 787–796. [Google Scholar] [CrossRef]
- Calza, L.; Fernandez, M.; Giuliani, A.; Aloe, L.; Giardino, L. Thyroid Hormone Activates Oligodendrocyte Precursors and Increases a Myelin-Forming Protein and NGF Content in the Spinal Cord during Experimental Allergic Encephalomyelitis. Proc. Natl. Acad. Sci. USA 2002, 99, 3258–3263. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Sixt, M.; Engelhardt, B.; Pausch, F.; Hallmann, R.; Wendler, O.; Sorokin, L.M. Endothelial Cell Laminin Isoforms, Laminins 8 and 10, Play Decisive Roles in T Cell Recruitment across the Blood-Brain Barrier in Experimental Autoimmune Encephalomyelitis. J. Cell Biol. 2001, 153, 933–946. [Google Scholar] [CrossRef]
- Agrawal, S.M.; Williamson, J.; Sharma, R.; Kebir, H.; Patel, K.; Prat, A.; Yong, V.W. Extracellular Matrix Metalloproteinase Inducer Shows Active Perivascular Cuffs in Multiple Sclerosis. Brain 2013, 136, 1760–1777. [Google Scholar] [CrossRef]
- Wu, C.; Ivars, F.; Anderson, P.; Hallmann, R.; Vestweber, D.; Nilsson, P.; Robenek, H.; Tryggvason, K.; Song, J.; Korpos, E.; et al. Endothelial Basement Membrane Laminin Alpha5 Selectively Inhibits T Lymphocyte Extravasation into the Brain. Nat. Med. 2009, 15, 519–527. [Google Scholar] [CrossRef]
- Compston, A.; Coles, A. Multiple Sclerosis. Lancet Lond. Engl. 2002, 359, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Frohman, E.M.; Racke, M.K.; Raine, C.S. Multiple Sclerosis--the Plaque and Its Pathogenesis. N. Engl. J. Med. 2006, 354, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H.; van Horssen, J.; Mahad, D. Progressive Multiple Sclerosis: Pathology and Pathogenesis. Nat. Rev. Neurol. 2012, 8, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Trapp, B.D.; Nave, K.-A. Multiple Sclerosis: An Immune or Neurodegenerative Disorder? Annu. Rev. Neurosci. 2008, 31, 247–269. [Google Scholar] [CrossRef]
- Zigmond, M.J.; Coyle, J.T.; Rowland, L.P. Neurobiology of Brain Disorders: Biological Basis of Neurological and Psychiatric Disorders; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 978-0-12-398280-3. [Google Scholar]
- Erblich, B.; Zhu, L.; Etgen, A.M.; Dobrenis, K.; Pollard, J.W. Absence of Colony Stimulation Factor-1 Receptor Results in Loss of Microglia, Disrupted Brain Development and Olfactory Deficits. PLoS ONE 2011, 6, e26317. [Google Scholar] [CrossRef]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate Mapping Analysis Reveals That Adult Microglia Derive from Primitive Macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef]
- Irvine, K.M.; Burns, C.J.; Wilks, A.F.; Su, S.; Hume, D.A.; Sweet, M.J. A CSF-1 Receptor Kinase Inhibitor Targets Effector Functions and Inhibits pro-Inflammatory Cytokine Production from Murine Macrophage Populations. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2006, 20, 1921–1923. [Google Scholar] [CrossRef]
- Adzemovic, M.V.; Adzemovic, M.Z.; Zeitelhofer, M.; Eriksson, U.; Olsson, T.; Nilsson, I. Imatinib Ameliorates Neuroinflammation in a Rat Model of Multiple Sclerosis by Enhancing Blood-Brain Barrier Integrity and by Modulating the Peripheral Immune Response. PLoS ONE 2013, 8, e56586. [Google Scholar] [CrossRef]
- Engelhardt, B.; Ransohoff, R.M. The Ins and Outs of T-Lymphocyte Trafficking to the CNS: Anatomical Sites and Molecular Mechanisms. Trends Immunol. 2005, 26, 485–495. [Google Scholar] [CrossRef]
- Sorokin, L. The Impact of the Extracellular Matrix on Inflammation. Nat. Rev. Immunol. 2010, 10, 712–723. [Google Scholar] [CrossRef]
- Larochelle, C.; Alvarez, J.I.; Prat, A. How Do Immune Cells Overcome the Blood–Brain Barrier in Multiple Sclerosis? FEBS Lett. 2011, 585, 3770–3780. [Google Scholar] [CrossRef] [PubMed]
- Alt, C.; Laschinger, M.; Engelhardt, B. Functional Expression of the Lymphoid Chemokines CCL19 (ELC) and CCL 21 (SLC) at the Blood-Brain Barrier Suggests Their Involvement in G-Protein-Dependent Lymphocyte Recruitment into the Central Nervous System during Experimental Autoimmune Encephalomyelitis. Eur. J. Immunol. 2002, 32, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Dorovini-Zis, K. The Blood-Brain Barrier in Health and Disease, Volume One: Morphology, Biology and Immune Function; CRC Press: Boca Raton, FL, USA, 2015; ISBN 978-1-4987-2706-8. [Google Scholar]
- Neuwelt, E.A.; Bauer, B.; Fahlke, C.; Fricker, G.; Iadecola, C.; Janigro, D.; Leybaert, L.; Molnár, Z.; O’Donnell, M.E.; Povlishock, J.T.; et al. Engaging Neuroscience to Advance Translational Research in Brain Barrier Biology. Nat. Rev. Neurosci. 2011, 12, 169–182. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, L.; Pu, H.; Mao, L.; Hu, X.; Jiang, X.; Xu, N.; Stetler, R.A.; Zhang, F.; Liu, X.; et al. Rapid Endothelial Cytoskeletal Reorganization Enables Early Blood–Brain Barrier Disruption and Long-Term Ischaemic Reperfusion Brain Injury. Nat. Commun. 2016, 7, 10523. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Joh, T.H. Matrix Metalloproteinases, New Insights into the Understanding of Neurodegenerative Disorders. Biomol. Ther. 2012, 20, 133–143. [Google Scholar] [CrossRef]
- Van den Steen, P.E.; Wuyts, A.; Husson, S.J.; Proost, P.; Van Damme, J.; Opdenakker, G. Gelatinase B/MMP-9 and Neutrophil Collagenase/MMP-8 Process the Chemokines Human GCP-2/CXCL6, ENA-78/CXCL5 and Mouse GCP-2/LIX and Modulate Their Physiological Activities. Eur. J. Biochem. 2003, 270, 3739–3749. [Google Scholar] [CrossRef]
- Van den Steen, P.E.; Proost, P.; Wuyts, A.; Van Damme, J.; Opdenakker, G. Neutrophil Gelatinase B Potentiates Interleukin-8 Tenfold by Aminoterminal Processing, Whereas It Degrades CTAP-III, PF-4, and GRO-Alpha and Leaves RANTES and MCP-2 Intact. Blood 2000, 96, 2673–2681. [Google Scholar] [CrossRef]
- Conway, J.G.; Pink, H.; Bergquist, M.L.; Han, B.; Depee, S.; Tadepalli, S.; Lin, P.; Crumrine, R.C.; Binz, J.; Clark, R.L.; et al. Effects of the cFMS Kinase Inhibitor 5-(3-Methoxy-4-((4-Methoxybenzyl)Oxy)Benzyl)Pyrimidine-2,4-Diamine (GW2580) in Normal and Arthritic Rats. J. Pharmacol. Exp. Ther. 2008, 326, 41–50. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borjini, N.; Fernandez, M.; Giardino, L.; Sorokin, L.; Calzà, L. Pharmacological Inhibition of Microglial Proliferation Supports Blood–Brain Barrier Integrity in Experimental Autoimmune Encephalomyelitis. Cells 2025, 14, 414. https://doi.org/10.3390/cells14060414
Borjini N, Fernandez M, Giardino L, Sorokin L, Calzà L. Pharmacological Inhibition of Microglial Proliferation Supports Blood–Brain Barrier Integrity in Experimental Autoimmune Encephalomyelitis. Cells. 2025; 14(6):414. https://doi.org/10.3390/cells14060414
Chicago/Turabian StyleBorjini, Nozha, Mercedes Fernandez, Luciana Giardino, Lydia Sorokin, and Laura Calzà. 2025. "Pharmacological Inhibition of Microglial Proliferation Supports Blood–Brain Barrier Integrity in Experimental Autoimmune Encephalomyelitis" Cells 14, no. 6: 414. https://doi.org/10.3390/cells14060414
APA StyleBorjini, N., Fernandez, M., Giardino, L., Sorokin, L., & Calzà, L. (2025). Pharmacological Inhibition of Microglial Proliferation Supports Blood–Brain Barrier Integrity in Experimental Autoimmune Encephalomyelitis. Cells, 14(6), 414. https://doi.org/10.3390/cells14060414