1. Introduction
Irradiation of the ocular globe is the first line treatment in patients who are suffering from a choroidal melanoma. Since these are small tumor findings, only a few millimeters in size, a focused radiation with minimizing damage to surrounding tissues is desirable.
In everyday clinical practice, plaque brachytherapy with, for example, Ruthenium-106 (beta radiation), Iodine-125 and Palladium-103 (both photon radiation) plaques and, where appropriate facilities are available, external beam radiotherapy with charged particles such as protons have become established [
1]. A targeted irradiation of the choroidal melanoma with proton beams allows an eye-preserving therapy [
2,
3,
4]. However, there is still a risk of developing a radiation retinopathy after ocular radiation, which can lead to a loss of function and, in rare cases, as well to an enucleation [
5]. Radiation retinopathy has a negative impact on the visual outcome of proton irradiation of choroidal melanoma [
6,
7].
In the course of clinical observations as well as studies, it was revealed that radiation retinopathy is a slowly progressive microangiopathy of the retinal vessels that becomes evident after a delay of approximately 2 years after proton irradiation [
8]. Various vascular pathologies can be observed, including ischemic or proliferative retinopathy and maculopathy [
1,
9,
10]. Based on these observations, there could be pathophysiological similarities to diabetic retinopathy, which is characterized by a microangiopathy, leading to progressive capillary nonperfusion [
11], but also an inflammatory component.
Central to the development of Radiation retinopathy is the dysfunction of endothelial cells, which line the interior surfaces of blood vessels and are pivotal in maintaining vascular integrity and the blood-retinal barrier. Radiation exposure disrupts these cells, leading to increased vascular permeability, loss of cell-cell contact, and subsequent cell death. These alterations can precipitate a cascade of pathophysiological events in the retina, including edema and neovascularization, which are characteristic of radiation retinopathy [
12].
The damage to endothelial cells from radiation is mediated primarily through oxidative stress and an inflammatory response. Oxidative stress, induced by radiation, results in the production of reactive oxygen species (ROS) that can damage cellular proteins, lipids, and DNA. Concurrently, radiation can trigger a robust inflammatory response characterized by the upregulation of cytokines and adhesion molecules, fostering an environment that facilitates leukocyte adhesion and migration into retinal tissues. These processes exacerbate endothelial injury and contribute to the vascular anomalies observed in radiation retinopathy [
13,
14].
Insights into these mechanisms have been greatly advanced by studies utilizing mouse models, which have been instrumental in elucidating the cellular and molecular underpinnings of radiation retinopathy. These models have highlighted significant changes in retinal vascular architecture following radiation exposure, including vessel dilation, tortuosity, and the formation of microaneurysms. Furthermore, mouse models have served as critical platforms for testing the efficacy of therapeutic interventions, such as anti-VEGF therapies, which aim to mitigate endothelial damage and preserve retinal function [
15].
Parallel clinical observations in humans have detailed a spectrum of microvascular abnormalities in radiation retinopathy, ranging from cotton wool spots to severe hemorrhages and neovascularization. The management of these clinical manifestations primarily involves the administration of anti-VEGF injections, corticosteroids, and in some cases, laser photocoagulation, highlighting the need for continued research into more effective therapeutic strategies [
16].
Recent advancements in this field are focusing on identifying early biomarkers for radiation retinopathy and developing novel protective agents that can shield endothelial cells from radiation-induced harm.
To date, there are no experimental studies using proton irradiation on mouse eyes. But in experiments with external beam radiation from rat eyes, including protons, endothelial cell loss, capillary dropout, vessel occlusion, retinal smooth muscle loss, retinal degradation with photoreceptor loss were observed [
1].
Most animal model studies focus on the vasculopathy of radiation retinopathy [
1]. Less is known about inflammatory mechanisms, but they also appear to play a significant role. Focusing on the inflammatory components of radiation retinopathy, such as microglia activation, and the roles of glial fibrillary acidic protein (GFAP), vimentin, and pericytes, is now increasingly important. While much of the research has traditionally concentrated on endothelial cell dysfunction, the broader inflammatory response plays a crucial role in the progression and severity of the disease. Microglia, the resident immune cells of the central nervous system, become activated in response to radiation and can exacerbate damage through the release of pro-inflammatory cytokines and reactive oxygen species. Similarly, upregulation of GFAP and vimentin in astrocytes and Müller cells, respectively, indicates reactive gliosis, which contributes to the inflammatory environment and structural disorganization of the retina. Pericytes, vital for capillary stability, undergo changes that further disrupt vascular integrity and blood-retinal barrier function. By shifting the focus toward these elements, researchers can uncover new therapeutic targets that address the complex network of cellular interactions and inflammatory pathways involved in radiation retinopathy, potentially leading to more effective treatments that mitigate the broader spectrum of pathological changes beyond just the endothelial cells. In a publication on radiation retinopathy in rat eyes following irradiation with X-rays, they showed an invasion of microglia and macrophages into the retinal pigment epithelium and that this observation is related to an outer blood retina barrier disruption [
17].
The aim of this study, therefore, was to establish a model of radiation retinopathy following proton beam irradiation (as in everyday clinical practice with humans) in mice and to investigate irradiation-related morphological and functional consequences in this model focusing on the inflammatory components such as microglia. Therefore, transgenic Cx3cr1 mice were included in the experiments. The homozygote Cx3cr1
gfp/gfp mice lack the fractalkine receptor, which is mainly expressed by microglia in the brain and retina and has an important role in trafficking microglia cells to and from the subretinal space [
18].
A possible effect of the altered microglia cell migration on the development of radiation retinopathy could be a new therapeutic approach in preserving visual function after proton irradiation of the ocular globe.
2. Methods
2.1. Study Approval
All animal experiments were committed in conformity with the German law of animal protection and the National Institute of Health guidelines for care and use of laboratory animals. All experiments were approved by the local ethics committee on animal research (LaGeSo, Landesamt für Gesundheit und Soziales Berlin, Germany, G0218/13). The experimental design and execution follow the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research.
2.2. Animal Model
C57/Bl6J mice were purchased from Janvier (Cedex, France) and were used to establish the irradiation protocol as well as to perform a first characterization of proton radiation-related anatomical and functional ophthalmological changes.
Subsequently, transgenic Cx3cr1
gfp/gfp mice on a C57BL/6J background were used to study the impact of Cx3cr1 signaling on the ophthalmological effects of proton radiation. In this mouse line, the first 390bp of the Cx3cr1 gene are replaced by the gene encoding the enhanced green fluorescent protein (EGFP) and therefore the homozygote mice lack the fractalkine receptor, which is mainly expressed by microglia in the brain and retina [
18,
19]. Heterozygote mice of this strain are often used as reporter mice to visualize microglia in vivo and ex vivo by EGFP fluorescence. Consequently, it is assumed that in heterozygote mice Cx3cr1 signaling is possible without restriction. However, microglia in Cx3cr1
gfp/+ mice exhibit an intermediate transcriptional profile between wild-type microglia and microglia in the Cx3cr1
gfp/gfp mice [
19,
20].
Therefore, we included this strain into our study to test whether the heterozygote mice (Cx3cr1gfp/+)compared to wild type mice (Cx3cr1+/+) and to investigate the impact of Cx3xr1 loss on the outcome in the homozygote mice (Cx3cr1gfp/gfp). The Cx3cr1 strain was kindly provided by F. Sennlaub (Department of Therapeutics, Sorbonne Université, INSERM, CNRS, Institute de la Vision, Paris, France) and tested negative for RD8 mutation. In our local animal facility, the breeding was sustained using the C57BL/6J line. Independent of the genotype, all mice were female, 10–12 weeks of age and weighing 20 to 25 g at the beginning of the experiments to prevent bias due to inter-individual variability.
2.3. Irradiation Procedure
The proton irradiation was performed at the Helmholtz-Zentrum Berlin für Materialien und Energie (HZB) in Berlin-Wannsee. The proton facility is primarily designed and built to treat patients, who suffer from choroidal melanoma [
21,
22]. The responsible radiation physicists on site created the plan for the irradiation of the mouse eyes in a blinded fashion for the genotype. Irradiation field and aperture were adapted to the murine anatomy. The mice were anesthetized and placed onto a customized platform in front of the clinical beam line. The anesthesia was done by an intraperitoneal injection of a dose of 100 mg/kg BW Ketamine and 12 mg/kg BW Xylazine. After irradiation, the mice were brought back to their cage to wake up. One eye was irradiated while the contralateral eye and both eyes of the sham animals served as control. Sham animals were only placed in front of the clinical beam line for 5 min to control for the stress without irradiating the animals.
The data on the radiation field include the aperture diameter, which measured 9 mm, the depth of penetration into the mouse tissue (range), which was 5mm, and the length of the does plateau (modulation) measuring 5 mm. The dose rate was approximately 10.5 Gy/min or 11.6 Cobalt Gray Equivalent (CGE)/min.
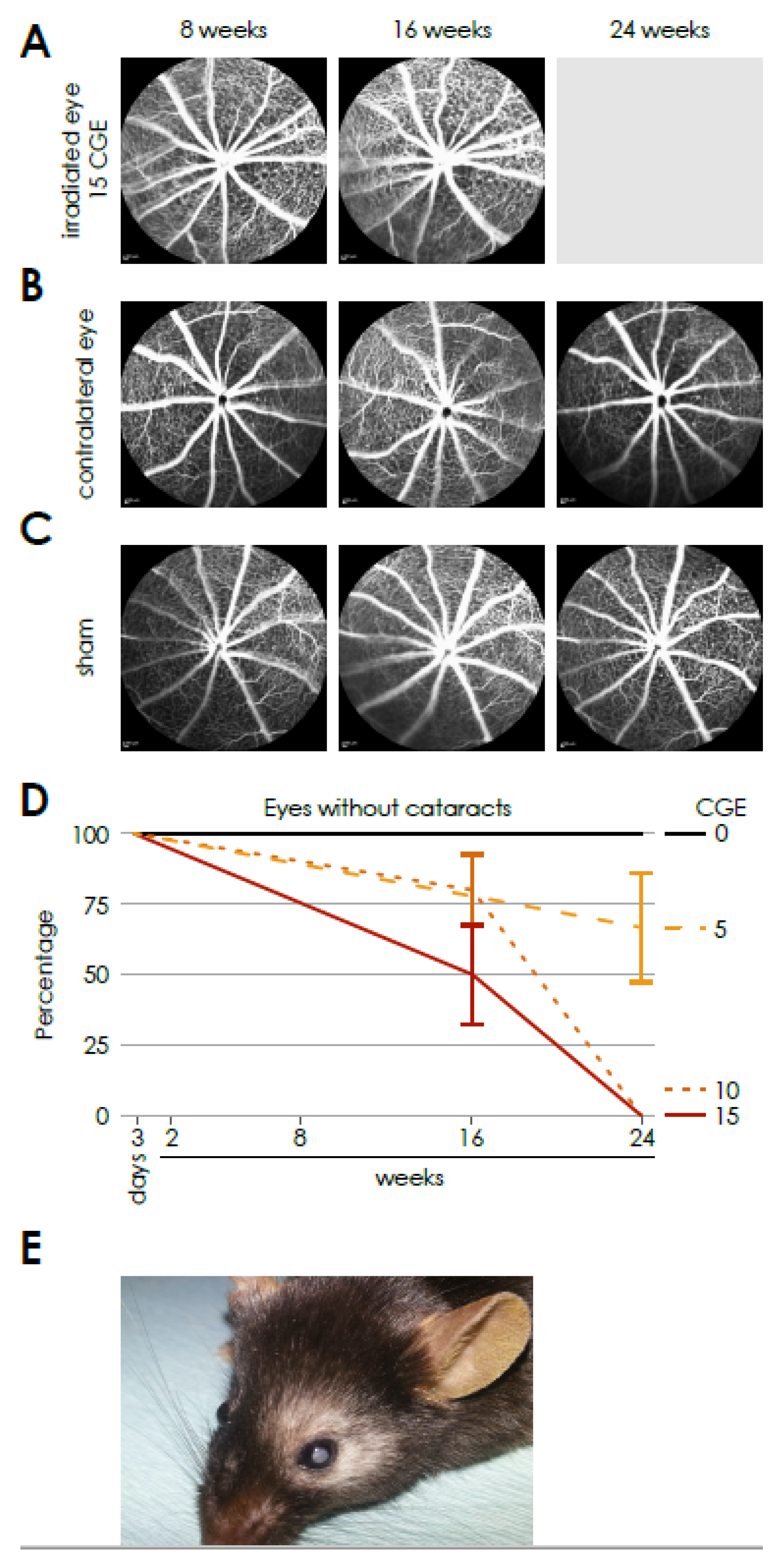
The following doses were applied for establishing a radiation dose-response curve for mice: 4.55 Gy or 5.0 CGE, 9.09 Gy or 10.0 CGE and 13.64 Gy or 15.0 CGE. The doses were determined by calculating the equivalent mouse dosages based on those used in patients. A detectable effect was observed at 13.64 Gy 15 GDE, which was subsequently chosen for further experiments.
2.4. Scanning Laser Ophthalmoscopy
The in vivo imaging of the retina of both mouse eyes was performed using a scanning laser ophthalmoscope at 8, 16 and 24 weeks after proton irradiation as described previously [
23].
Briefly, the mice were anesthetized prior to the examination. The corneas were moisturized with Hylo-Vision (NaCl 0.1%; Omnivision, Puchheim, Germany) and the mice were placed on a customized platform in front of the device. Spectralis HRA-OCT (Heidelberg Engineering, Heidelberg, Germany) was settled with a c-curve of 3.5 and incorporated a wide-angle lens 55°.
Fluorescence angiography (FA) was performed to assess the vessel structure and any leakage. For this purpose, the mouse eyes were examined with the FA mode (diode laser, wavelength of 488 nm), 5 min after the subcutaneous injection of fluorescein (5 mg/kg, fluorescein 10%; Alcon, Freiburg, Germany).
The auto-fluorescence mode in the ophthalmoscope (AF, 488 nm) was used to display the EFGP positive cells of the heterozygous Cx3cr1gfp/+ and homozygous Cx3cr1gfp/gfp mice.
Images of the inner and outer retina were taken centrally and peripherally and were analyzed using the Heidelberg Eye Explorer 1.7.0.0 software.
2.5. ERG
The retinal function was measured with the help of Ganzfeld electroretinography (ERG) as described previously [
24].
Briefly, before the measurement, the mice were adapted to the dark overnight. Then, after the mice were anesthetized with xylazine (15 mg/kg body weight) and ketamine (100 mg/kg) and the pupils dilated with 0.5% tropicamide and 2.5% phenylephrine hydrochloride, they were placed on a warm plate in a Ganzfeld bowl (Roland Consult, Brandenburg, Germany).
The ERG was recorded with monopolar contact lens electrodes. Platin needles, which were fixed subcutaneously, served as reference and ground electrodes.
The examination took place 3 days and 2, 8, 16 and 24 weeks after proton irradiation.
2.6. Immunohistochemistry
The eyes for the immunofluorescence staining were obtained 24 weeks after proton irradiation. To visualize GFAP and Vimentin to examine gliosis and fibrosis, the enucleated eyes were fixed in 4% PFA overnight and embedded in paraffin to make sections from the central bulbus with a thickness of 4 µm. After the sections were deparaffinized and rehydrated, they were incubated with Proteinase K for antigen retrieval and permeabilized in TBS with 0.5% Triton X-100. After 1 h blocking with 5% BSA, the sections were incubated overnight with the primary antibodies (guinea pig anti-GFAP, Synaptic Systems, Göttingen, Germany, 173004, 1:100, chicken anti-Vimentin, Novus Biologicals, Centennial, CO, USA, NB300-223, 1:50) and the day after with DAPI (Roche, Basel, Switzerland, 10236276001, 1:100) and the secondary antibodies (Alexa Fluor 568 goat anti-guinea pig, A11075, 1:8000, Alexa Fluor 488 donkey anti-chicken, Jackson, West Grove, PA, USA, 711-545-152, 1:4000) for one hour. Covered by glass slides, sections were visualized with an Axio Imager M2 fluorescence microscope (ZEN-Software 3.1 Blue Edition, Zeiss, Oberkochen, Germany). The 488 and 568 channel images were analyzed separately with Image j 1.53a (Wayne Rasband, National Institute of Health, USA) and the retinal area was defined excluding the photoreceptor outer segments. Finally, the integrated density, as sum of values of the pixels in the image or selection equivalent to the product of area and mean gray value, was measured of a defined retinal area excluding the photoreceptor outer segments.
To further investigate changes in the vascular component on a cellular level, we employed isolectin B4 (IB4) for general vascular labeling and NG2 for specific pericyte identification in retinal flat mounts. Isolectin B4 is a lectin isolated from the seeds of the tropical African legume Griffonia simplicifolia, which specifically binds to carbohydrate structures, primarily N-acetyld-galactosamine end groups. It is widely utilized for staining murine endothelial cells to visualize and analyze vascular structures. However, Isolectin B4 also exhibits cross-reactivity with microglia [
25]. Neuroglial antigen 2 (NG2), a chondroitin sulfate proteoglycan, is predominantly expressed on the surface of oligodendrocyte precursor cells and pericytes. In the context of vascular biology, NG2 plays a crucial role in vascular stability and angiogenesis. Antibodies against NG2 are commonly used as pericyte markers in retinal vessels, facilitating the quantification of the pericyte/endothelial cell ratio [
26,
27]. In this experimental study, the number of pericytes was analyzed in relation to the length of Isolectin B4-positive retinal vessels. We have used the Isolectin B4 staining to visualize vessels in toto with no specific focus on endothelial cells.
To count microglia, cells were labeled with Iba1 in retinal flat mounts. Iba1 (ionized calcium-binding adaptor molecule 1) is specifically expressed in microglia and macrophages and is widely used as a marker to identify and study these cell types in the retina [
28].
For this purpose, the eyes were fixed in 4% PFA for 15 min. Subsequently, the cornea was removed via a circumferential limbal incision, lens and vitreous were removed and four radial incisions were made, to achieve a flattening of the bulbus. Finally, the retina was dissected from the choroid. Retinal flat mounts were transferred into a 24-well plate and permeabilized with 5% Triton X-100 in TBS overnight. The flat mounts were incubated with Isolectin-IB4 (either GS Isolectin-B4, Alexa Fluor 488-conjugated, Invitrogen, I21411, 1:200 or Alexa Fluor 647-conjugated, Invitrogen, Carlsbad, California, USA, I32450, 1:200) to visualize the vasculature. In samples treated with the 488-conjugated isolectine, the primary antibody (rabbit anti-NG2 Chondroitin Sulfate Proteoglycan, CY3-conjugated, Sigma-Aldrich, Burlington, Massachusetts, USA, AB5320C3, 1:200) was applied overnight, after one hour blocking with 15% serum in TBS. NG2 labeled cells were counted in images from the inner and outer vessel layer using an Axio Imager M2 fluorescence microscope (ZEN-Software 3.1 Blue Edition, Zeiss, Oberkochen, Germany). Samples treated with the 647-conjugated Isolectin, were incubated with a primary antibody for Iba1 (rabbit anti-Iba1, Abcam, Cambridge, UK, ab178847, 1:200) overnight followed by 2 h incubation with a secondary antibody (Alexa Fluor 488 donkey anti-rabbit, Dianova, Hamburg, Germany, 711-545-152, 1:1000) before the samples were mounted on glass slides. To count Iba1 positive cells the covered flat mounts were scanned by using the z-stack mode at a confocal Leica SPE microscope (Leica Microsystems GmbH, Wetzlar, Germany).
2.7. FACS
Single cell suspensions were obtained from the retinas by mechanical dissociation and cell separation through a moist cell sieve with pores of 70 µm in size. Cells were labeled with CD11b Monoclonal Antibody (M1/70, PE-Cyanine7, eBioscience™, San Diego, CA, USA, 25-0112-82) and CD45 eflour 450 (30-F11, eBioscience™, San Diego, USA, 48-0451-82) for 15 min in FACS buffer (PBS and 1% FCS) on ice in the dark and immediately analyzed using a FACS Canto II (BD, Franklin Lakes, NJ, USA) and the FlowJo v10.10 software as described elsewhere [
29].
2.8. Data Analysis
The Shapiro-Wilk-Test was used to evaluate the normal distribution of the variables and the Levene Test to assess the equality of variances. In case of normally distributed variables the paired samples t-test was performed to compare the variables. One-way (one independent variable) or two-way (two independent variables) ANOVA was used when comparing more than two groups. The Bonferroni-Test was used as post hoc test. For non-normally distributed variables non-parametric tests were used: the Wilcoxon-Test for two dependent variables, the Mann-Whitney-U-Test for two independent variables, the Friedmann-Test for multiple dependent variables and the Kruskal-Wallis-Test for multiple independent variables. p-values lower than 0.05 were determined to be significant. In case of missing data, a pairwise deletion was made.
The data analysis was performed with the program SPSS Statistics (Version 23.0 for Mac, Chicago, IL, USA). For the graphical representation of the data box-and-whisker plots (whiskers represent the minimum and the maximum value of the data set) or bar charts (mean ± SEM) were used. The single data points were superimposed in both plots.
4. Discussion
Proton irradiation has become the therapy of choice for central choroidal melanoma in humans, primarily due to its ability to deliver a highly focused radiation dose while minimizing damage to surrounding tissues [
1,
31,
32,
33]. However, radiation retinopathy remains a significant long-term complication, with a reported 5-year incidence rate of 85% [
6,
7]. Like diabetic retinopathy, it is characterized by a microangiopathy, including endothelial cell loss, ischemia, bleeding, and macular edema, leading to significant visual impairment [
1,
9,
10].
Previous research has primarily focused on tumor control, with limited exploration of the pathophysiological mechanisms underlying radiation retinopathy, particularly in experimental models [
1]. Our study is the first to use a targeted proton irradiation model for mouse eyes, enabling an investigation of dose-dependent vascular and retinal changes.
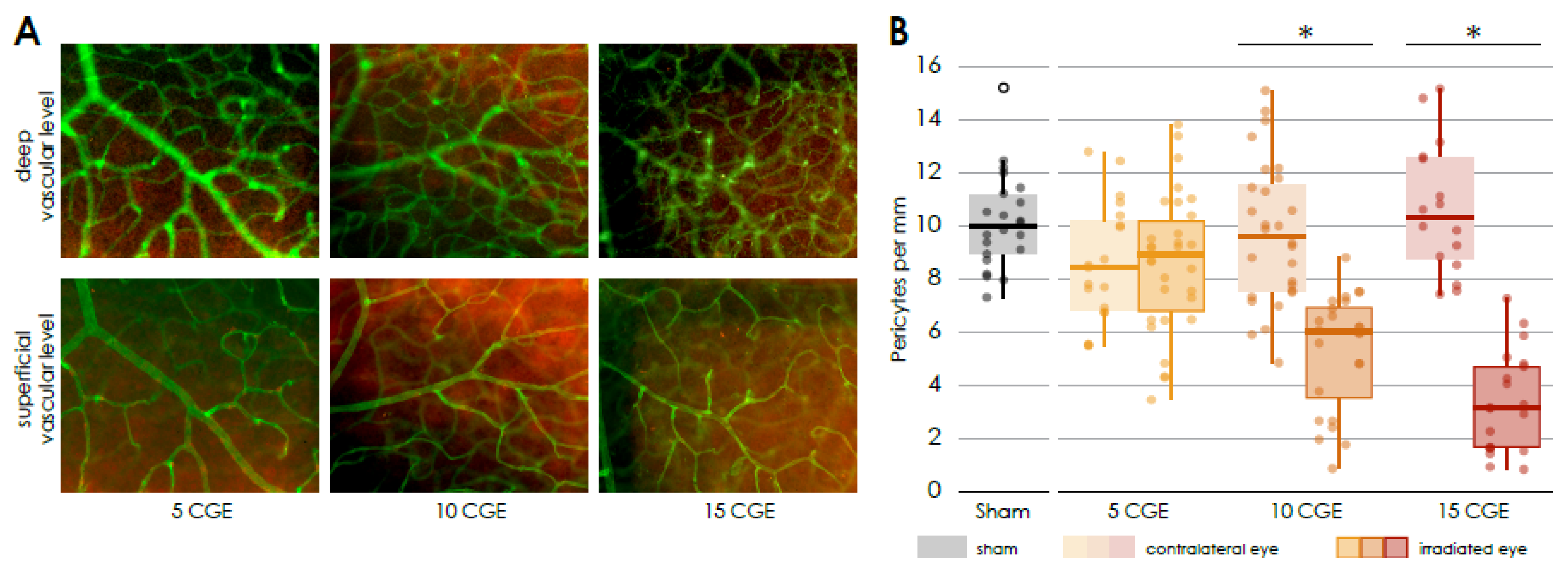
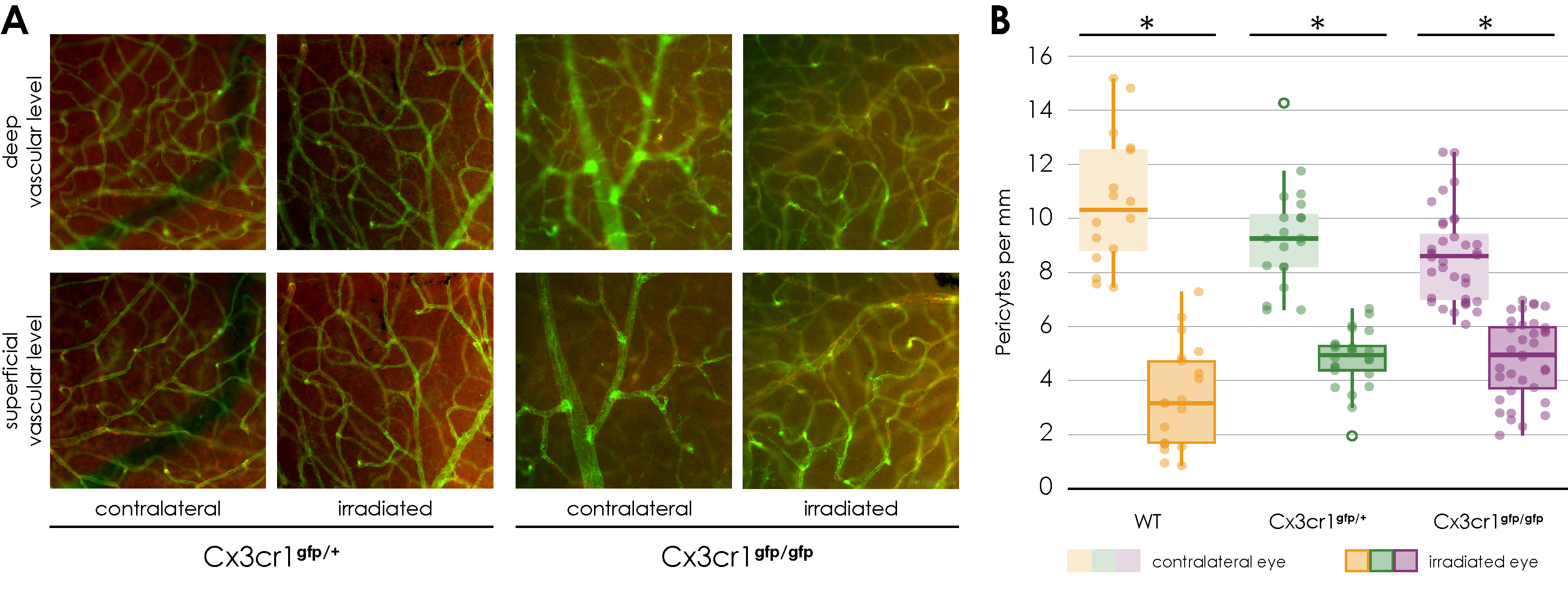
We observed a dose-dependent loss of pericytes in irradiated retinal vessels, resembling early diabetic retinopathy where pericyte loss is a hallmark [
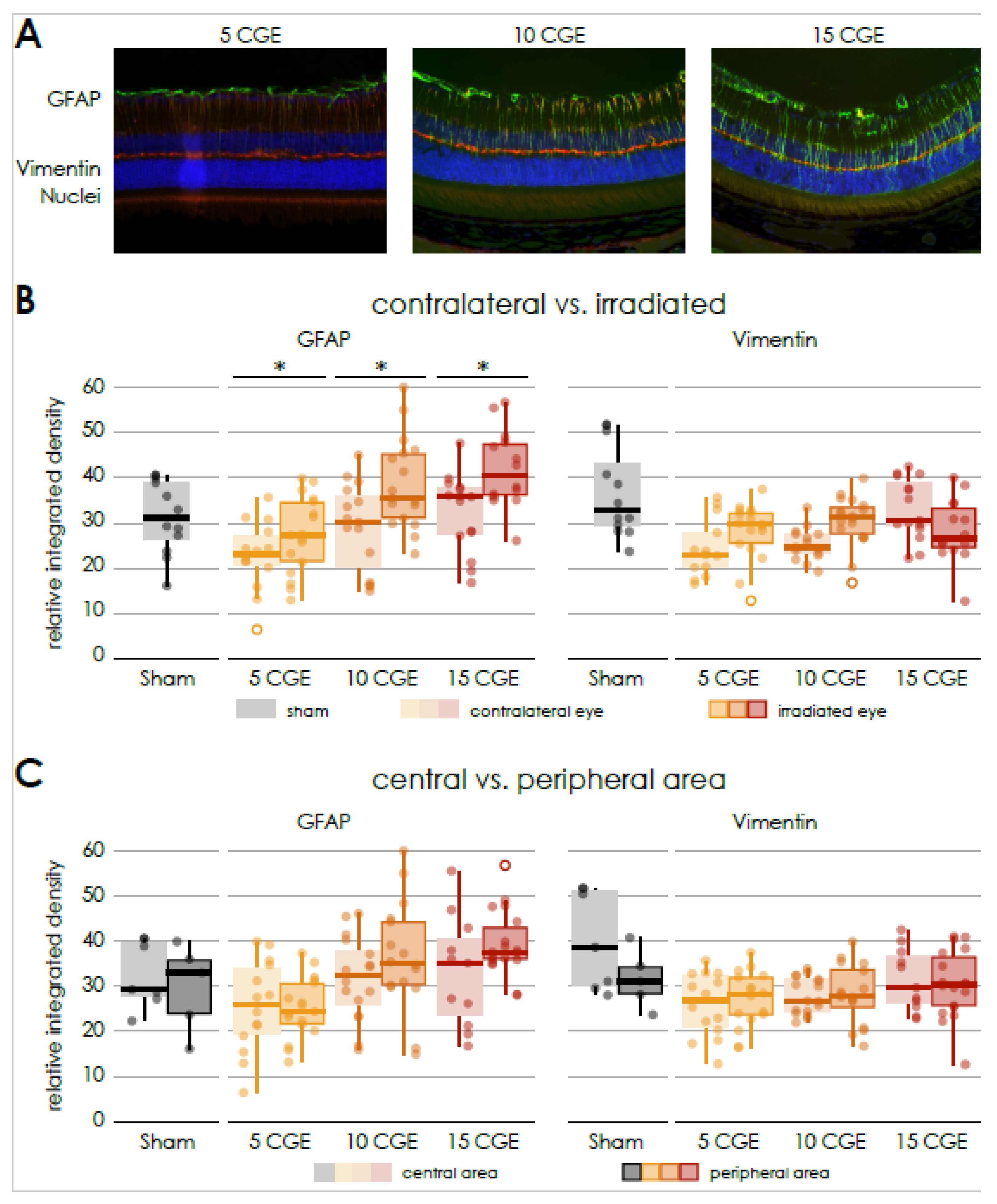
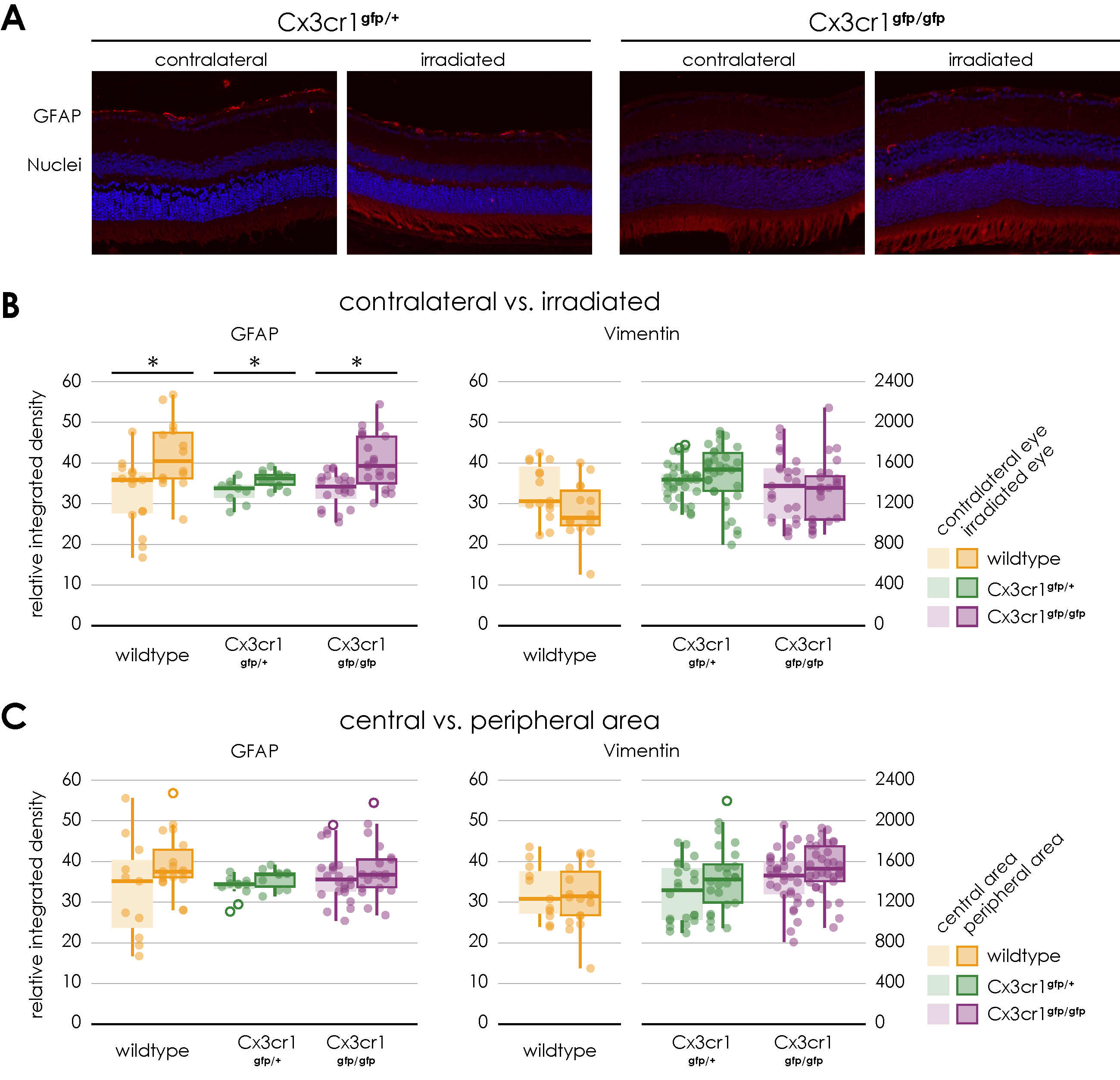
34]. Additionally, retinal gliosis, indicated by increased GFAP expression, was demonstrated in a dose-dependent manner, consistent with reactive glial activation reported in other retinal diseases [
35].
Inflammatory responses were evidenced by increased Iba1-positive microglial cells post-irradiation. Interestingly, Cx3cr1 deficiency was associated with a functional protective effect, potentially due to restricted microglial infiltration into the deep vascular layer. This finding contrasts with diabetic retinopathy models, where Cx3cr1 deficiency exacerbated inflammation and vascular damage [
36].
Functionally, retinal impairment was primarily observed in photoreceptor and bipolar cells, with significant alterations in the ERG a-wave and b-wave amplitudes at higher radiation doses. Despite the dense cataract development hindering in vivo observations at later stages, our findings provide insights into the structural and functional impact of proton irradiation on the retina and highlight the need for further mechanistic studies on the role of the Cx3cr1 pathway in mitigating radiation-induced damage.
In humans, the presence of radiation retinopathy is primarily assessed by performing fluorescence angiography of the retinal vessels. The characteristic findings are severe retinal capillary nonperfusion, capillary dilation, and microaneurysms, frequently in combination with macular edema or ischemia [
9].
Therefore, retinal fluorescence angiography was performed in the mice in week 8, 16 and 24 after proton therapy. However, no gross vessel abnormalities could be detected. The main side effect of irradiation was a dose and time-dependent development of dense cataracts. Therefore an in vivo examination of eyes at 24 weeks after irradiation with 15 CGE was impossible. Hence, at 24 weeks, changes in the sense of a radiation retinopathy could not be detected in vivo, but also cannot be excluded, at least for the 15 CGE group. Thus, in vivo, we were able to perform FAG and subsequent vessel analysis only prior to cataract formation. While a few samples were collected despite cataract formation and prepared as flat mounts, vessel abnormalities were observed. However, our focus was on investigating changes before cataract formation to align these findings with the in vivo FAG data. We acknowledge that, in a follow-up study, it would be valuable to examine eyes that developed cataracts, but this was not the primary objective of the current study.
A key finding of previous publications on the pathophysiology of radiation retinopathy is a microangiopathy that seems to be similar to diabetic retinopathy [
9].
The loss of endothelial cells and associated pericytes have already been described as primary vascular event in experiments with other forms of ionizing radiation and other species such as rats [
10,
37].
In the context of this experimental study, we showed for the first time a dose-dependent loss of NG2 positive pericytes surrounding Isolectin B4 positive endothelial cells of retinal vessels of the irradiated mouse eyes. This dose-dependent loss of pericytes seems to precede an endothelial loss, showing a similarity to early diabetic retinopathy, where pericyte loss is considered as main feature [
34]. This stands in contrast to the publication by Gardiner et al. [
11], who reported that in the context of radiation retinopathy, there is a loss of microvascular endothelial cells in the presence of surviving pericytes, as demonstrated through vascular digests obtained from both clinical radiation retinopathy and animal models. In this experimental study, Isolectin B4 was utilized to label endothelial cells for the visualization and analysis of vascular architecture, though it does not provide direct information regarding cell viability. Consequently, we can only conclude that, in comparison to the length of Isolectin B4-positive vessels, there is a dose-dependent loss of pericytes. However, we cannot ascertain the impact of radiation on the viability of endothelial cells based on our data. Additionally, the experimental designs differ; we used proton radiation as opposed to photon radiation, utilized a different animal model, and applied different radiation intensities. It would be valuable to further investigate how these differing irradiation modalities might influence the results regarding pericyte and endothelial cell interaction.
Possible degenerative processes were examined by the means of an immunofluorescence staining of GFAP and vimentin to show activated retinal glial cells as a non-specific neuropathological reaction. An increasing astrocyte and Mueller cell reactive gliosis, evident by upregulation of GFAP expression, could also be demonstrated in a dose-dependent manner. No increased expression of GFAP was reported in the central retina 3 months after irradiation in the context of a mouse model of chronic glaucoma [
35]. However, since the type and dose of irradiation as well as the time frame differ from our experimental setting, a direct comparison is not possible.
Since the severity of the phenotype was more evident in the 15 CGE group, the further experiments were carried out with this radiation dose. And since the results of the non-irradiated eyes correspond to those of the sham eyes, the sham group was dispensed in the subsequent experiments.
While in X-radiation (2–20 Gy) an detrimental effect on photoreceptors was reported [
38], recent publications indicate that photoreceptors, as non-replicating cells, are relatively stable to damage from radiation (e.g., brachytherapy, external beam radiotherapy, proton beam radiation, helium ion radiotherapy, and gamma knife radiotherapy at approximately 45 Gy) [
39]. However, impairment of the retinal pigment epithelium through radiation has already been described. It is characterized by a loss of melanin, an accumulation of lipofuscin and changes in choroidal vessels [
14,
39]. There are only a few publications on ERG and radiation retinopathy. Fujii et al. described in a book chapter that there was no significant change in the ERG results after beta radiation with 51 Gy of rabbit eyes [
40].
As part of this publication, the retinal function after proton irradiation, including the photoreceptor activity and function of the retinal pigment epithelium, was examined using the Ganzfeld ERG. In wild-type mice, no significant difference between the irradiated and control eyes could be observed. However, there was a tendency towards impairment of the retinal function after proton irradiation, especially of the photoreceptor activity and bipolar cells.
The role of inflammatory processes in the pathogenesis of radiation retinopathy is unclear. Clinical data and the success of treatment with corticosteroids suggest that an inflammatory reaction may be involved. In the few existing publications, microglia seem to play a major role [
1,
41,
42].
The results from the FACS experiments demonstrate no significant difference between irradiated and control eyes regarding the proportion of microglia (CD11b+CD45lo) and monocytes (CD11b+CD45hi) in the entire retina. These surface markers therefore do not seem to be increasingly expressed following proton radiation.
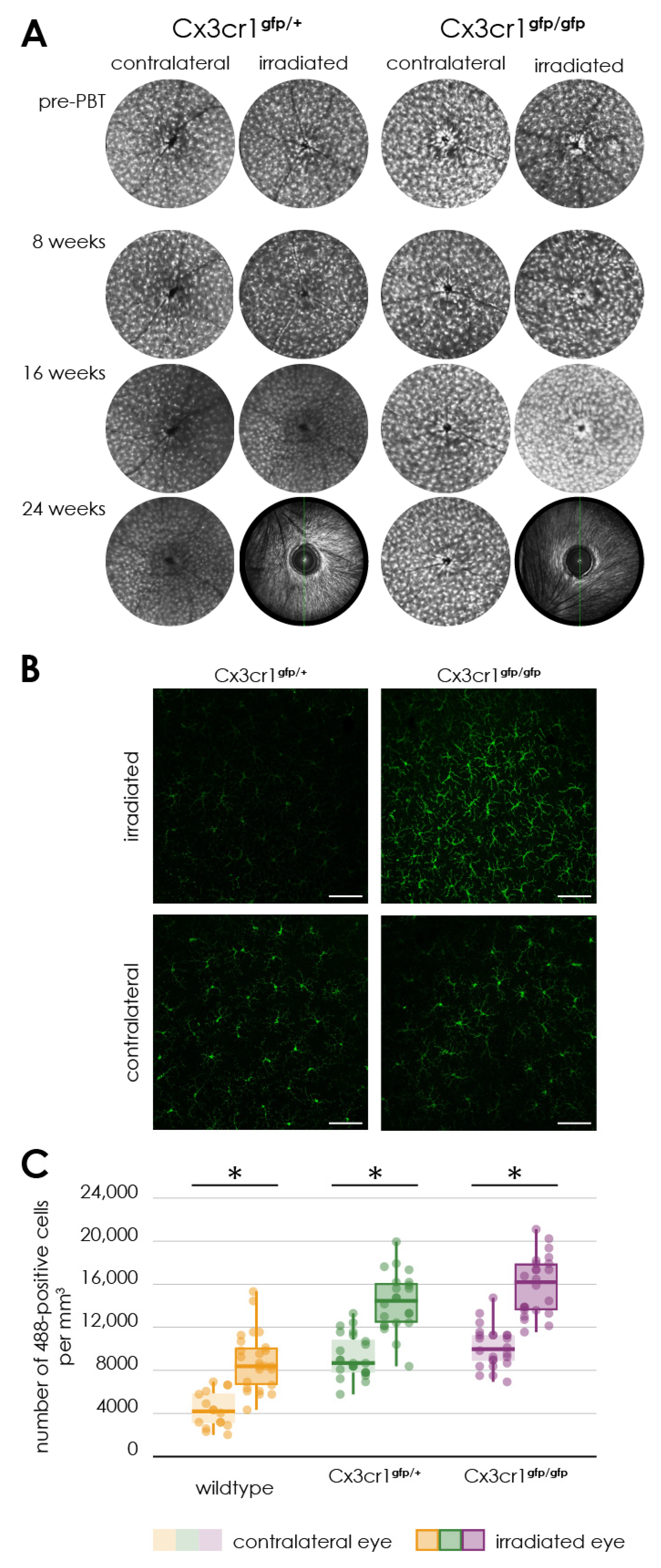
In contrast, the immunofluorescence staining of Iba1 in retinal flat mounts of the wild- type showed a significantly accumulation of Iba1 positive cells in the retina on the side of the irradiation compared to the control eye. Subsequently, the experiments were repeated with heterozygous Cx3cr1gfp/+ and homozygous Cx3cr1gfp/gfp mice to investigate the extent of inflammatory involvement respectively the influence of the Cx3cr1 signaling pathway on the processes in the retina after proton irradiation.
The Cx3cr1/CX3CL1 signaling pathway is mainly used to recruit monocytes to regions with injury and inflammation [
43]. This also applies to the retina. In the case of a disturbed blood-retina barrier, for example, recruited monocytes can be detected in the retina [
44]. Otherwise, in the retina the Cx3cr1 receptor is constant, invariably and apparently exclusively expressed by retinal microglial cells [
18]. It has already been published that Cx3cr1 has an important role in trafficking microglia cells to and from the subretinal space and that the ligand CX3CL1 displays pro-angiogenic activity [
18].
In general, the increased expression of Iba1 in microglia and macrophages correlates with increased inflammatory activity [
28]. Most recently, the increase in Iba-1 was associated with a detrimental microglia phenotype, called “dark microglia” [
45]. In the present publication, it could be demonstrated for the first time that targeted proton irradiation of eyes leads to a significant inflammatory reaction within the retina, maybe pushing the microglia towards the “dark” side.
In the literature, so far, no influence of the Cx3cr1 signaling pathway on the number of retinal pericytes has been described.
Comparable to wild-type mice, there is a significant decrease in the number of pericytes in the retinal flat mounts of the irradiated eyes of both Cx3cr1 genotypes, evenly in all areas and layers. Based on our experiments, the CX3cr1 signaling does not seem to have any influence on the decrease in the number of pericytes because of the irradiation.
According to the findings with the wild-type mice, the proton irradiation of both Cx3cr1 genotype mice also resulted in a significant increase in the expression of GFAP in the retina. However, there is no significant difference between the genotypes and no significant difference between the central and peripheral areas of the irradiated retina.
In our experiments, neither the proton irradiation in general nor the genotype respectively the absence of the Cx3cr1 signal, did affect the expression of Vimentin as a structural marker of Müller cells in the retina.
Regarding retinal function, ERG results from the irradiated eyes of heterozygous Cx3cr1gfp/+ and homozygous Cx3cr1gfp/gfp mice tend to be consistently reduced than the results of the control eyes 24 weeks after proton irradiation, which correspond to the results of the wild-type mice. Photoreceptor and bipolar cell activity are mainly affected.
In homozygous Cx3cr1gfp/gfp mice, a-waves and b-wave amplitudes at 3 cds/m2 were significantly worse on the irradiated side in relation to the contralateral side. However, the retinal function of homozygous Cx3cr1gfp/gfp mice seems to be generally better than of the heterozygous Cx3cr1gfp/+ and wildtype mice 24 weeks after irradiation. This difference is even significant regarding the function of the pigment epithelium and of the amacrine cells as well as their interconnections. The absence of the Cx3cr1 signaling obviously has a positive effect on the loss of retinal function after proton irradiation.
Like the findings in wild type, the density of Iba1/Cx3cr1 positive cells is increased after irradiation in both Cx3cr1 genotypes. In contrast to the results of the wild-type genotype, the distribution of Iba1/Cx3cr1 positive cells is different. In the wild-type genotype we observe an even increase in the superficial and the deep vascular layer, whereas in both Cx3cr1 genotypes the increase is more in the superficial than in the deep vascular layer. Since Cx3cr1 is related to trafficking of these cells, the cells might get “stuck” in the superficial layer and do not migrate to the deep vascular layer. Since the loss of Cx3cr1 seems to be protective on a functional level as measured with the ERG, the penetration of Iba1/Cx3cr1 positive cells into the deep vascular layer might be a hallmark of radiation-induced retinopathy.
In experimental diabetic retinopathy, there is also an increase in the number of activated microglial cells and migration of these cells into the outer plexiform layer and the photoreceptor layer [
46]. In this case Cx3cr1 deficiency activated microglia, enhanced the inflammatory response, disrupted the vascular integrity and accelerated the progression of diabetic retinopathy [
36].
In line with our findings, one other recent study using 45 Gy photon irradiation has shown increased inflammation and subsequent cell loss due to enhanced microglia activation and migration throughout the whole retina [
17].
In the context of this publication, the experimentally proton irradiation of mouse eyes was carried out for the first time and the consequences of this ocular irradiation were investigated. The morphological findings after irradiation are essentially characterized by the development of a dense cataract. A degenerative component is reflected by an increased expression of GFAP in the retina and the increased accumulation of Iba1 positive cells demonstrates an involvement of inflammatory processes. In addition, an impairment of retinal function can be observed. The Cx3cr1 signaling pathway appears to have at least a positive effect on the retinal function that might be due to a prevention of the infiltration of Cx3cr1/Iba positive cells into the deep vascular layer. Overall, the model is valid to study radiation-related changes in vivo. However, the impact of the Cx3cr1 and its mechanistic role regarding microglia migration and function is still speculative and should be investigated further. Functionally, while photoreceptor and bipolar cell activity were primarily affected, the general preservation of retinal function in Cx3cr1-deficient mice offers a promising avenue for therapeutic intervention. The protective role of Cx3cr1 deficiency highlights the need to explore strategies that selectively modulate microglial activity and migration without compromising their physiological roles in retinal homeostasis.
Given the limited therapeutic options currently available, these findings provide a foundation for developing targeted strategies aimed at preserving vascular integrity, modulating glial and inflammatory responses, and ultimately improving outcomes for patients undergoing proton irradiation. Future research should focus on the mechanistic role of the Cx3cr1 pathway and explore pharmacological interventions that can be translated into clinical settings such as intravitreal Injections of antibodies against Cx3cr1.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}