
Spent Hop (Humulus lupulus L.) Extract and Its Flaxseed Polysaccharide-Based Encapsulates Attenuate Inflammatory Bowel Diseases Through the Nuclear Factor-Kappa B, Extracellular Signal-Regulated Kinase, and Protein Kinase B Signalling Pathways
 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. SHE
2.2. Obtaining SHE and Its Encapsulation
2.3. Simulated Gastrointestinal Digestion of SHE and Its Encapsulates (ESHE)
2.4. Antioxidant Activity of SHE and Encapsulates
2.4.1. Preparation of Extracts from the Digested and Non-Digested SHE, FP and Encapsulates (ESHE) to Determine Antioxidant Activity
2.4.2. Determination of Total Polyphenol Content (TPC) by Folin–Ciocalteau Reagent (FCR)
2.4.3. Determination of Flavonoid Content
2.4.4. Determination of Antioxidant Activity Using ABTS+ Assay
2.4.5. Determination of Antioxidant Activity Using the DPPH Assay
2.4.6. Determination of Reducing Power Potential by the Method of FRAP Assay
2.4.7. Qualitative and Quantitative Analysis of Phenolic Compounds in Extract by UHPLC-DAD
2.5. Surface Morphology Assessment with Scanning Electron Microscopy
2.6. Cell Culture
2.7. Cell Viability Assay
2.8. RNA Extraction and Quantitative Real Time PCR (q-PCR)
2.9. Protein Samples Preparation and Western Blot Analysis
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Zymography Assay
2.12. Statistical Analysis
3. Results
3.1. The Composition of the Tested Agents
3.2. Assessment of the Surface Morphology of Microcapsules
3.3. Antioxidant Activity
3.4. Effect of the Tested Agents on Viability of HIEC-6 and CCD841CoN Cells
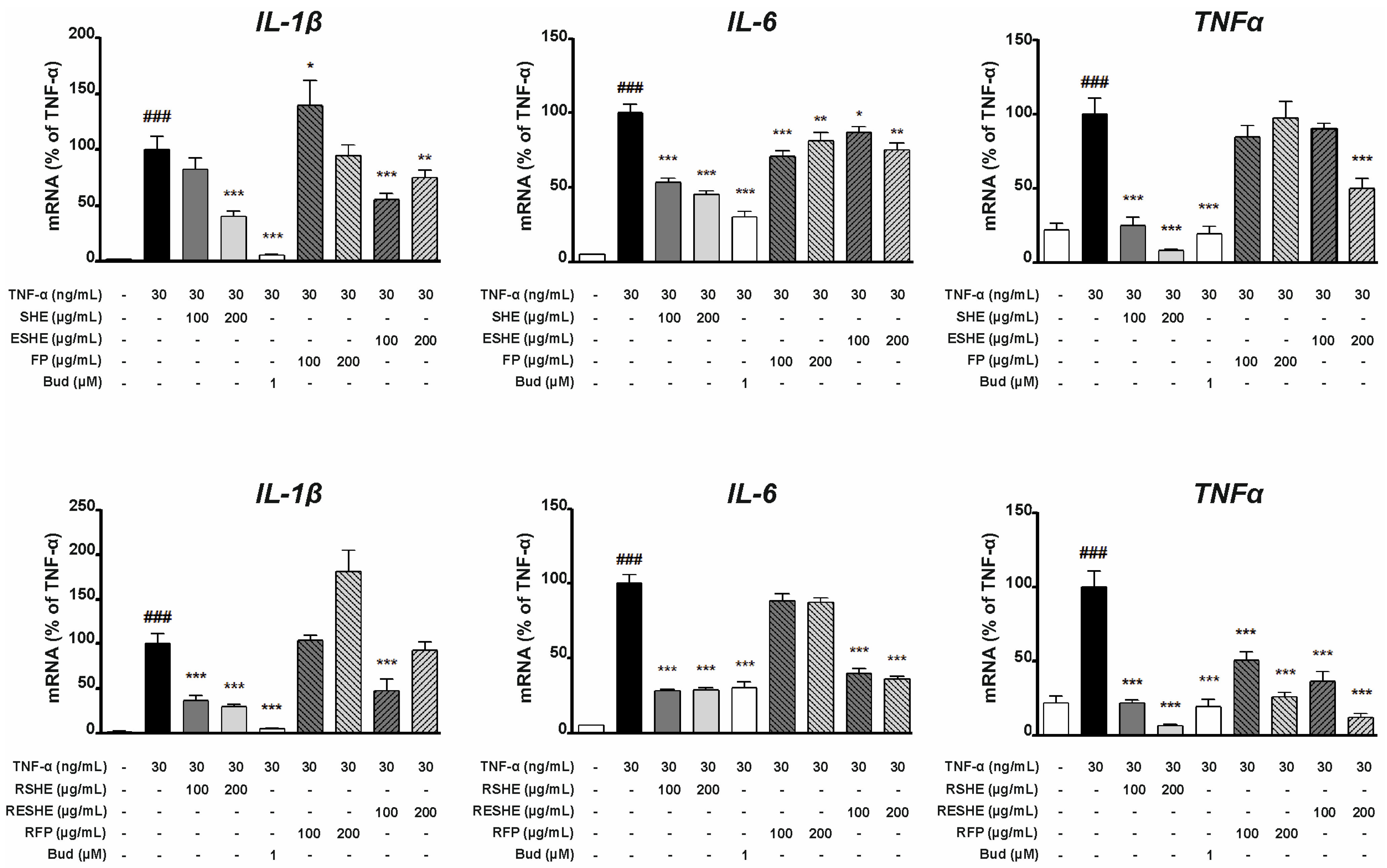
3.5. Effect of the Tested Agents on Expression of Pro-Inflammatory Cytokines (IL-1β, IL-6, TNF-α)
3.6. Effect of the Tested Agents on Expression and Activity of Gelatinases (MMP-2, MMP-9)
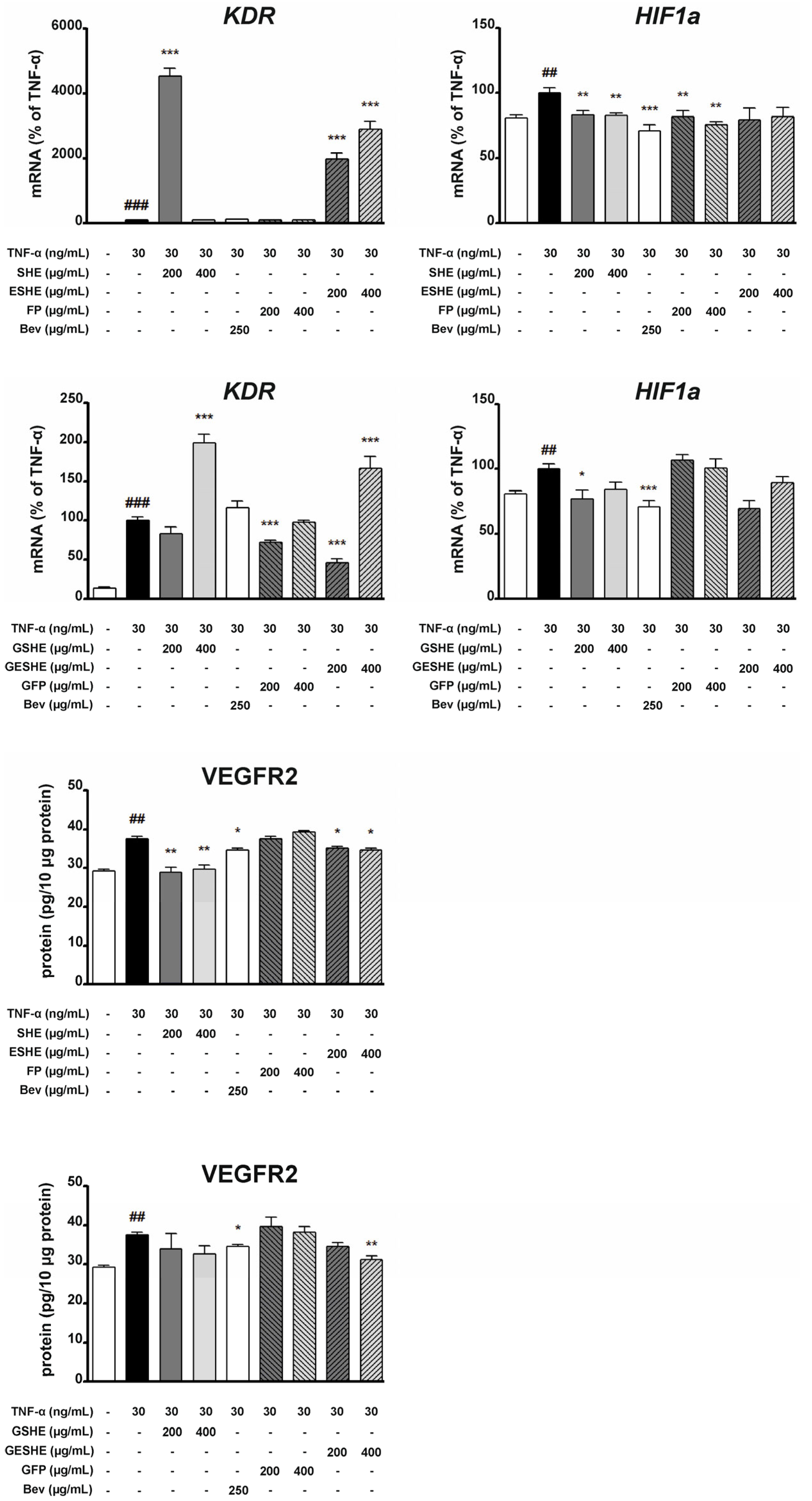
3.7. Effect of the Tested Agents on VEGFR2 and HIF1α Expression
3.8. Effect of the Tested Agents on the Activity of the Nuclear Factor Kappa B (NF-κB) Signalling Pathway
3.9. Effect of the Tested Agents on the Activity of the Extracellular Signal-Regulated Kinase (ERK) Signalling Pathway
3.10. Effect of the Tested Agents on the Activity of the Protein Kinase B (Akt) Signalling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bruner, L.P.; White, A.M.; Proksell, S. Inflammatory Bowel Diseases. Prim. Care 2023, 50, 411–427. [Google Scholar] [CrossRef]
- Kaplan, G.G.; Windsor, J.W. The four epidemiological stages in the global evolution of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q. A Comprehensive Review and Update on the Pathogenesis of Inflammatory Bowel Disease. J. Immunol. Res. 2019, 2019, 7247238. [Google Scholar] [CrossRef] [PubMed]
- Verstockt, B.; Verstockt, S.; Cremer, J.; Sabino, J.; Ferrante, M.; Vermeire, S.; Sudhakar, P. Distinct transcriptional signatures in purified circulating immune cells drive heterogeneity in disease location in IBD. BMJ Open Gastroenterol. 2023, 10, e001003. [Google Scholar] [CrossRef]
- Shah, S.C.; Itzkowitz, S.H. Colorectal Cancer in Inflammatory Bowel Disease: Mechanisms and Management. Gastroenterology 2022, 162, 715–730.e3. [Google Scholar] [CrossRef]
- Sinha, A.; Roy, S. Prospective therapeutic targets and recent advancements in the treatment of inflammatory bowel disease. Immunopharmacol. Immunotoxicol. 2024, 46, 550–563. [Google Scholar] [CrossRef]
- Cusimano, F.A.; Damas, O.M. Diet as a treatment for inflammatory bowel disease: Is it ready for prime time? Curr. Opin. Gastroenterol. 2022, 38, 358–372. [Google Scholar] [CrossRef]
- Reznikov, E.A.; Suskind, D.L. Current Nutritional Therapies in Inflammatory Bowel Disease: Improving Clinical Remission Rates and Sustainability of Long-Term Dietary Therapies. Nutrients 2023, 15, 668. [Google Scholar] [CrossRef]
- Roncoroni, L.; Gori, R.; Elli, L.; Tontini, G.E.; Doneda, L.; Norsa, L.; Cuomo, M.; Lombardo, V.; Scricciolo, A.; Caprioli, F.; et al. Nutrition in Patients with Inflammatory Bowel Diseases: A Narrative Review. Nutrients 2022, 14, 751. [Google Scholar] [CrossRef]
- Hossen, I.; Hua, W.; Ting, L.; Mehmood, A.; Song, J.; Duoxia, X.; Yanping, C.; Hongqing, W.; Zhipeng, G.; Kaiqi, Z.; et al. Phytochemicals and inflammatory bowel disease: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1321–1345. [Google Scholar] [CrossRef]
- Vaghari-Tabari, M.; Alemi, F.; Zokaei, M.; Moein, S.; Qujeq, D.; Yousefi, B.; Farzami, P.; Hosseininasab, S.S. Polyphenols and inflammatory bowel disease: Natural products with therapeutic effects? Crit. Rev. Food Sci. Nutr. 2024, 64, 4155–4178. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Mohana, T.; Athesh, K.; Hillary, V.E.; Vasconcelos, A.B.S.; de Franca, M.N.F.; Montalvão, M.M.; Ceasar, S.A.; Jothi, G.; Sridharan, G.; et al. Anti-inflammatory natural products modulate interleukins and their related signaling markers in inflammatory bowel disease: A systematic review. J. Pharm. Anal. 2023, 13, 1408–1428. [Google Scholar] [CrossRef]
- Caban, M.; Lewandowska, U. Encapsulation of polyphenolic compounds based on hemicelluloses to enhance treatment of inflammatory bowel diseases and colorectal cancer. Molecules 2023, 28, 4189. [Google Scholar] [CrossRef]
- Caban, M.; Chojnacka, K.; Owczarek, K.; Laskowska, J.; Fichna, J.; Podsedek, A.; Sosnowska, D.; Lewandowska, U. Spent hops (Humulus lupulus L.) extract as modulator of the inflammatory response in lipopolysaccharide stimulated RAW 264.7 macrophages. J. Physiol. Pharmacol. 2020, 71, 67–78. [Google Scholar]
- Caban, M.; Owczarek, K.; Chojnacka, K.; Podsedek, A.; Sosnowska, D.; Lewandowska, U. Chemopreventive properties of spent hops (Humulus lupulus L.) extract against angiogenesis, invasion and migration of colorectal cancer cells. J. Physiol. Pharmacol. 2022, 73, 431–442. [Google Scholar]
- Kowalska, G.; Rosicka-Kaczmarek, J.; Miśkiewicz, K.; Zakłos-Szyda, M.; Rohn, S.; Kanzler, C.; Wiktorska, M.; Niewiarowska, J. Arabinoxylan-Based Microcapsules Being Loaded with Bee Products as Bioactive Food Components are Able to Modulate the Cell Migration and Inflammatory Response—In Vitro Study. Nutrients 2022, 14, 2529. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, G.; Rosicka-Kaczmarek, J.; Miśkiewicz, K.; Wiktorska, M.; Gumul, D.; Orczykowska, M.; Dędek, K. Influence of Rye Bran Heteropolysaccharides on the Physicochemical and Antioxidant Properties of Honeydew Honey Microcapsules. Food Bioprod. Process. 2021, 130, 171–181. [Google Scholar] [CrossRef]
- El Hariri, B.; Sallé, G.; Andary, C. Involvement of flavonoids in the resistance of two poplar cultivars to mistletoe (Viscum album L.). Protoplasma 1991, 162, 20–26. [Google Scholar] [CrossRef]
- Gumul, D.; Oracz, J.; Kowalski, S.; Mikulec, A.; Skotnicka, M.; Karwowska, K.; Areczuk, A. Bioactive Compounds and Antioxidant Composition of Nut Bars with Addition of Various Edible Insect Flours. Molecules 2023, 28, 3556. [Google Scholar] [CrossRef]
- Kim, J.-S. Antioxidant Activity of Various Soluble Melanoidins Isolated from Black Garlic after Different Thermal Processing Steps. Prev. Nutr. Food Sci. 2020, 25, 301–309. [Google Scholar] [CrossRef]
- Oracz, J.; Żyżelewicz, D.; Pacholczyk-Sienicka, B. UHPLC-DAD-ESI-HRMS/MS profile of phenolic compounds in northern red oak (Quercus rubra L., syn. Q. borealis F. Michx) seeds and its transformation during thermal processing. Ind. Crops Prod. 2022, 189, 115860. [Google Scholar]
- Zhang, J.; Cen, L.; Zhang, X.; Tang, C.; Chen, Y.; Zhang, Y.; Yu, M.; Lu, C.; Li, M.; Li, S.; et al. MPST deficiency promotes intestinal epithelial cell apoptosis and aggravates inflammatory bowel disease via AKT. Redox Biol. 2022, 56, 102469. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.F.; Caetano, M.A.F.; Magalhães, H.I.R.; Castelucci, P. Study of tumor necrosis factor receptor in the inflammatory bowel disease. World J. Gastroenterol. 2023, 29, 2733–2746. [Google Scholar] [CrossRef] [PubMed]
- Olivera, G.C.; Ren, X.; Vodnala, S.K.; Lu, J.; Coppo, L.; Leepiyasakulchai, C.; Holmgren, A.; Kristensson, K.; Rottenberg, M.E. Nitric Oxide Protects against Infection-Induced Neuroinflammation by Preserving the Stability of the Blood-Brain Barrier. PLoS Pathog. 2016, 12, e1005442. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, H.A.; Quinn, G.A.; Nasef, M.M.; Mishra, V.; Aljabali, A.A.A.; El-Tanani, M.; Serrano-Aroca, Á.; Da Silva, M.W.; McCarron, P.A.; Tambuwala, M.M. Crocin Inhibits Angiogenesis and Metastasis in Colon Cancer via TNF-α/NF-kB/VEGF Pathways. Cells 2022, 11, 1502. [Google Scholar] [CrossRef]
- Hu, M.; Huang, Y.; Du, X.; Liu, G.; Qi, B.; Li, Y. The synergistic effect of epigallocatechin-3-gallate and quercetin co-loaded hydrogel beads on inflammatory bowel disease. Food Funct. 2023, 14, 4539–4551. [Google Scholar] [CrossRef]
- Lu, S.-Y.; Dan, L.; Sun, S.; Fu, T.; Chen, J. Dietary quercetin intake is associated with lower ulcerative colitis risk but not Crohn’s disease in a prospective cohort study and in vivo experiments. Food Funct. 2024, 15, 6553–6564. [Google Scholar] [CrossRef]
- Ashiqueali, S.A.; Chaudhari, D.; Zhu, X.; Noureddine, S.; Siddiqi, S.; Garcia, D.N.; Gostynska, A.; Stawny, M.; Rubis, B.; Zanini, B.M.; et al. Fisetin modulates the gut microbiota alongside biomarkers of senescence and inflammation in a DSS-induced murine model of colitis. Geroscience 2024, 46, 3085–3103. [Google Scholar] [CrossRef]
- Ekhtiar, M.; Ghasemi-Dehnoo, M.; Mirzaei, Y.; Azadegan-Dehkordi, F.; Amini-Khoei, H.; Lorigooini, Z.; Samiei-Sefat, A.; Bagheri, N. The coumaric acid and syringic acid ameliorate acetic acid-induced ulcerative colitis in rats via modulator of Nrf2/HO-1 and pro-inflammatory cytokines. Int. Immunopharmacol. 2025, 120, 110309. [Google Scholar] [CrossRef]
- Antonio, A.M.R.; Martínez, I.I.P.; Márquez-Flores, Y.K.; Solano, A.H.J.; Ramos, M.A.T.; Hernández, M.C.R. Protective effect of (E)-(2,4-dihydroxy)-α-aminocinnamic acid, a hydroxy cinnamic acid derivative, in an ulcerative colitis model induced by TNBS. Biosci. Rep. 2024, 44, BSR20240797. [Google Scholar]
- Brás, J.P.; Bravo, J.; Freitas, J.; Barbosa, M.A.; Santos, S.G.; Summavielle, T.; Almeida, M.I. TNF-alpha-induced microglia activation requires miR-342: Impact on NF-kB signaling and neurotoxicity. Cell Death Dis. 2020, 11, 415. [Google Scholar] [CrossRef]
- Godala, M.; Gaszyńska, E.; Walczak, K.; Małecka-Wojciesko, E. Role of Serum Interleukin-6, Interleukin-1β and Interleukin-10 in Assessment of Disease Activity and Nutritional Status in Patients with Inflammatory Bowel Disease. J. Clin. Med. 2023, 12, 5956. [Google Scholar] [CrossRef]
- Dudek, P.; Talar-Wojnarowska, R. Current Approach to Risk Factors and Biomarkers of Intestinal Fibrosis in Inflammatory Bowel Disease. Medicina 2024, 60, 305. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Tan, H.; Wang, J.; Huang, L.; Liu, H.; Shi, Y.; Zhong, C.; Weng, S.; Chen, C.; Zhao, W.; et al. β-hydroxybutyrate restrains colitis-associated tumorigenesis by inhibiting HIF-1α-mediated angiogenesis. Cancer Lett. 2024, 593, 216940. [Google Scholar] [CrossRef] [PubMed]
- Laurindo, L.F.; de Oliveira dos Santos, A.R.; de Carvalho, A.C.A.; Bechara, M.D.; Guiguer, E.L.; de Alvares Goulart, R.; Sinatora, R.V.; Araújo, A.C.; Barbalho, S.M. Phytochemicals and Regulation of NF-kB in Inflammatory Bowel Diseases: An Overview of In Vitro and In Vivo Effects. Metabolites 2023, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Lu, B.; Xi, J.; Ocansey, D.K.W.; Mao, F.; Hao, D.; Yan, Y. hucMSC-Ex Alleviates IBD-Associated Intestinal Fibrosis by Inhibiting ERK Phosphorylation in Intestinal Fibroblasts. Stem Cells Int. 2023, 2023, 2828981. [Google Scholar] [CrossRef]
- Wang, Z.; Li, B.; Bao, L.; Chen, Y.; Yang, J.; Xu, F.; Shi, S.; Chen, W.; Wang, B.; Liu, Y. Fusobacterium Nucleatum Aggravates Intestinal Barrier Impairment and Colitis Through IL-8 Induced Neutrophil Chemotaxis by Activating Epithelial Cells. J. Inflamm. Res. 2024, 17, 8407–8420. [Google Scholar] [CrossRef]
- Liu, B.; Piao, X.; Niu, W.; Zhang, Q.; Ma, C.; Wu, T.; Gu, Q.C.; Cui, T.; Li, S. Kuijieyuan Decoction Improved Intestinal Barrier Injury of Ulcerative Colitis by Affecting TLR4-Dependent PI3K/AKT/NF-κB Oxidative and Inflammatory Signaling and Gut Microbiota. Front. Pharmacol. 2020, 11, 1036. [Google Scholar] [CrossRef]
- Dong, L.; Du, H.; Zhang, M.; Xu, H.; Pu, X.; Chen, Q.; Luo, R.; Hu, Y.; Wang, Y.; Tu, H.; et al. Anti-inflammatory effect of Rhein on ulcerative colitis via inhibiting PI3K/Akt/mTOR signaling pathway and regulating gut microbiota. Phytother. Res. 2022, 36, 2081–2094. [Google Scholar] [CrossRef]
- Ntuli, S.; Leuschner, M.; Bester, M.J.; Serem, J.C. Stability, Morphology, and Effects of In Vitro Digestion on the Antioxidant Properties of Polyphenol Inclusion Complexes with β-Cyclodextrin. Molecules 2022, 27, 3808. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Liu, J.; Lei, L.; Ye, F.; Zhou, Y.; Younis, H.G.R.; Zhao, G. Aggregates of octenylsuccinate oat β-glucan as novel capsules to stabilize curcumin over food processing, storage and digestive fluids and to enhance its bioavailability. Food Funct. 2018, 9, 491–501. [Google Scholar] [CrossRef]
- Li, T.; Teng, D.; Mao, R.; Hao, Y.; Wang, X.; Wang, J. Recent progress in preparation and agricultural application of microcapsules. J. Biomed. Mater. Res. Part A 2019, 107, 2371–2385. [Google Scholar] [CrossRef]
- Tran, N.; Tran, M.; Truong, H.; Le, L. Spray-Drying Microencapsulation of High Concentration of Bioactive Compounds Fragments from Euphorbia hirta L. Extract and Their Effect on Diabetes Mellitus. Foods 2020, 9, 881. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Li, T.; He, F.; Chen, C.; Xu, X.; Tian, W.; Yang, Y.; He, X.; Li, H.; Chen, K.; et al. Microencapsulation of nattokinase from fermentation by spray drying: Optimization, comprehensive score, and stability. Food Sci. Nutr. 2021, 9, 3906–3916. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.M.; Arpagaus, C.; Cerqueira, M.A.; Samborska, K. Nano spray drying of food ingredients; materials, processing and applications. Trends Food Sci. Technol. 2021, 109, 632–646. [Google Scholar] [CrossRef]
- Saleem, M.H.; Ali, S.; Hussain, S.; Kamran, M.; Chattha, M.S.; Ahmad, S.; Aqeel, M.; Rizwan, M.; Aljarba, N.H.; Alkahtani, S.; et al. Flax (Linum usitatissimum L.): A Potential Candidate for Phytoremediation? Biological and Economical Points of View. Plants 2020, 9, 496. [Google Scholar] [CrossRef]
- Haseeb, M.T.; Muhammad, G.; Hussain, M.A.; Bukhari, S.N.A.; Sheikh, F.A. Flaxseed (Linum usitatissimum) mucilage: A versatile stimuli-responsive functional biomaterial for pharmaceuticals and healthcare. Int. J. Biol. Macromol. 2024, 278, 134817. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Sharma, A.; Singhal, R.S. A water-absorbent hydrogel prepared with arabinoxylans from flaxseed (Linum usitatissimum) and galactomannans from subabul (Leucaena leucocephala) impacts the growth and composition of cherry tomatoes in potted experiments. J. Clean. Prod. 2023, 429, 139346. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Z.; Huang, F.; Yang, C.; Huang, Q. In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed. Molecules 2020, 25, 4354. [Google Scholar] [CrossRef]
- Sommella, E.; Verna, G.; Liso, M.; Salviati, E.; Esposito, T.; Carbone, D.; Pecoraro, C.; Chieppa, M.; Campiglia, P. Hop-derived fraction rich in beta acids and prenylflavonoids regulates the inflammatory response in dendritic cells differently from quercetin: Unveiling metabolic changes by mass spectrometry-based metabolomics. Food Funct. 2021, 12, 12800–12811. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Tao, S.; Guo, F.; Wang, B.; Yang, L.; Ma, L.; Fu, P. Natural flavonol fisetin attenuated hyperuricemic nephropathy via inhibiting IL-6/JAK2/STAT3 and TGF-β/SMAD3 signaling. Phytomedicine 2021, 87, 153552. [Google Scholar] [CrossRef]
- Ren, Q.; Guo, F.; Tao, S.; Huang, R.; Ma, L.; Fu, P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-κB p65 and MAPK signaling pathways in septic AKI mice. Biomed. Pharmacother. 2020, 122, 109772. [Google Scholar] [CrossRef]
- Zaragozá, C.; Villaescusa, L.; Monserrat, J.; Zaragozá, F.; Álvarez-Mon, M. Potential Therapeutic Anti-Inflammatory and Immunomodulatory Effects of Dihydroflavones, Flavones, and Flavonols. Molecules 2020, 25, 1017. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agent | Phenolic Compounds Amount [mg/100 g] | Hydroxycinnamic Acids Amount [mg/100 g] | Hydroxybenzoic Acids Amount [mg/100 g] | Flavan-3-Ols Amount [mg/100 g] | Flavonols Amount [mg/100 g] | Prenylflavonoids Amount [mg/100 g] |
|---|---|---|---|---|---|---|
| SHE | 879.73 ± 3.21 | 207.16 ± 1.02 | 19.48 ± 0.20 | 9.53 ± 0.10 | 554.75 ± 4.21 | 88.81 ± 1.21 |
| GSHE | 305.11 ± 2.01 | 72.08 ± 0.68 | 13.49 ± 0.45 | 1.89 ± 0.04 | 209.03 ± 1.98 | 8.62 ± 0.25 |
| RSHE | 131.17 ± 0.95 | 27.30 ± 0.45 | 3.64 ± 0.10 | nd | 74.88 ± 0.85 | 25.35 ± 1.12 |
| ESHE | 543.03 ± 1.04 | 142.25 ± 1.02 | 12.77 ± 0.21 | 1.80 ± 0.08 | 347.32 ± 2.11 | 38.89 ± 0.89 |
| GESHE | 192.53 ± 0.98 | 48.25 ± 0.60 | 9.96 ± 0.11 | 4.87 ± 0.16 | 121.82 ± 1.01 | 7.63 ± 0.27 |
| RESHE | 230.76 ± 1.10 | 36.72 ± 1.11 | 6.07 ± 0.10 | 4.50 ± 0.10 | 151.04 ± 1.11 | 32.43 ± 0.79 |
| FP | 15.23 ± 0.38 | 8.77 ± 0.11 | 3.65 ± 0.43 | nd | 2.81 ± 0.30 | nd |
| GFP | 5.26 ± 0.23 | 4.46 ± 0.28 | 0.80 ± 0.10 | nd | nd | nd |
| RFP | 1.82 ± 0.08 | 1.82 ± 0.08 | nd | nd | nd | nd |
| Agent | Flavonols Amount [mg/100 g] | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Quercetin | Quercetin 3-O-Glucoside | Quercetin 3-O-Arabinoside | Quercetin 3-O-Galactoside | Quercetin 3-O-Rutinoside | Quercetin 3-O-Rhamnoside | Kaempferol 3-O-Galactoside | Kaempferol 3-O-Rutinoside | Kaempferol 3-O-Glucoside | Total | |
| SHE | 5.91 ± 0.09 | 166.81 ± 1.00 | 63.30 ± 0.67 | 4.80 ± 0.05 | 148.39 ± 0.98 | 9.08 ± 0.90 | 54.88 ± 0.45 | 64.33 ± 0.67 | 37.25 ± 0.25 | 554.75 ± 4.21 |
| GSHE | nd | 58.25 ± 0.48 | 27.49 ± 0.29 | 0.66 ± 0.01 | 58.64 ± 0.30 | 4.49 ± 0.50 | 21.20 ± 0.30 | 21.85 ± 0.30 | 16.45 ± 0.10 | 209.03 ± 1.98 |
| RSHE | nd | 23.16 ± 0.20 | 7.37 ± 0.08 | 0.58 ± 0.02 | 21.15 ± 0.45 | 1.26 ± 0.10 | 8.19 ± 0.10 | 9.32 ± 0.10 | 3.85 ± 0.03 | 74.88 ± 0.85 |
| ESHE | 3.20 ± 0.08 | 100.17 ± 1.02 | 42.20 ± 0.45 | 4.70 ± 0.05 | 93.51 ± 0.98 | 6.13 ± 0.10 | 34.11 ± 0.25 | 38.30 ± 0.40 | 25.00 ± 0.30 | 347.32 ± 2.11 |
| GESHE | nd | 33.21 ± 0.34 | 16.18 ± 0.20 | 1.55 ± 0.01 | 34.79 ± 0.42 | 2.74 ± 0.34 | 11.90 ± 0.20 | 11.81 ± 0.23 | 9.64 ± 0.09 | 121.82 ± 1.01 |
| RESHE | nd | 43.15 ± 0.45 | 18.31 ± 0.21 | 2.27 ± 0.02 | 42.59 ± 0.51 | 2.77 ± 0.21 | 15.60 ± 0.12 | 16.92 ± 0.15 | 9.43 ± 0.10 | 151.04 ± 1.11 |
| FP | nd | nd | nd | nd | nd | nd | 0.91 ± 0.04 | 1.90 ± 0.01 | nd | 2.81 ± 0.30 |
| GFP | nd | nd | nd | nd | nd | nd | nd | nd | nd | 0 |
| RFP | nd | nd | nd | nd | nd | nd | nd | nd | nd | 0 |
| Agent | Hydroxycinnamic Acids Amount [mg/100 g] | |||||
|---|---|---|---|---|---|---|
| Caffeic Acid | Ferulic Acid | p-Coumaric Acid | Chlorogenic Acid | Neochlorogenic Acid | Total | |
| SHE | 20.64 ± 0.21 | 5.05 ± 0.20 | 14.33 ± 0.70 | 38.31 ± 0.87 | 128.83 ± 1.30 | 207.16 ± 1.02 |
| GSHE | 8.34 ± 0.10 | 7.55 ± 0.26 | 4.58 ± 0.20 | 11.00 ± 0.70 | 40.61 ± 0.98 | 72.08 ± 0.68 |
| RSHE | 3.29 ± 0.15 | 2.25 ± 0.10 | 10.64 ± 0.60 | 2.39 ± 0.11 | 8.73 ± 0.32 | 27.30 ± 0.45 |
| ESHE | 9.08 ± 0.25 | 4.53 ± 0.21 | 10.99 ± 0.50 | 25.81 ± 0.22 | 91.84 ± 1.07 | 142.25 ± 1.02 |
| GESHE | 5.85 ± 0.40 | 1.93 ± 0.09 | 9.34 ± 0.36 | 6.80 ± 0.30 | 24.33 ± 0.78 | 48.25 ± 0.60 |
| RESHE | 6.66 ± 0.41 | 1.90 ± 0.08 | 5.77 ± 0.20 | 5.00 ± 0.45 | 17.39 ± 1.01 | 36.72 ± 1.11 |
| FP | nd | 4.85 ± 0.11 | 3.92 ± 0.10 | nd | nd | 8.77 ± 0.11 |
| GFP | nd | 2.43 ± 0.10 | 2.03 ± 0.30 | nd | nd | 4.46 ± 0.28 |
| RFP | nd | 0.98 ± 0.06 | 0.84 ± 0.05 | nd | nd | 1.82 ± 0.08 |
| Agent | Hydroxybenzoic Acids Amount [mg/100 g] | ||||
|---|---|---|---|---|---|
| Vanillic Acid | Syringic Acid | 3,4-Dihydroxybenzoic Acid | 4-Hydroxybenzoic Acid | Total | |
| SHE | nd | 4.41 ± 0.20 | 9.06 ± 0.09 | 6.01 ± 0.10 | 19.48 ± 0.20 |
| GSHE | 3.44 ± 0.18 | 1.65 ± 0.10 | 7.10 ± 0.35 | 1.30 ± 0.10 | 13.49 ± 0.45 |
| RSHE | 0.72 ± 0.01 | 0.53 ± 0.08 | 2.30 ± 0.15 | 0.09 ± 0.02 | 3.64 ± 0.10 |
| ESHE | nd | 2.65 ± 0.10 | 5.52 ± 0.19 | 4.60 ± 0.5 | 12.77 ± 0.21 |
| GESHE | 2.55 ± 0.11 | 1.00 ± 0.09 | 5.94 ± 0.10 | 0.47 ± 0.01 | 9.96 ± 0.11 |
| RESHE | 1.85 ± 0.09 | 1.07 ± 0.11 | 3.04 ± 0.14 | 0.11 ± 0.03 | 6.07 ± 0.10 |
| FP | nd | 0.52 ± 0.08 | nd | 3.13 ± 0.10 | 3.65 ± 0.43 |
| GFP | nd | nd | nd | 0.80 ± 0.06 | 0.80 ± 0.06 |
| RFP | nd | nd | nd | nd | nd |
| Agent | Prenylflavonoids Amount [mg/100 g] | |||
|---|---|---|---|---|
| Isoxanthohumol | Xanthohumol | 8-Prenylnaringenin | Total | |
| SHE | 6.25 ± 0.10 | 72.70 ± 1.11 | 9.86 ± 0.45 | 88.81 ± 1.21 |
| GSHE | 1.14 ± 0.10 | 7.04 ± 0.35 | 0.44 ± 0.04 | 8.62 ± 0.25 |
| RSHE | 3.09 ± 0.34 | 20.76 ± 1.01 | 1.50 ± 0.50 | 25.35 ± 1.12 |
| ESHE | 4.94 ± 0.36 | 32.01 ± 1.12 | 1.94 ± 0.10 | 38.89 ± 0.98 |
| GESHE | 2.18 ± 0.17 | 5.04 ± 0.30 | 0.41 ± 0.03 | 7.63 ± 0.27 |
| RESHE | 5.63 ± 0.46 | 24.93 ± 1.10 | 1.87 ± 0.40 | 32.43 ± 0.79 |
| FP | nd | nd | nd | nd |
| GFP | nd | nd | nd | nd |
| RFP | nd | nd | nd | nd |
| Agent | Total Polyphenol Content [mg GAE/100 g] | Total Flavonoid Content [mg QE/100 g] | ABTS [mM TE/100 g] | DPPH [mM TE/100 g] | FRAP [mM Fe2+/100 g] |
|---|---|---|---|---|---|
| SHE | 4418.48 ± 1.91 | 2295.71 ± 1.56 | 52.91 ± 1.34 | 34.07 ± 3.29 | 14.97 ± 0.98 |
| GSHE | 1304.44 ± 1.25 | 353.11 ± 0.98 | 13.04 ± 1.27 | 1.47 ± 0.22 | 9.26 ± 0.31 |
| RSHE | 775.01 ± 0.98 | 302.80 ± 1.03 | 11.92 ± 1.14 | 0.51 ± 0.09 | 5.19 ± 0.30 |
| ESHE | 2758.32 ± 1.98 | 1389.72 ± 1.65 | 29.51 ± 3.31 | 16.79 ± 3.18 | 7.64 ± 0.23 |
| GESHE | 1510.40 ± 1.45 | 601.33 ± 1.21 | 20.19 ± 3.01 | 5.91 ± 0.27 | 13.66 ± 0.40 |
| RESHE | 1741.70 ± 0.98 | 939.38 ± 1.45 | 36.16 ± 2.45 | 1.38 ± 0.21 | 12.67 ± 0.38 |
| FP | 242.49 ± 1.65 | 103.49 ± 1.45 | 5.53 ± 0.15 | 16.85 ± 3.08 | 2.90 ± 1.45 |
| GFP | 219.22 ± 0.78 | 62.84 ± 0.55 | 8.01 ± 0.20 | 3.82 ± 0.10 | 3.43 ± 0.20 |
| RFP | 337.61 ± 0.86 | 51.05 ± 0.61 | 13.65 ± 0.98 | 4.24 ± 0.09 | 4.06 ± 0.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caban, M.; Owczarek, K.; Rosicka-Kaczmarek, J.; Miśkiewicz, K.; Oracz, J.; Pawłowski, W.; Niewinna, K.; Lewandowska, U. Spent Hop (Humulus lupulus L.) Extract and Its Flaxseed Polysaccharide-Based Encapsulates Attenuate Inflammatory Bowel Diseases Through the Nuclear Factor-Kappa B, Extracellular Signal-Regulated Kinase, and Protein Kinase B Signalling Pathways. Cells 2025, 14, 1099. https://doi.org/10.3390/cells14141099
Caban M, Owczarek K, Rosicka-Kaczmarek J, Miśkiewicz K, Oracz J, Pawłowski W, Niewinna K, Lewandowska U. Spent Hop (Humulus lupulus L.) Extract and Its Flaxseed Polysaccharide-Based Encapsulates Attenuate Inflammatory Bowel Diseases Through the Nuclear Factor-Kappa B, Extracellular Signal-Regulated Kinase, and Protein Kinase B Signalling Pathways. Cells. 2025; 14(14):1099. https://doi.org/10.3390/cells14141099
Chicago/Turabian StyleCaban, Miłosz, Katarzyna Owczarek, Justyna Rosicka-Kaczmarek, Karolina Miśkiewicz, Joanna Oracz, Wojciech Pawłowski, Karolina Niewinna, and Urszula Lewandowska. 2025. "Spent Hop (Humulus lupulus L.) Extract and Its Flaxseed Polysaccharide-Based Encapsulates Attenuate Inflammatory Bowel Diseases Through the Nuclear Factor-Kappa B, Extracellular Signal-Regulated Kinase, and Protein Kinase B Signalling Pathways" Cells 14, no. 14: 1099. https://doi.org/10.3390/cells14141099
APA StyleCaban, M., Owczarek, K., Rosicka-Kaczmarek, J., Miśkiewicz, K., Oracz, J., Pawłowski, W., Niewinna, K., & Lewandowska, U. (2025). Spent Hop (Humulus lupulus L.) Extract and Its Flaxseed Polysaccharide-Based Encapsulates Attenuate Inflammatory Bowel Diseases Through the Nuclear Factor-Kappa B, Extracellular Signal-Regulated Kinase, and Protein Kinase B Signalling Pathways. Cells, 14(14), 1099. https://doi.org/10.3390/cells14141099