


Brain Organoid Transplantation: A Comprehensive Guide to the Latest Advances and Practical Applications—A Systematic Review



Abstract
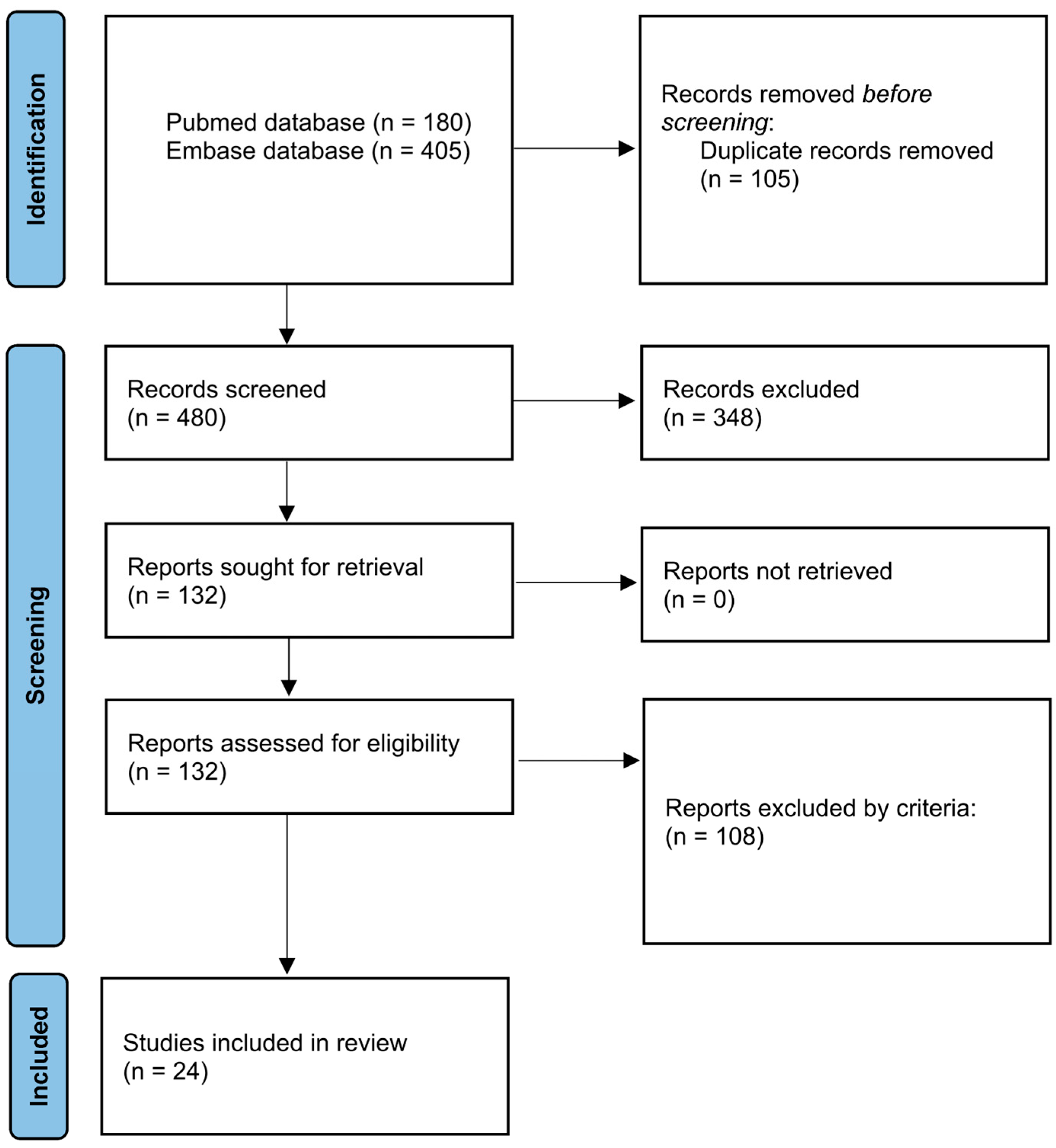
1. Background
2. Search Strategy
3. Selection Criteria
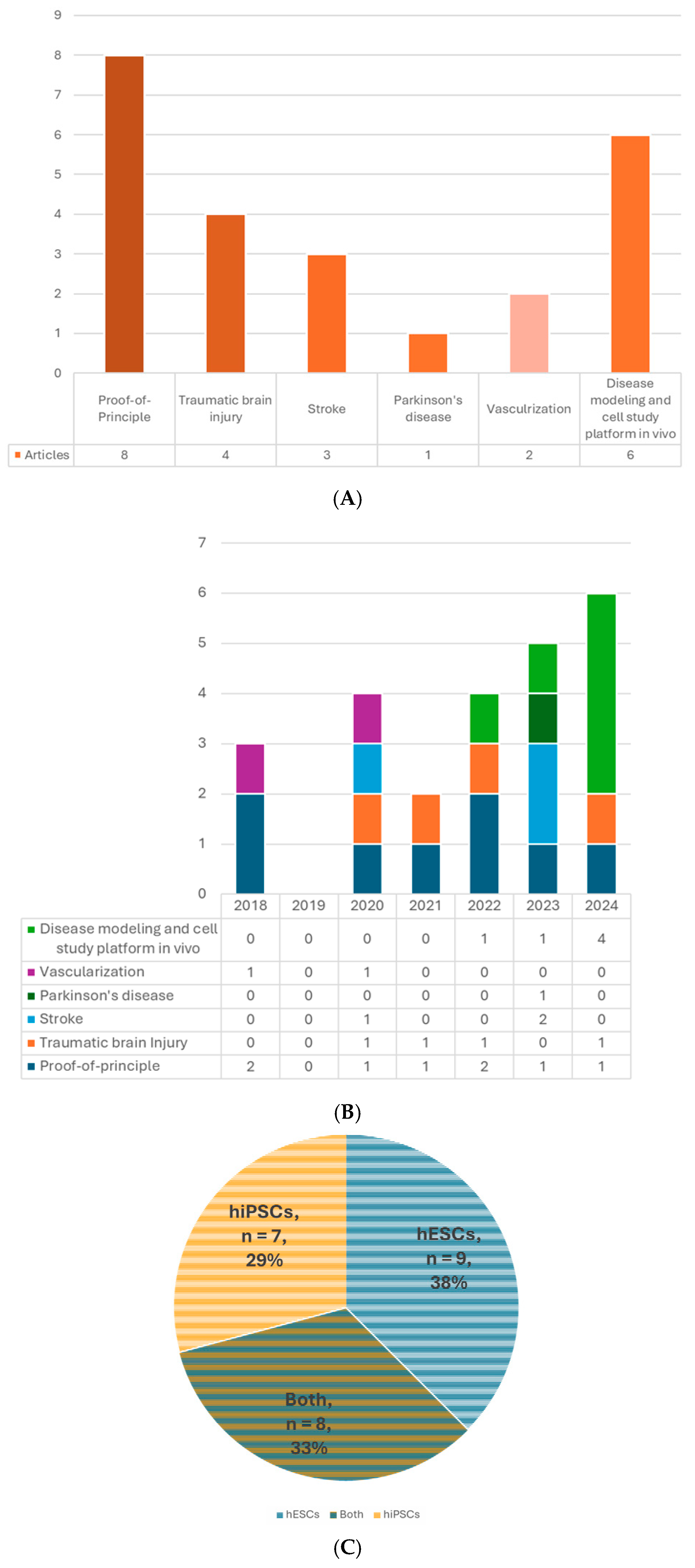
4. Overview
5. Organoid
5.1. Cell Source
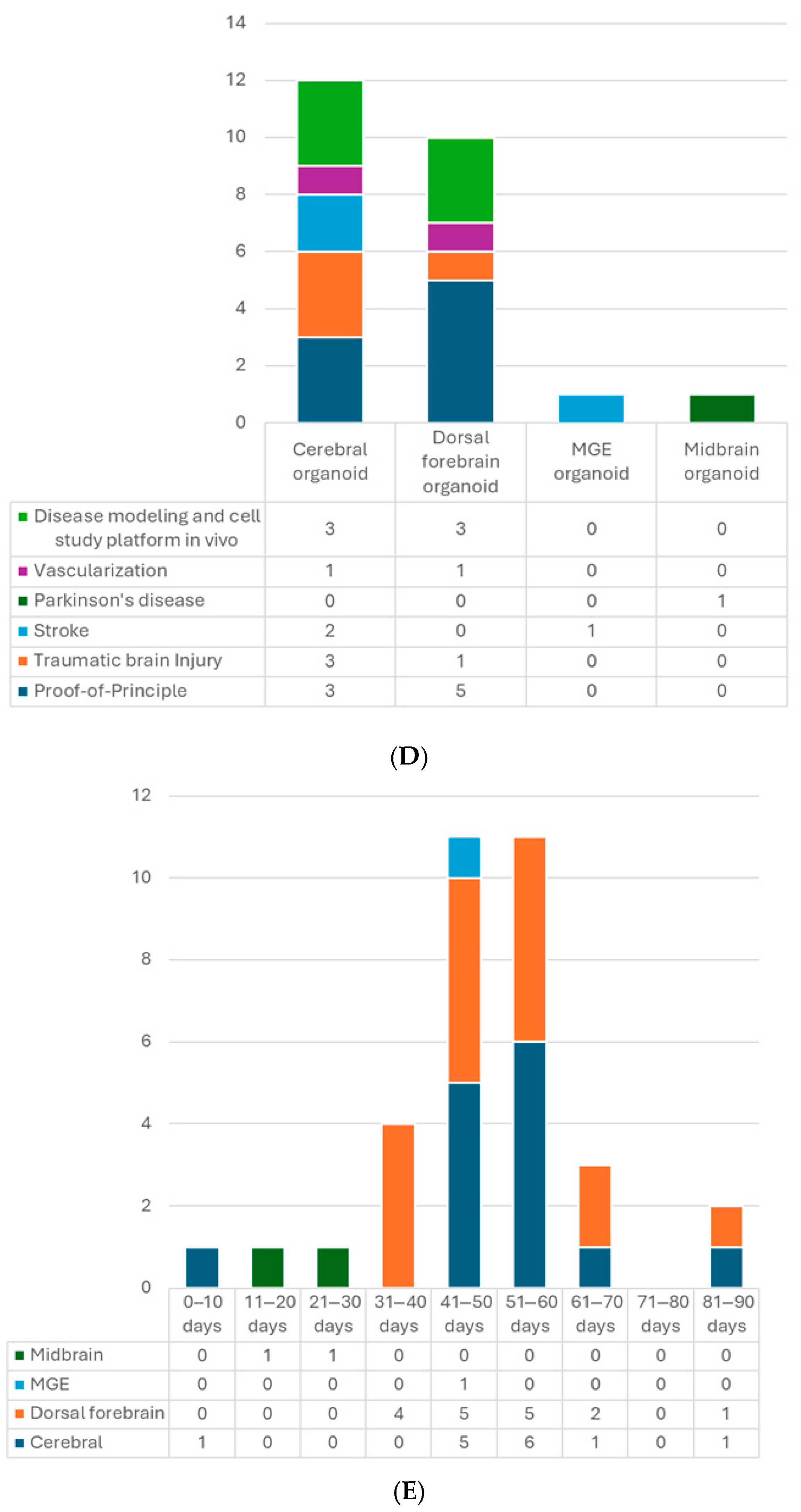
5.2. Types of Organoids
5.3. Age of Organoids at the Time of Transplantation
5.4. Size and Cell Number
5.5. Additional Treatment of the Organoids In Vitro
5.6. Brain Organoids for Transplantation
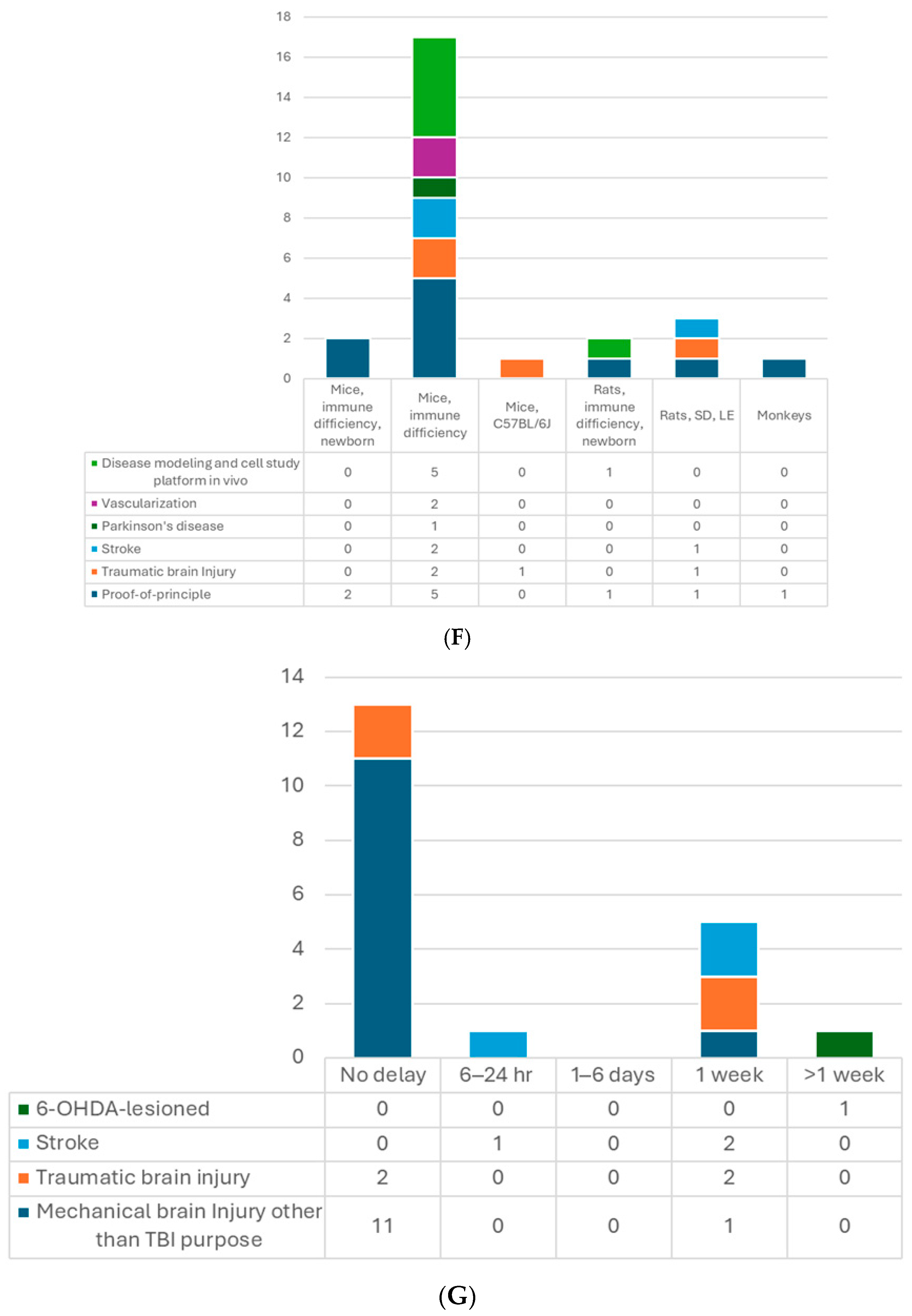
6. Recipient
7. Transplantation
7.1. Site of Transplantation
7.2. The Timing of Transplantation
7.3. Procedure
8. Outcomes
8.1. General Survival
8.2. Cell Differentiation and Maturation
8.3. Axonal Projection, Cell Migration, and Synaptic Formation
8.4. Electrophysiological Neuronal Activity and Functional Integration
8.5. Vascularization
8.6. Host Brain Aspects—Neurogenesis and Immune Modulation
8.7. Host Individual Aspects—Behavioral Function
9. Post-Transplantation Modulation In Vivo
10. Current Applications
11. Safety Concerns
12. Current Ethics
13. Challenges and Future Perspectives
13.1. Cell Diversity
13.2. Heterogeneity, Necrotic Core, and Quality Control
13.3. Time Issues
13.4. Translational Differences
13.5. Essential Brain Injury
13.6. Ethics for the Future
13.7. Brain Organoids on Chip
14. Limitation of the Review
15. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, X.-Y.; Wu, S.; Wang, D.; Chu, C.; Hong, Y.; Tao, M.; Hu, H.; Xu, M.; Guo, X.; Liu, Y. Human organoids in basic research and clinical applications. Signal Transduct. Target. Ther. 2022, 7, 168. [Google Scholar] [CrossRef] [PubMed]
- Eichmüller, O.L.; Knoblich, J.A. Human cerebral organoids—A new tool for clinical neurology research. Nat. Rev. Neurol. 2022, 18, 661–680. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zeng, P.-M.; Wu, J.; Luo, Z.-G. Advances and applications of brain organoids. Neurosci. Bull. 2023, 39, 1703–1716. [Google Scholar] [CrossRef]
- Mulder, L.A.; Depla, J.A.; Sridhar, A.; Wolthers, K.; Pajkrt, D.; Vieira de Sá, R. A beginner’s guide on the use of brain organoids for neuroscientists: A systematic review. Stem Cell Res. Ther. 2023, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Avansino, J.R.; Chen, D.C.; Hoagland, V.D.; Woolman, J.D.; Stelzner, M. Orthotopic transplantation of intestinal mucosal organoids in rodents. Surgery 2006, 140, 423–434. [Google Scholar] [CrossRef]
- Nie, Y.-Z.; Zheng, Y.-W.; Ogawa, M.; Miyagi, E.; Taniguchi, H. Human liver organoids generated with single donor-derived multiple cells rescue mice from acute liver failure. Stem Cell Res. Ther. 2018, 9, 5. [Google Scholar] [CrossRef]
- Lebreton, F.; Lavallard, V.; Bellofatto, K.; Bonnet, R.; Wassmer, C.H.; Perez, L.; Kalandadze, V.; Follenzi, A.; Boulvain, M.; Kerr-Conte, J. Insulin-producing organoids engineered from islet and amniotic epithelial cells to treat diabetes. Nat. Commun. 2019, 10, 4491. [Google Scholar] [CrossRef]
- van den Berg, C.W.; Ritsma, L.; Avramut, M.C.; Wiersma, L.E.; van den Berg, B.M.; Leuning, D.G.; Lievers, E.; Koning, M.; Vanslambrouck, J.M.; Koster, A.J. Renal subcapsular transplantation of PSC-derived kidney organoids induces neo-vasculogenesis and significant glomerular and tubular maturation in vivo. Stem Cell Rep. 2018, 10, 751–765. [Google Scholar] [CrossRef]
- McLelland, B.T.; Lin, B.; Mathur, A.; Aramant, R.B.; Thomas, B.B.; Nistor, G.; Keirstead, H.S.; Seiler, M.J. Transplanted hESC-derived retina organoid sheets differentiate, integrate, and improve visual function in retinal degenerate rats. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2586–2603. [Google Scholar] [CrossRef]
- Hsia, G.S.P.; Esposito, J.; da Rocha, L.A.; Ramos, S.L.G.; Okamoto, O.K. Clinical Application of Human Induced Pluripotent Stem Cell-Derived Organoids as an Alternative to Organ Transplantation. Stem Cells Int. 2021, 2021, 6632160. [Google Scholar] [CrossRef]
- Grade, S.; Götz, M. Neuronal replacement therapy: Previous achievements and challenges ahead. NPJ Regen. Med. 2017, 2, 29. [Google Scholar] [CrossRef]
- Mansour, A.A.; Gonçalves, J.T.; Bloyd, C.W.; Li, H.; Fernandes, S.; Quang, D.; Johnston, S.; Parylak, S.L.; Jin, X.; Gage, F.H. An in vivo model of functional and vascularized human brain organoids. Nat. Biotechnol. 2018, 36, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Daviaud, N.; Friedel, R.H.; Zou, H. Vascularization and engraftment of transplanted human cerebral organoids in mouse cortex. Eneuro 2018, 5, 0219-18. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.N.; Xu, T.Y.; Hong, C.; Cheng, M.H.; Zhu, P.X.; Lin, J.S.; Su, D.F.; Miao, C.Y. Cerebral organoids transplantation improves neurological motor function in rat brain injury. CNS Neurosci. Ther. 2020, 26, 682–697. [Google Scholar] [CrossRef]
- Bao, Z.; Fang, K.; Miao, Z.; Li, C.; Yang, C.; Yu, Q.; Zhang, C.; Miao, Z.; Liu, Y.; Ji, J. Human cerebral organoid implantation alleviated the neurological deficits of traumatic brain injury in mice. Oxidative Med. Cell. Longev. 2021, 2021, 6338722. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-N.; Wang, Z.; Xu, T.-Y.; Cheng, M.-H.; Li, W.-L.; Miao, C.-Y. Cerebral organoids repair ischemic stroke brain injury. Transl. Stroke Res. 2020, 11, 983–1000. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.-Y.; Yang, D.; Huang, Z.-Q.; Lin, Y.-H.; Wu, H.-Y.; Chang, L.; Luo, C.-X.; Xu, Y.; Liu, Y.; Zhu, D.-Y. Cerebral organoids transplantation repairs infarcted cortex and restores impaired function after stroke. NPJ Regen. Med. 2023, 8, 27. [Google Scholar] [CrossRef]
- Madrid, M.; Sumen, C.; Aivio, S.; Saklayen, N. Autologous induced pluripotent stem cell–based cell therapies: Promise, progress, and challenges. Curr. Protoc. 2021, 1, e88. [Google Scholar] [CrossRef]
- Revah, O.; Gore, F.; Kelley, K.W.; Andersen, J.; Sakai, N.; Chen, X.; Li, M.-Y.; Birey, F.; Yang, X.; Saw, N.L. Maturation and circuit integration of transplanted human cortical organoids. Nature 2022, 610, 319–326. [Google Scholar] [CrossRef]
- Jgamadze, D.; Lim, J.T.; Zhang, Z.; Harary, P.M.; Germi, J.; Mensah-Brown, K.; Adam, C.D.; Mirzakhalili, E.; Singh, S.; Gu, J.B. Structural and functional integration of human forebrain organoids with the injured adult rat visual system. Cell Stem Cell 2023, 30, 137–152.e7. [Google Scholar] [CrossRef]
- Pham, M.T.; Pollock, K.M.; Rose, M.D.; Cary, W.A.; Stewart, H.R.; Zhou, P.; Nolta, J.A.; Waldau, B. Generation of human vascularized brain organoids. Neuroreport 2018, 29, 588. [Google Scholar] [CrossRef]
- Shi, Y.; Sun, L.; Wang, M.; Liu, J.; Zhong, S.; Li, R.; Li, P.; Guo, L.; Fang, A.; Chen, R. Vascularized human cortical organoids (vOrganoids) model cortical development in vivo. PLoS Biol. 2020, 18, e3000705. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, 71. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; De Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef]
- Kitahara, T.; Sakaguchi, H.; Morizane, A.; Kikuchi, T.; Miyamoto, S.; Takahashi, J. Axonal extensions along corticospinal tracts from transplanted human cerebral organoids. Stem Cell Rep. 2020, 15, 467–481. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Xu, S.-B.; Chen, X.; Tao, M.; Tang, X.-Y.; Fang, K.-H.; Xu, M.; Pan, Y.; Chen, Y.; He, S. Human cerebral organoids establish subcortical projections in the mouse brain after transplantation. Mol. Psychiatry 2021, 26, 2964–2976. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.N.; Thunemann, M.; Liu, X.; Lu, Y.; Puppo, F.; Adams, J.W.; Kim, J.-H.; Ramezani, M.; Pizzo, D.P.; Djurovic, S. Multimodal monitoring of human cortical organoids implanted in mice reveal functional connection with visual cortex. Nat. Commun. 2022, 13, 7945. [Google Scholar] [CrossRef]
- Li, X.H.; Hu, N.; Chang, Z.H.; Shi, J.X.; Fan, X.; Chen, M.M.; Bao, S.Q.; Chen, C.; Zuo, J.C.; Zhang, X.W.; et al. Brain organoid maturation and implantation integration based on electrical signals input. J. Adv. Res. 2024, 73, 375–395. [Google Scholar] [CrossRef]
- Kim, J.T.; Kim, T.Y.; Youn, D.H.; Han, S.W.; Park, C.H.; Lee, Y.; Jung, H.; Rhim, J.K.; Park, J.J.; Ahn, J.H.; et al. Human embryonic stem cell-derived cerebral organoids for treatment of mild traumatic brain injury in a mouse model. Biochem. Biophys. Res. Commun. 2022, 635, 169–178. [Google Scholar] [CrossRef]
- Hu, N.; Shi, J.-X.; Chen, C.; Xu, H.-H.; Chang, Z.-H.; Hu, P.-F.; Guo, D.; Zhang, X.-W.; Shao, W.-W.; Fan, X. Constructing organoid-brain-computer interfaces for neurofunctional repair after brain injury. Nat. Commun. 2024, 15, 9580. [Google Scholar] [CrossRef]
- Cao, S.-Y.; Tao, M.-D.; Lou, S.-N.; Yang, D.; Lin, Y.-H.; Wu, H.-Y.; Chang, L.; Luo, C.-X.; Xu, Y.; Liu, Y. Functional reconstruction of the impaired cortex and motor function by hMGEOs transplantation in stroke. Biochem. Biophys. Res. Commun. 2023, 671, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Han, D.; Liu, W.; Wang, X.; Pan, N.; Wang, Y.; Chen, Z. Human iPSC-derived midbrain organoids functionally integrate into striatum circuits and restore motor function in a mouse model of Parkinson’s disease. Theranostics 2023, 13, 2673. [Google Scholar] [CrossRef]
- Huang, S.; Huang, F.; Zhang, H.; Yang, Y.; Lu, J.; Chen, J.; Shen, L.; Pei, G. In vivo development and single-cell transcriptome profiling of human brain organoids. Cell Prolif. 2022, 55, e13201. [Google Scholar] [CrossRef]
- Schafer, S.T.; Mansour, A.A.; Schlachetzki, J.C.; Pena, M.; Ghassemzadeh, S.; Mitchell, L.; Mar, A.; Quang, D.; Stumpf, S.; Ortiz, I.S. An in vivo neuroimmune organoid model to study human microglia phenotypes. Cell 2023, 186, 2111–2126.e2120. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, L.; Novak, S.W.; Yu, J.; Gallina, I.S.; Xu, L.L.; Lim, C.K.; Fernandes, S.; Shokhirev, M.N.; Williams, A.E. Morphological diversification and functional maturation of human astrocytes in glia-enriched cortical organoid transplanted in mouse brain. Nat. Biotechnol. 2024, 43, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Birey, F.; Li, M.-Y.; Revah, O.; Levy, R.; Thete, M.V.; Reis, N.; Kaganovsky, K.; Onesto, M.; Sakai, N. Antisense oligonucleotide therapeutic approach for Timothy syndrome. Nature 2024, 628, 818–825. [Google Scholar] [CrossRef]
- Xu, S.B.; Li, X.R.; Fan, P.; Li, X.; Hong, Y.; Han, X.; Wu, S.; Chu, C.; Chen, Y.; Xu, M. Single-Cell Transcriptome Landscape and Cell Fate Decoding in Human Brain Organoids after Transplantation. Adv. Sci. 2024, 11, 2402287. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-H.; Guo, D.; Chen, L.-Q.; Chang, Z.-H.; Shi, J.-X.; Hu, N.; Chen, C.; Zhang, X.-W.; Bao, S.-Q.; Chen, M.-M. Low-intensity ultrasound ameliorates brain organoid integration and rescues microcephaly deficits. Brain 2024, 147, 3817–3833. [Google Scholar] [CrossRef]
- Romito, A.; Cobellis, G. Pluripotent stem cells: Current understanding and future directions. Stem Cells Int. 2016, 2016, 9451492. [Google Scholar] [CrossRef]
- Narsinh, K.H.; Plews, J.; Wu, J.C. Comparison of human induced pluripotent and embryonic stem cells: Fraternal or identical twins? Mol. Ther. 2011, 19, 635–638. [Google Scholar] [CrossRef]
- Mallon, B.S.; Hamilton, R.S.; Kozhich, O.A.; Johnson, K.R.; Fann, Y.C.; Rao, M.S.; Robey, P.G. Comparison of the molecular profiles of human embryonic and induced pluripotent stem cells of isogenic origin. Stem Cell Res. 2014, 12, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Murillo, A.B.; Griesser, E.; Sinclair, L.; Davidson, L.; Prescott, A.; Singh, F.; Hogg, E.; Espejo-Serrano, C.; Jiang, H.; Yoshikawa, H. Proteomic and functional comparison between human induced and embryonic stem cells. Elife 2024, 13, RP92025. [Google Scholar]
- Barad, L.; Schick, R.; Zeevi-Levin, N.; Itskovitz-Eldor, J.; Binah, O. Human embryonic stem cells vs human induced pluripotent stem cells for cardiac repair. Can. J. Cardiol. 2014, 30, 1279–1287. [Google Scholar] [CrossRef]
- Riera, M.; Fontrodona, L.; Albert, S.; Ramirez, D.M.; Seriola, A.; Salas, A.; Muoz, Y.; Ramos, D.; Villegas-Perez, M.P.; Zapata, M.A. Comparative study of human embryonic stem cells (hESC) and human induced pluripotent stem cells (hiPSC) as a treatment for retinal dystrophies. Mol. Ther. Methods Clin. Dev. 2016, 3, 16010. [Google Scholar] [CrossRef]
- Kadoshima, T.; Sakaguchi, H.; Nakano, T.; Soen, M.; Ando, S.; Eiraku, M.; Sasai, Y. Self-organization of axial polarity, inside-out layer pattern, and species-specific progenitor dynamics in human ES cell–derived neocortex. Proc. Natl. Acad. Sci. USA 2013, 110, 20284–20289. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; O’Laughlin, R.; Song, H.; Ming, G.-l. Patterning of brain organoids derived from human pluripotent stem cells. Curr. Opin. Neurobiol. 2022, 74, 102536. [Google Scholar] [CrossRef]
- Bagley, J.A.; Reumann, D.; Bian, S.; Lévi-Strauss, J.; Knoblich, J.A. Fused cerebral organoids model interactions between brain regions. Nat. Methods 2017, 14, 743–751. [Google Scholar] [CrossRef]
- Birey, F.; Andersen, J.; Makinson, C.D.; Islam, S.; Wei, W.; Huber, N.; Fan, H.C.; Metzler, K.R.C.; Panagiotakos, G.; Thom, N. Assembly of functionally integrated human forebrain spheroids. Nature 2017, 545, 54–59. [Google Scholar] [CrossRef]
- Miura, Y.; Li, M.-Y.; Birey, F.; Ikeda, K.; Revah, O.; Thete, M.V.; Park, J.-Y.; Puno, A.; Lee, S.H.; Porteus, M.H. Generation of human striatal organoids and cortico-striatal assembloids from human pluripotent stem cells. Nat. Biotechnol. 2020, 38, 1421–1430. [Google Scholar] [CrossRef]
- Xiang, Y.; Tanaka, Y.; Patterson, B.; Kang, Y.-J.; Govindaiah, G.; Roselaar, N.; Cakir, B.; Kim, K.-Y.; Lombroso, A.P.; Hwang, S.-M. Fusion of regionally specified hPSC-derived organoids models human brain development and interneuron migration. Cell Stem Cell 2017, 21, 383–398.e387. [Google Scholar] [CrossRef]
- Sakaguchi, H.; Kadoshima, T.; Soen, M.; Narii, N.; Ishida, Y.; Ohgushi, M.; Takahashi, J.; Eiraku, M.; Sasai, Y. Generation of functional hippocampal neurons from self-organizing human embryonic stem cell-derived dorsomedial telencephalic tissue. Nat. Commun. 2015, 6, 8896. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Tanaka, Y.; Cakir, B.; Patterson, B.; Kim, K.-Y.; Sun, P.; Kang, Y.-J.; Zhong, M.; Liu, X.; Patra, P. hESC-derived thalamic organoids form reciprocal projections when fused with cortical organoids. Cell Stem Cell 2019, 24, 487–497.e487. [Google Scholar] [CrossRef]
- Qian, X.; Nguyen, H.N.; Song, M.M.; Hadiono, C.; Ogden, S.C.; Hammack, C.; Yao, B.; Hamersky, G.R.; Jacob, F.; Zhong, C. Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef] [PubMed]
- Tieng, V.; Stoppini, L.; Villy, S.; Fathi, M.; Dubois-Dauphin, M.; Krause, K.-H. Engineering of midbrain organoids containing long-lived dopaminergic neurons. Stem Cells Dev. 2014, 23, 1535–1547. [Google Scholar] [CrossRef]
- Muguruma, K.; Nishiyama, A.; Kawakami, H.; Hashimoto, K.; Sasai, Y. Self-organization of polarized cerebellar tissue in 3D culture of human pluripotent stem cells. Cell Rep. 2015, 10, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Eura, N.; Matsui, T.K.; Luginbühl, J.; Matsubayashi, M.; Nanaura, H.; Shiota, T.; Kinugawa, K.; Iguchi, N.; Kiriyama, T.; Zheng, C. Brainstem organoids from human pluripotent stem cells. Front. Neurosci. 2020, 14, 538. [Google Scholar] [CrossRef]
- Paşca, A.M.; Sloan, S.A.; Clarke, L.E.; Tian, Y.; Makinson, C.D.; Huber, N.; Kim, C.H.; Park, J.-Y.; O’rourke, N.A.; Nguyen, K.D. Functional cortical neurons and astrocytes from human pluripotent stem cells in 3D culture. Nat. Methods 2015, 12, 671–678. [Google Scholar] [CrossRef]
- Van De Leemput, J.; Boles, N.C.; Kiehl, T.R.; Corneo, B.; Lederman, P.; Menon, V.; Lee, C.; Martinez, R.A.; Levi, B.P.; Thompson, C.L. CORTECON: A temporal transcriptome analysis of in vitro human cerebral cortex development from human embryonic stem cells. Neuron 2014, 83, 51–68. [Google Scholar] [CrossRef]
- Monzel, A.S.; Smits, L.M.; Hemmer, K.; Hachi, S.; Moreno, E.L.; van Wuellen, T.; Jarazo, J.; Walter, J.; Brüggemann, I.; Boussaad, I. Derivation of human midbrain-specific organoids from neuroepithelial stem cells. Stem Cell Rep. 2017, 8, 1144–1154. [Google Scholar] [CrossRef]
- Di Lullo, E.; Kriegstein, A.R. The use of brain organoids to investigate neural development and disease. Nat. Rev. Neurosci. 2017, 18, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Zhao, M.; Chen, H.; Zhou, X.; Lenahan, C.; Ou, Y.; He, Y. The application of brain organoid technology in stroke research: Challenges and prospects. Front. Cell. Neurosci. 2021, 15, 646921. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.; Yoon, S.-J.; Tran, S.S.; Makinson, C.D.; Park, J.Y.; Andersen, J.; Valencia, A.M.; Horvath, S.; Xiao, X.; Huguenard, J.R. Long-term maturation of human cortical organoids matches key early postnatal transitions. Nat. Neurosci. 2021, 24, 331–342. [Google Scholar] [CrossRef]
- Fu, C.-L.; Dong, B.-C.; Jiang, X.; Li, D.; Yao, J. A cell therapy approach based on iPSC-derived midbrain organoids for the restoration of motor function in a Parkinson’s disease mouse model. Heliyon 2024, 10, e24234. [Google Scholar] [CrossRef]
- Choe, M.S.; Kim, S.J.; Oh, S.T.; Bae, C.M.; Choi, W.-Y.; Baek, K.M.; Kim, J.S.; Lee, M.Y. A simple method to improve the quality and yield of human pluripotent stem cell-derived cerebral organoids. Heliyon 2021, 7, e07350. [Google Scholar] [CrossRef]
- Qian, X.; Su, Y.; Adam, C.D.; Deutschmann, A.U.; Pather, S.R.; Goldberg, E.M.; Su, K.; Li, S.; Lu, L.; Jacob, F. Sliced human cortical organoids for modeling distinct cortical layer formation. Cell Stem Cell 2020, 26, 766–781.e769. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef]
- Jgamadze, D.; Harary, P.M.; Castellanos, M.; Blue, R.; Song, H.; Ming, G.-l.; Chen, H.I. Protocol for human brain organoid transplantation into a rat visual cortex to model neural repair. STAR Protoc. 2023, 4, 102470. [Google Scholar] [CrossRef]
- Kichula, E.A.; Huntley, G.W. Developmental and comparative aspects of posterior medial thalamocortical innervation of the barrel cortex in mice and rats. J. Comp. Neurol. 2008, 509, 239–258. [Google Scholar] [CrossRef]
- Kelley, K.W.; Revah, O.; Gore, F.; Kaganovsky, K.; Chen, X.; Deisseroth, K.; Pașca, S.P. Host circuit engagement of human cortical organoids transplanted in rodents. Nat. Protoc. 2024, 19, 3542–3567. [Google Scholar] [CrossRef]
- Datto, J.P.; Yang, J.; Dietrich, W.D.; Pearse, D.D. Does being female provide a neuroprotective advantage following spinal cord injury? Neural Regen. Res. 2015, 10, 1533–1536. [Google Scholar] [PubMed]
- Murphy, S.J.; McCullough, L.D.; Smith, J.M. Stroke in the female: Role of biological sex and estrogen. ILAR J. 2004, 45, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Espuny-Camacho, I.; Michelsen, K.A.; Gall, D.; Linaro, D.; Hasche, A.; Bonnefont, J.; Bali, C.; Orduz, D.; Bilheu, A.; Herpoel, A.; et al. Pyramidal Neurons Derived from Human Pluripotent Stem Cells Integrate Efficiently into Mouse Brain Circuits In Vivo. Neuron 2013, 77, 440–456. [Google Scholar] [CrossRef] [PubMed]
- Yousefifard, M.; Rahimi-Movaghar, V.; Nasirinezhad, F.; Baikpour, M.; Safari, S.; Saadat, S.; Jafari, A.M.; Asady, H.; Tousi, S.R.; Hosseini, M. Neural stem/progenitor cell transplantation for spinal cord injury treatment; A systematic review and meta-analysis. Neuroscience 2016, 322, 377–397. [Google Scholar] [CrossRef]
- Péron, S.; Droguerre, M.; Debarbieux, F.; Ballout, N.; Benoit-Marand, M.; Francheteau, M.; Brot, S.; Rougon, G.; Jaber, M.; Gaillard, A. A delay between motor cortex lesions and neuronal transplantation enhances graft integration and improves repair and recovery. J. Neurosci. 2017, 37, 1820–1834. [Google Scholar] [CrossRef]
- Yamagami, K.; Samata, B.; Doi, D.; Tsuchimochi, R.; Kikuchi, T.; Amimoto, N.; Ikeda, M.; Yoshimoto, K.; Takahashi, J. Progranulin enhances the engraftment of transplanted human iPS cell-derived cerebral neurons. Stem Cells Transl. Med. 2024, 13, 1113–1128. [Google Scholar] [CrossRef]
- Berríos-Torres, S.I.; Umscheid, C.A.; Bratzler, D.W.; Leas, B.; Stone, E.C.; Kelz, R.R.; Reinke, C.E.; Morgan, S.; Solomkin, J.S.; Mazuski, J.E. Centers for disease control and prevention guideline for the prevention of surgical site infection, 2017. JAMA Surg. 2017, 152, 784–791. [Google Scholar] [CrossRef]
- Ruan, J.; Yao, Y. Behavioral tests in rodent models of stroke. Brain Hemorrhages 2020, 1, 171–184. [Google Scholar] [CrossRef]
- Schaar, K.L.; Brenneman, M.M.; Savitz, S.I. Functional assessments in the rodent stroke model. Exp. Transl. Stroke Med. 2010, 2, 13. [Google Scholar] [CrossRef]
- Feeney, D.M.; Boyeson, M.G.; Linn, R.T.; Murray, H.M.; Dail, W.G. Responses to cortical injury: I. Methodology and local effects of contusions in the rat. Brain Res. 1981, 211, 67–77. [Google Scholar] [CrossRef]
- Zarruk, J.G.; Garcia-Yebenes, I.; Romera, V.G.; Ballesteros, I.; Moraga, A.; Cuartero, M.I.; Hurtado, O.; Sobrado, M.; Pradillo, J.M.; Fernandez-Lopez, D. Neurological tests for functional outcome assessment in rodent models of ischaemic stroke. Rev. Neurol. 2011, 53, 607–618. [Google Scholar]
- Lueptow, L.M. Novel object recognition test for the investigation of learning and memory in mice. J. Vis. Exp. JoVE 2017, 30, 55718. [Google Scholar]
- Müller, I.; Fendt, M. Fear conditioning in laboratory rodents. In Psychiatric Vulnerability, Mood, and Anxiety Disorders: Tests and Models in Mice and Rats; Springer: New York, NY, USA, 2022; pp. 119–160. [Google Scholar]
- Björklund, A.; Dunnett, S.B. The amphetamine induced rotation test: A re-assessment of its use as a tool to monitor motor impairment and functional recovery in rodent models of Parkinson’s disease. J. Park. Dis. 2019, 9, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Meredith, G.E.; Kang, U.J. Behavioral models of Parkinson’s disease in rodents: A new look at an old problem. Mov. Disord. 2006, 21, 1595–1606. [Google Scholar] [CrossRef]
- Lindvall, O.; Kokaia, Z. Stem cell research in stroke: How far from the clinic? Stroke 2011, 42, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Kokaia, Z.; Darsalia, V. Human neural stem cells for ischemic stroke treatment. In Human Neural Stem Cells: Results and Problems in Cell Differentiation; Springer: Cham, Switzerland, 2018; Volume 66, pp. 249–263. [Google Scholar]
- Díaz, L.; Zambrano, E.; Flores, M.E.; Contreras, M.; Crispín, J.C.; Alemán, G.; Bravo, C.; Armenta, A.; Valdés, V.J.; Tovar, A. Ethical considerations in animal research: The principle of 3R’s. Rev. De Investig. Clin. 2021, 73, 199–209. [Google Scholar] [CrossRef]
- Chen, H.I.; Wolf, J.A.; Blue, R.; Song, M.M.; Moreno, J.D.; Ming, G.-l.; Song, H. Transplantation of human brain organoids: Revisiting the science and ethics of brain chimeras. Cell Stem Cell 2019, 25, 462–472. [Google Scholar] [CrossRef]
- Kataoka, M.; Gyngell, C.; Savulescu, J.; Sawai, T. The ethics of human brain organoid transplantation in animals. Neuroethics 2023, 16, 27. [Google Scholar] [CrossRef]
- Azevedo, F.A.; Carvalho, L.R.; Grinberg, L.T.; Farfel, J.M.; Ferretti, R.E.; Leite, R.E.; Filho, W.J.; Lent, R.; Herculano-Houzel, S. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J. Comp. Neurol. 2009, 513, 532–541. [Google Scholar] [CrossRef]
- Greene, M.; Schill, K.; Takahashi, S.; Bateman-House, A.; Beauchamp, T.; Bok, H.; Cheney, D.; Coyle, J.; Deacon, T.; Dennett, D. Moral issues of human-non-human primate neural grafting. Science 2005, 309, 385–386. [Google Scholar] [CrossRef]
- Sawai, T.; Hayashi, Y.; Niikawa, T.; Shepherd, J.; Thomas, E.; Lee, T.-L.; Erler, A.; Watanabe, M.; Sakaguchi, H. Mapping the ethical issues of brain organoid research and application. AJOB Neurosci. 2022, 13, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Erler, A. Human Brain Organoid Transplantation: Testing the Foundations of Animal Research Ethics. Neuroethics 2024, 17, 20. [Google Scholar] [CrossRef]
- Hyun, I. From naïve pluripotency to chimeras: A new ethical challenge? Development 2015, 142, 6–8. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. The Emerging Field of Human NEURAL Organoids, Transplants, and Chimeras: Science, Ethics, and Governance; The National Academies Press: Washington, DC, USA, 2021. [Google Scholar]
- Wang, M.; Gage, F.H.; Schafer, S.T. Transplantation strategies to enhance maturity and cellular complexity in brain organoids. Biol. Psychiatry 2023, 93, 616–621. [Google Scholar] [CrossRef]
- Ormel, P.R.; Vieira de Sá, R.; Van Bodegraven, E.J.; Karst, H.; Harschnitz, O.; Sneeboer, M.A.; Johansen, L.E.; van Dijk, R.E.; Scheefhals, N.; Berdenis van Berlekom, A. Microglia innately develop within cerebral organoids. Nat. Commun. 2018, 9, 4167. [Google Scholar] [CrossRef]
- Sun, X.-Y.; Ju, X.-C.; Li, Y.; Zeng, P.-M.; Wu, J.; Zhou, Y.-Y.; Shen, L.-B.; Dong, J.; Chen, Y.-J.; Luo, Z.-G. Generation of vascularized brain organoids to study neurovascular interactions. Elife 2022, 11, e76707. [Google Scholar] [CrossRef] [PubMed]
- Dao, L.; You, Z.; Lu, L.; Xu, T.; Sarkar, A.K.; Zhu, H.; Liu, M.; Calandrelli, R.; Yoshida, G.; Lin, P. Modeling blood-brain barrier formation and cerebral cavernous malformations in human PSC-derived organoids. Cell Stem Cell 2024, 31, 818–833.e811. [Google Scholar] [CrossRef]
- Matsui, T.K.; Tsuru, Y.; Hasegawa, K.; Kuwako, K.-i. Vascularization of human brain organoids. Stem Cells 2021, 39, 1017–1024. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, J.; Xu, Z.; Yan, H.; Tang, B.; Liu, C.; Chen, C.; Meng, Q. Microglia-containing human brain organoids for the study of brain development and pathology. Mol. Psychiatry 2023, 28, 96–107. [Google Scholar] [CrossRef]
- Madhavan, M.; Nevin, Z.S.; Shick, H.E.; Garrison, E.; Clarkson-Paredes, C.; Karl, M.; Clayton, B.L.; Factor, D.C.; Allan, K.C.; Barbar, L. Induction of myelinating oligodendrocytes in human cortical spheroids. Nat. Methods 2018, 15, 700–706. [Google Scholar] [CrossRef]
- Shaker, M.R.; Pietrogrande, G.; Martin, S.; Lee, J.-H.; Sun, W.; Wolvetang, E.J. Rapid and efficient generation of myelinating human oligodendrocytes in organoids. Front. Cell. Neurosci. 2021, 15, 631548. [Google Scholar] [CrossRef] [PubMed]
- Paşca, S.P. Assembling human brain organoids. Science 2019, 363, 126–127. [Google Scholar] [CrossRef]
- Ao, Z.; Cai, H.; Havert, D.J.; Wu, Z.; Gong, Z.; Beggs, J.M.; Mackie, K.; Guo, F. One-stop microfluidic assembly of human brain organoids to model prenatal cannabis exposure. Anal. Chem. 2020, 92, 4630–4638. [Google Scholar] [CrossRef]
- Jo, J.; Xiao, Y.; Sun, A.X.; Cukuroglu, E.; Tran, H.-D.; Göke, J.; Tan, Z.Y.; Saw, T.Y.; Tan, C.-P.; Lokman, H. Midbrain-like organoids from human pluripotent stem cells contain functional dopaminergic and neuromelanin-producing neurons. Cell Stem Cell 2016, 19, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.-N.; Jin, Y.; An, Y.; Kim, J.; Choi, Y.S.; Lee, J.S.; Kim, J.; Choi, W.-Y.; Koo, D.-J.; Yu, W. Microfluidic device with brain extracellular matrix promotes structural and functional maturation of human brain organoids. Nat. Commun. 2021, 12, 4730. [Google Scholar] [CrossRef]
- Giandomenico, S.L.; Mierau, S.B.; Gibbons, G.M.; Wenger, L.M.; Masullo, L.; Sit, T.; Sutcliffe, M.; Boulanger, J.; Tripodi, M.; Derivery, E. Cerebral organoids at the air–liquid interface generate diverse nerve tracts with functional output. Nat. Neurosci. 2019, 22, 669–679. [Google Scholar] [CrossRef]
- Charles, S.; Jackson-Holmes, E.; Sun, G.; Zhou, Y.; Siciliano, B.; Niu, W.; Han, H.; Nikitina, A.; Kemp, M.L.; Wen, Z. Non-Invasive Quality Control of Organoid Cultures Using Mesofluidic CSTR Bioreactors and High-Content Imaging. Adv. Mater. Technol. 2025, 5, 2400473. [Google Scholar] [CrossRef]
- Vives, J.; Batlle-Morera, L. The challenge of developing human 3D organoids into medicines. Stem Cell Res. Ther. 2020, 11, 72. [Google Scholar] [CrossRef]
- Dossena, M.; Piras, R.; Cherubini, A.; Barilani, M.; Dugnani, E.; Salanitro, F.; Moreth, T.; Pampaloni, F.; Piemonti, L.; Lazzari, L. Standardized GMP-compliant scalable production of human pancreas organoids. Stem Cell Res. Ther. 2020, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- van Zanten, J.; Jorritsma-Smit, A.; Westra, H.; Baanstra, M.; de Bruin-Jellema, A.; Allersma, D.; Gareb, B.; Coppes, R.P. Optimization of the Production Process of Clinical-Grade Human Salivary Gland Organoid-Derived Cell Therapy for the Treatment of Radiation-Induced Xerostomia in Head and Neck Cancer. Pharmaceutics 2024, 16, 435. [Google Scholar] [CrossRef]
- Ahn, S.-J.; Lee, S.; Kwon, D.; Oh, S.; Park, C.; Jeon, S.; Lee, J.H.; Kim, T.S.; Oh, I.U. Essential guidelines for manufacturing and application of organoids. Int. J. Stem Cells 2024, 17, 102. [Google Scholar] [CrossRef] [PubMed]
- Cobb, H.; Aparicio-Domingo, S.; Canto-Soler, M.V. Transitioning into GMP-compliance: Alternative methods for producing retinal organoids for transplantation. Transl. Vis. Sci. Technol. 2021, 10, 9. [Google Scholar] [CrossRef]
- Tomaskovic-Crook, E.; Crook, J.M. Clinically amendable, defined, and rapid induction of human brain organoids from induced pluripotent stem cells. In Organoids: Stem Cells, Structure, and Function; Humana: New York, NY, USA, 2019; pp. 13–22. [Google Scholar]
- Mohamed, N.-V.; Lépine, P.; Lacalle-Aurioles, M.; Sirois, J.; Mathur, M.; Reintsch, W.; Beitel, L.K.; Fon, E.A.; Durcan, T.M. Microfabricated disk technology: Rapid scale up in midbrain organoid generation. Methods 2022, 203, 465–477. [Google Scholar] [CrossRef]
- Zeiss, C.J. Comparative milestones in rodent and human postnatal central nervous system development. Toxicol. Pathol. 2021, 49, 1368–1373. [Google Scholar] [CrossRef] [PubMed]
- Urrestizala-Arenaza, N.; Cerchio, S.; Cavaliere, F.; Magliaro, C. Limitations of human brain organoids to study neurodegenerative diseases: A manual to survive. Front. Cell. Neurosci. 2024, 18, 1419526. [Google Scholar] [CrossRef]
- Grenier, K.; Kao, J.; Diamandis, P. Three-dimensional modeling of human neurodegeneration: Brain organoids coming of age. Mol. Psychiatry 2020, 25, 254–274. [Google Scholar] [CrossRef]
- Hossain, M.K.; Kim, H.-R.; Chae, H.J. Aging phenotype in AD brain organoids: Track to success and challenges. Ageing Res. Rev. 2024, 96, 102256. [Google Scholar] [CrossRef] [PubMed]
- Saboori, M.; Riazi, A.; Taji, M.; Yadegarfar, G. Traumatic brain injury and stem cell treatments: A review of recent 10 years clinical trials. Clin. Neurol. Neurosurg. 2024, 239, 108219. [Google Scholar] [CrossRef]
- Cabral, D.F.; Fried, P.; Koch, S.; Rice, J.; Rundek, T.; Pascual-Leone, A.; Sacco, R.; Wright, C.B.; Gomes-Osman, J. Efficacy of mechanisms of neuroplasticity after a stroke. Restor. Neurol. Neurosci. 2022, 40, 73–84. [Google Scholar] [CrossRef]
- Darsalia, V.; Allison, S.J.; Cusulin, C.; Monni, E.; Kuzdas, D.; Kallur, T.; Lindvall, O.; Kokaia, Z. Cell number and timing of transplantation determine survival of human neural stem cell grafts in stroke-damaged rat brain. J. Cereb. Blood Flow Metab. 2011, 31, 235–242. [Google Scholar] [CrossRef]
- Ya, J.; Pellumbaj, J.; Hashmat, A.; Bayraktutan, U. The role of stem cells as therapeutics for ischaemic stroke. Cells 2024, 13, 112. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.S., Jr.; Hetz, R.A.; Liao, G.P.; Aertker, B.M.; Ewing-Cobbs, L.; Juranek, J.; Savitz, S.I.; Jackson, M.L.; Romanowska-Pawliczek, A.M.; Triolo, F. Treatment of severe adult traumatic brain injury using bone marrow mononuclear cells. Stem Cells 2017, 35, 1065–1079. [Google Scholar] [CrossRef]
- Eigenhuis, K.N.; Somsen, H.B.; van der Kroeg, M.; Smeenk, H.; Korporaal, A.L.; Kushner, S.A.; de Vrij, F.M.; van den Berg, D.L. A simplified protocol for the generation of cortical brain organoids. Front. Cell. Neurosci. 2023, 17, 1114420. [Google Scholar] [CrossRef] [PubMed]
- Lindborg, B.A.; Brekke, J.H.; Vegoe, A.L.; Ulrich, C.B.; Haider, K.T.; Subramaniam, S.; Venhuizen, S.L.; Eide, C.R.; Orchard, P.J.; Chen, W. Rapid induction of cerebral organoids from human induced pluripotent stem cells using a chemically defined hydrogel and defined cell culture medium. Stem Cells Transl. Med. 2016, 5, 970–979. [Google Scholar] [CrossRef]
- Ding, L.; Hartung, T.; Smirnova, L. Brain organoids on demand: Cryopreservation as a solution. Cryobiology 2023, 113, 104635. [Google Scholar] [CrossRef]
- Xue, W.; Li, H.; Xu, J.; Yu, X.; Liu, L.; Liu, H.; Zhao, R.; Shao, Z. Effective cryopreservation of human brain tissue and neural organoids. Cell Rep. Methods 2024, 4, 100777. [Google Scholar] [CrossRef]
- McGonigle, P.; Ruggeri, B. Animal models of human disease: Challenges in enabling translation. Biochem. Pharmacol. 2014, 87, 162–171. [Google Scholar] [CrossRef]
- Kataoka, M.; Niikawa, T.; Nagaishi, N.; Lee, T.-L.; Erler, A.; Savulescu, J.; Sawai, T. Beyond consciousness: Ethical, legal, and social issues in human brain organoid research and application. Eur. J. Cell Biol. 2024, 104, 151470. [Google Scholar] [CrossRef]
- de Jongh, D.; Massey, E.K.; Bunnik, E.M. Organoids: A systematic review of ethical issues. Stem Cell Res. Ther. 2022, 13, 337. [Google Scholar] [CrossRef]
- Farhang Doost, N.; Srivastava, S.K. A comprehensive review of organ-on-a-chip technology and its applications. Biosensors 2024, 14, 225. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, N.; Bian, S.; Sawan, M. Brain organoids-on-chip for neural diseases modeling: History, challenges and trends. J. Pharm. Anal. 2025, 101323. [Google Scholar] [CrossRef]
- Huh, D.; Matthews, B.D.; Mammoto, A.; Montoya-Zavala, M.; Hsin, H.Y.; Ingber, D.E. Reconstituting organ-level lung functions on a chip. Science 2010, 328, 1662–1668. [Google Scholar] [CrossRef] [PubMed]
- Papamichail, L.; Koch, L.S.; Veerman, D.; Broersen, K.; van der Meer, A.D. Organoids-on-a-chip: Microfluidic technology enables culture of organoids with enhanced tissue function and potential for disease modeling. Front. Bioeng. Biotechnol. 2025, 13, 1515340. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, L.; Yu, H.; Yin, F.; Wang, Y.; Liu, H.; Jiang, L.; Qin, J. In situ generation of human brain organoids on a micropillar array. Lab Chip 2017, 17, 2941–2950. [Google Scholar] [CrossRef] [PubMed]
- Saorin, G.; Caligiuri, I.; Rizzolio, F. Microfluidic organoids-on-a-chip: The future of human models. Semin. Cell Dev. Biol. 2023, 144, 41–54. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Guo, Y.; Zhu, Y.; Qin, J. Engineering stem cell-derived 3D brain organoids in a perfusable organ-on-a-chip system. RSC Adv. 2018, 8, 1677–1685. [Google Scholar] [CrossRef]
- Castiglione, H.; Vigneron, P.-A.; Baquerre, C.; Yates, F.; Rontard, J.; Honegger, T. Human brain organoids-on-chip: Advances, challenges, and perspectives for preclinical applications. Pharmaceutics 2022, 14, 2301. [Google Scholar] [CrossRef]
- Berger, E.; Magliaro, C.; Paczia, N.; Monzel, A.S.; Antony, P.; Linster, C.L.; Bolognin, S.; Ahluwalia, A.; Schwamborn, J.C. Millifluidic culture improves human midbrain organoid vitality and differentiation. Lab Chip 2018, 18, 3172–3183. [Google Scholar] [CrossRef]
- Salmon, I.; Grebenyuk, S.; Fattah, A.R.A.; Rustandi, G.; Pilkington, T.; Verfaillie, C.; Ranga, A. Engineering neurovascular organoids with 3D printed microfluidic chips. Lab Chip 2022, 22, 1615–1629. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, X.; Sun, L.; Wang, Y.; Zhao, Y. Engineering human brain assembloids by microfluidics. Adv. Mater. 2023, 35, 2210083. [Google Scholar] [CrossRef]
- Soscia, D.A.; Lam, D.; Tooker, A.C.; Enright, H.A.; Triplett, M.; Karande, P.; Peters, S.K.; Sales, A.P.; Wheeler, E.K.; Fischer, N.O. A flexible 3-dimensional microelectrode array for in vitro brain models. Lab Chip 2020, 20, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.; Di Lisa, D.; Giacomozzi, F.; Cian, A.; Giubertoni, D.; Martinoia, S.; Lorenzelli, L. Development of multi-depth probing 3D microelectrode array to record electrophysiological activity within neural cultures. J. Micromech. Microeng. 2023, 33, 115002. [Google Scholar] [CrossRef]
- Park, Y.; Franz, C.K.; Ryu, H.; Luan, H.; Cotton, K.Y.; Kim, J.U.; Chung, T.S.; Zhao, S.; Vazquez-Guardado, A.; Yang, D.S. Three-dimensional, multifunctional neural interfaces for cortical spheroids and engineered assembloids. Sci. Adv. 2021, 7, eabf9153. [Google Scholar] [CrossRef]
- Del Rio, J.S.; Ro, J.; Yoon, H.; Park, T.-E.; Cho, Y.-K. Integrated technologies for continuous monitoring of organs-on-chips: Current challenges and potential solutions. Biosens. Bioelectron. 2023, 224, 115057. [Google Scholar]
- Karzbrun, E.; Kshirsagar, A.; Cohen, S.R.; Hanna, J.H.; Reiner, O. Human brain organoids on a chip reveal the physics of folding. Nat. Phys. 2018, 14, 515–522. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Zhu, Y.; Qin, J. Human brain organoid-on-a-chip to model prenatal nicotine exposure. Lab Chip 2018, 18, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Wang, Y.; Zhu, Y.; Tao, T.; Yin, F.; Guo, Y.; Liu, H.; Li, F.; Wang, P.; Chen, Y. Neurodevelopmental impairment induced by prenatal valproic acid exposure shown with the human cortical organoid-on-a-chip model. Microsyst. Nanoeng. 2020, 6, 49. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Chen, W.; Cao, R.; Xie, Y.; Wang, P.; Wu, Y.; Wang, Y.; Qin, J. Brain organoid-on-chip system to study the effects of breast cancer derived exosomes on the neurodevelopment of brain. Cell Regen. 2022, 11, 7. [Google Scholar] [CrossRef]
- Cai, H.; Ao, Z.; Tian, C.; Wu, Z.; Liu, H.; Tchieu, J.; Gu, M.; Mackie, K.; Guo, F. Brain organoid reservoir computing for artificial intelligence. Nat. Electron. 2023, 6, 1032–1039. [Google Scholar] [CrossRef]
- Barker, R.A.; Lao-Kaim, N.P.; Guzman, N.V.; Athauda, D.; Bjartmarz, H.; Björklund, A.; Church, A.; Cutting, E.; Daft, D.; Dayal, V. The TransEuro open-label trial of human fetal ventral mesencephalic transplantation in patients with moderate Parkinson’s disease. Nat. Biotechnol. 2025, 1–9. [Google Scholar] [CrossRef]
- Sawamoto, N.; Doi, D.; Nakanishi, E.; Sawamura, M.; Kikuchi, T.; Yamakado, H.; Taruno, Y.; Shima, A.; Fushimi, Y.; Okada, T. Phase I/II trial of iPS-cell-derived dopaminergic cells for Parkinson’s disease. Nature 2025, 641, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.J.; Bock, M.; Zhang, S.; An, S.B.; Han, I. Therapeutic Transplantation of Human Central Nervous System Organoids for Neural Reconstruction. Int. J. Mol. Sci. 2024, 25, 8540. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; Lombardozzi, G.; Ammannito, F.; Scenna, M.S.; Maceroni, E.; Quintiliani, M.; d’Angelo, M.; Cimini, A.; Castelli, V. Brain organoids: A game-changer for drug testing. Pharmaceutics 2024, 16, 443. [Google Scholar] [CrossRef] [PubMed]
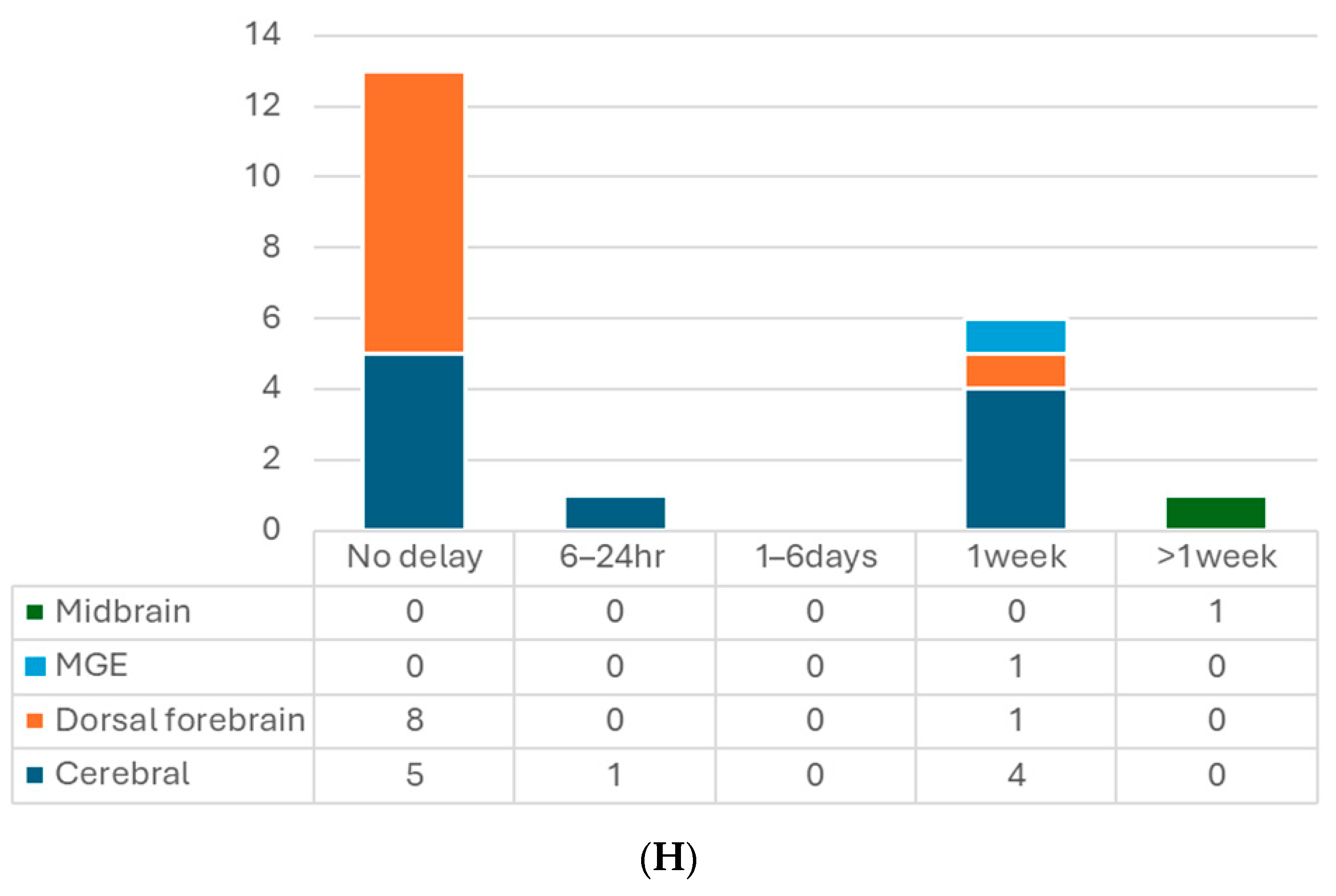
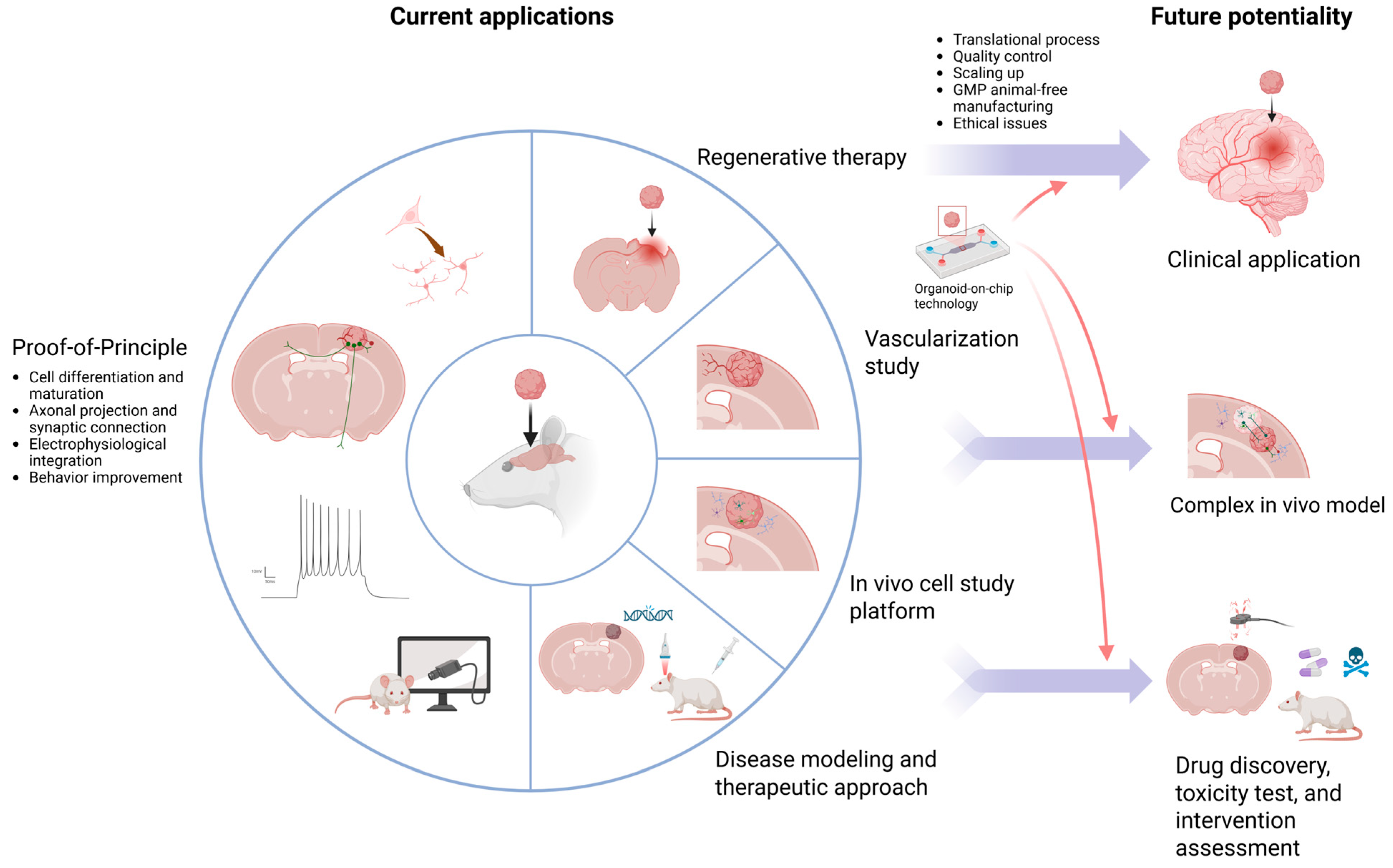

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PubMed | Embase |
|---|---|
| (“Organoids” [Mesh] OR spheroid OR assembloid) AND (Brain [Mesh] OR cerebr* OR cortical OR forebrain OR hindbrain OR midbrain OR striatal OR thalam* OR hypothalam* OR cerebel* OR ganglionic eminence [Mesh]) AND (“Transplantation” [Mesh] OR “Transplants” [Mesh] OR implant* OR graft) | ((Brain or cerebr* or cortical or forebrain or hindbrain or midbrain or striatal or thalam* or hypothalam* or cerebel* or ganglionic eminence) organoid OR ((brain or cerebr* or cerebel* or cortical*) spheroid) OR brain assembloid) AND (transplant* or implant* or graft) |
| Proof-of-Principle | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Transplanted Organoids | Cell Type Derived | Extracellular Scaffold During Organoid Culture | Receiver Age or Size | Immunosuppressive Treatment | Brain Injury Site | Transplant Timing | Evaluation Time Post Implantation | Main Findings and Measurements | References |
|
| Matrigel |
| None |
| Immediately after injury | 0.5–8 months |
| Mansour et al., 2018 [12] |
|
| Matrigel |
| None |
| Immediately after injury | 2 and 4 weeks |
| Daviaud et al., 2018 [13] |
|
| ND |
| None |
| No delay or 1 week after injury | 12 weeks |
| Kitahara et al., 2020 [25] |
| Tacrolimus hydrate |
| 1 week after injury | ||||||
|
| ND |
| None |
| N/A | 1–5 months |
| Dong et al., 2021 [26] |
|
(control, TS) | ND |
| None |
| N/A | 3–8 months |
| Revah et al., 2022 [19] |
|
| ND |
| None |
| Immediately after injury | 8–11 weeks |
| Wilson et al., 2022 [27] |
|
| Matrigel |
| Cyclosporine A |
| Immediately after injury | 3 months |
| Jgamadze et al., 2023 [20] |
|
| ND |
| None |
| Immediately after injury | months |
| Li et al., 2024 [28] |
| Therapeutic-Traumatic Brain Injury | |||||||||
| Transplanted Organoids | Cell Type Derived | Extracellular Scaffold | Receiver Age or Size | Immunosuppressive Treatment | Brain Injury Site | Transplant Timing | Evaluation Time Post Implantation | Main findings and Measurements | References |
|
| Matrigel |
| Cyclosporine A |
| Immediately after injury | 8 weeks |
| Wang Z. et al., 2020 [14] |
|
| Matrigel |
| None |
| 1 week after injury | 70 days |
| Bao et al., 2021 [15] |
|
| Matrigel |
| Cyclosporine A |
| 1 week after injury | 7 and 14 days |
| Kim et al., 2022 [29] |
|
| ND |
| None |
| Immediately after injury | 40–180 days |
| Hu et al., 2024 [30] |
| Therapeutic-Stroke | |||||||||
| Transplanted Organoids | Cell type Derived | Extracellular Scaffold | Receiver Age or Size | Immunosuppressive Treatment | Brain Injury Site | Transplant Timing | Evaluation Time Post Implantation | Main Findings and Measurements | References |
|
| Matrigel |
| Cyclosporine A |
| 6 h, 24 h, or 7 days after MCAO | 4 weeks |
| Wang S. N. et al., 2020 [16] |
|
| ND |
| None |
| 1 week post-stroke | 180 days |
| Cao et al., 2023 [17] |
|
| ND |
| None |
| 1 week post-stroke | 150 days |
| Cao et al., 2023 [31] |
| Therapeutic-Parkinson’s Disease | |||||||||
| Transplanted Organoids | Cell Type Derived | Extracellular Scaffold | Receiver Age or Size | Immunosuppressive Treatment | Brain Injury Site | Transplant Timing | Evaluation Time Post Implantation | Main Findings and Measurements | References |
|
| Matrigel |
| None |
| 4-week post-6-OHDA treatment | 6, 12, and 16 weeks |
| Zheng et al., 2023 [32] |
| Vascularization | |||||||||
| Transplanted Organoids | Cell Type Derived | Extracellular Scaffold | Receiver Age or Size | Immunosuppressive Treatment | Brain Injury Site | Transplant Timing | Evaluation Time Post Implantation | Main Findings and Measurements | References |
|
| Matrigel |
| None |
| Immediately after injury | 2 weeks |
| Pham, M. T. et al., 2018 [21] |
|
| Matrigel |
| None |
| Immediately after injury | 3 days to 2 months |
| Shi, Y. et al., 2020 [22] |
| Disease Modeling and Cell Study Platform In Vivo | |||||||||
| Transplanted Organoids | Cell Type Derived | Extracellular Scaffold | Receiver Age or Size | Immunosuppressive Treatment | Brain Injury Site | Transplant Timing | Evaluation Time Post Implantation | Main Findings and Measurements | References |
|
| Matrigel |
| None |
| N/A | 2 months |
| Huang et al., 2022 [33] |
|
| Matrigel |
| None |
| Immediately after injury | 6–12 weeks |
| Schafer et al., 2023 [34] |
|
| Matrigel |
| None |
| Immediately after injury | 5, 6, and 8 months |
| Wang et al., 2024 [35] |
|
| ND |
| None |
| N/A | 14 days |
| Chen et al., 2024 [36] |
|
| ND |
| None |
| N/A | 2–4 months |
| Xu et al., 2024 [37] |
|
| ND |
| None |
| Immediately after injury | 2–5 months |
| Li et al., 2024 [38] |
| Proof-of-Principle | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sequence Generation | Baseline Characteristics | Allocation Concealment | Random Housing | Performance Blinding | Random Outcome Assessment | Detection Blinding | Incomplete Outcome Data | Selective Outcome Reporting | Other Sources of Bias | |
| Mansour et al., 2018 [12] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | High Risk | High Risk | Low Risk | Low Risk | Low Risk |
| Daviaud et al., 2018 [13] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Low Risk | Low Risk |
| Kitahara et al., 2020 [25] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Low Risk | Low Risk |
| Dong et al., 2021 [26] | Low Risk | Unclear Risk | Low Risk | Unclear Risk | Low Risk | Low Risk | Low Risk | Low Risk | Low Risk | Low Risk |
| Revah et al., 2022 [19] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Low Risk | Unclear Risk | Low Risk | Low Risk | Low Risk | Low Risk |
| Wilson et al., 2022 [27] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Low Risk | Low Risk | Low Risk |
| Jgamadze et al., 2023 [20] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Low Risk | Low Risk | Low Risk |
| Li et al., 2024 [28] | Low Risk * | Unclear Risk | Low Risk | Unclear Risk | Low Risk | Low Risk | Low Risk | Low Risk | Low Risk | Low Risk |
| Therapeutic—Traumatic Brain Injury | ||||||||||
| Sequence Generation | Baseline Characteristics | Allocation Concealment | Random Housing | Performance Blinding | Random Outcome Assessment | Detection Blinding | Incomplete Outcome Data | Selective Outcome Reporting | Other Sources of Bias | |
| Wang Z. et al., 2020 [14] | Low Risk * | Low Risk | Low Risk | Unclear Risk | Low Risk | Low Risk | Low Risk | Low Risk | Low Risk | Low Risk |
| Bao et al., 2021 [15] | Low Risk * | Unclear Risk | Low Risk | Unclear Risk | Unclear Risk | Unclear risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Kim et al., 2022 [29] | Low Risk * | Unclear Risk | Low Risk | Unclear Risk | Unclear Risk | Unclear Risk | Low Risk | Low Risk | Low Risk | Low Risk |
| Hu et al., 2024 [30] | Low Risk * | Unclear Risk | Low Risk | Unclear Risk | Low Risk | Low Risk | Low Risk | Unclear risk | Low Risk | Low Risk |
| Therapeutic—Stroke | ||||||||||
| Sequence Generation | Baseline Characteristics | Allocation Concealment | Random Housing | Performance Blinding | Random Outcome Assessment | Detection Blinding | Incomplete Outcome Data | Selective Outcome Reporting | Other Sources of Bias | |
| Wang S. N. et al., 2020 [16] | Low Risk * | Low Risk | Low Risk | Unclear Risk | Low Risk | Low Risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Cao et al., 2023 [17] | Unclear Risk | Low Risk | Unclear risk | Unclear Risk | Low Risk | Unclear risk | Low Risk | Low Risk | Low Risk | Low Risk |
| Cao et al., 2023 [31] | Low Risk * | Low Risk | Low Risk | Unclear Risk | Low Risk | Low Risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Therapeutic—Parkinson’s Disease | ||||||||||
| Sequence Generation | Baseline Characteristics | Allocation Concealment | Random Housing | Performance Blinding | Random Outcome Assessment | Detection Blinding | Incomplete Outcome Data | Selective Outcome Reporting | Other Sources of Bias | |
| Zheng et al., 2023 [32] | Unclear Risk | Low Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear risk | Unclear risk | Unclear Risk | Low Risk | Low Risk |
| Vascularization | ||||||||||
| Sequence Generation | Baseline Characteristics | Allocation Concealment | Random Housing | Performance Blinding | Random Outcome Assessment | Detection Blinding | Incomplete Outcome Data | Selective Outcome Reporting | Other Sources of Bias | |
| Pham, M. T. et al., 2018 [21] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear risk | Unclear risk | Low Risk | Low Risk | High Risk |
| Shi, Y. et al., 2020 [22] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Disease Modeling and Cell Study Platform In Vivo | ||||||||||
| Sequence Generation | Baseline Characteristics | Allocation Concealment | Random Housing | Performance Blinding | Random Outcome Assessment | Detection Blinding | Incomplete Outcome Data | Selective Outcome Reporting | Other Sources of Bias | |
| Huang et al. 2022 [33] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Schafer et al., 2023 [34] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | Unclear risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Wang et al., 2024 [35] | Low Risk * | Unclear Risk | Low Risk | Unclear Risk | Unclear Risk | Low Risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Chen et al., 2024 [36] | Unclear Risk | Unclear Risk | Unclear Risk | Unclear Risk | High Risk | Unclear risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Xu et al., 2024 [37] | Low Risk * | Unclear Risk | Low Risk | Unclear Risk | Unclear Risk | Low Risk | Unclear risk | Low Risk | Low Risk | Low Risk |
| Li et al., 2024 [38] | Low Risk * | Unclear Risk | Low Risk | Unclear Risk | Unclear Risk | Low Risk | Low Risk | Low Risk | Low Risk | Low Risk |
| Proof-of-Principle | |||||
|---|---|---|---|---|---|
| Animal | Location | Lesion Created Method and Size or Delivery Method | Fix and Cover | Immunosuppressive Treatment | |
| Mansour et al., 2018 [12] | SCID mice |
|
| Covered by 5 mm cover slip sealed with adhesive glue | None |
| Daviaud et al., 2018 [13] | CD 1 mice (newborn) |
|
| Covered by Bone flap sealed with fibrin glue | None |
| Kitahara et al., 2020 [25] | SCID mice (newborn and adult) |
|
| Returned the hinged bone flap | None |
| Cynomolgus monkeys |
|
| ND | Tacrolimus hydrate | |
| Dong et al., 2021 [26] | SCID mice |
|
| ND | None |
| Revah et al., 2022 [19] | Athymic rats (newborn) |
|
| ND | None |
| Wilson et al., 2022 [27] | SCID mice |
|
| Covered by a glass plug with 16 channels microelectrode array sealed with dental adhesive | None |
| Jgamadze et al., 2023 [20] | LE rats |
|
| Covered by PDMS cranioplasty cap sealed with bone cement PMMA | Cyclosporine A |
| Li et al., 2024 [28] | SCID mice |
|
| Returned the hinged bone flap | None |
| Therapeutic—Traumatic Brain Injury | |||||
| Animal | Location | Lesion Created Method and Size or Delivery Method | Fix and Cover | Immunosuppressive Treatment | |
| Wang Z. et al., 2020 [14] | SD rats |
|
| Sealed with piece of skull and bone wax. | Cyclosporine A |
| Bao et al., 2021 [15] | SCID mice |
|
| ND | None |
| Kim et al., 2022 [29] | C57BL/6J mice |
|
| Returned the hinged bone flap and sealed with fibrin glue | Cyclosporine A |
| Hu et al., 2024 [30] | SCID mice |
|
| Returned the hinged bone flap | None |
| Therapeutic—Stroke | |||||
| Animal | Model and Location | Lesion Created Method and Size or Delivery Method | Fix and Cover | Immunosuppressive Treatment | |
| Wang S. N. et al., 2020 [16] | SD rats |
|
| Covered by piece of excised skull sealed with the bone wax | Cyclosporine A |
| Cao et al., 2023 [17] | SCID mice |
|
| ND | None |
| Cao et al., 2023 [31] | SCID mice |
|
| ND | None |
| Therapeutic—Parkinson’s Disease | |||||
| Animal | Model and Location | Lesion Created Method and Size or Delivery Method | Fix and Cover | Immunosuppressive Treatment | |
| Zheng et al., 2023 [32] | SCID mice |
|
| ND | None |
| Vascularization | |||||
| Animal | Location | Lesion Created Method and Size or Delivery Method | Fix and Cover | Immunosuppressive Treatment | |
| Pham, M. T. et al., 2018 [21] | SCID mice |
|
| ND | None |
| Shi, Y. et al., 2020 [22] | SCID mice |
|
| 3% low-melting agarose and adhesive glue to fix grafts | None |
| Disease modeling and cell study platform in vivo | |||||
| Animal | Location and Cavity size | Lesion Created Method and Size or Delivery Method | Fix and Cover | Immunosuppressive Treatment | |
| Huang et al. 2022 [33] | SCID mice |
|
| ND | None |
| Schafer et al., 2023 [34] | SCID mice |
|
| Covered by custom titanium head plate fixed with dental cement | None |
| Wang et al., 2024 [35] | SCID mice |
|
| Covered by 5 mm cover slip sealed with adhesive glue and dental cement for the wound | None |
| Chen et al., 2024 [36] | Athymic rats (newborn) |
|
| ND | None |
| Xu et al., 2024 [37] | SCID mice |
|
| ND | None |
| Li et al., 2024 [38] | SCID mice |
|
| Returned the hinged bone flap | None |
| Proof-of-Principle | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Cell Survival | Neuronal Differentiation | Axonal Projection | Electrophysiology | In vivo Microscopic Imaging | Optogenetics | Vascularization | Behavior Tests | Others | |
| Mansour et al., 2018 [12] | + | + | ND | + | + | + | + |
| |
| Daviaud et al., 2018 [13] | + | + | ND | ND | ND | ND | + | ND | |
| Kitahara et al., 2020 [25] | + | + | + | ND | ND | ND | + | ND |
|
| Dong et al., 2021 [26] | + | + | ND | + | ND | + | ND |
| |
| Revah et al., 2022 [19] | + | + | + | + | + | + | + |
|
|
| Wilson et al., 2022 [27] | + | + | ND | + | + | ND | + | ND |
|
| Jgamadze et al., 2023 [20] | + | + | + | + | ND | ND | + | ND |
|
| Li et al., 2024 [28] | + | + | + | + | + | - | + |
|
|
| Therapeutic—Traumatic Brain Injury | |||||||||
| Cell Survival | Neuronal Differentiation | Axonal Projection | Electrophysiology | In Vivo Microscopic Imaging | Optogenetics | Vascularization | Behavior Tests | Others | |
| Wang Z. et al., 2020 [14] | + | + | ND | ND | ND | ND | + |
|
|
| Bao et al., 2021 [15] | + | + | ND | + | ND | ND | + |
|
|
| Kim et al., 2022 [29] | + | + | ND | ND | ND | ND | + |
|
|
| Hu et al., 2024 [30] | + | + | + | + | ND | ND | + |
|
|
| Therapeutic—Stroke | |||||||||
| Cell Survival | Neuronal Differentiation | Axonal Projection | Electrophysiology | In Vivo Microscopic Imaging | Optogenetics | Vascularization | Behavior Tests | Others | |
| Wang S. N. et al., 2020 [16] | + | + | + | ND | ND | ND | + |
| |
| Cao et al., 2023 [17] | + | + | + | + | ND | + | ND |
| |
| Cao et al., 2023 [31] | + | + | ND | + | + | ND | ND |
| |
| Therapeutic—Parkinson’s Disease | |||||||||
| Cell Survival | Neuronal Differentiation | Axonal Projection | Electrophysiology | In vivo Microscopic Imaging | Optogenetics | Vascularization | Behavior Tests | Others | |
| Zheng et al., 2023 [32] | + | + | + | + | ND | ND | ND |
|
|
| Vascularization | |||||||||
| Cell Survival | Neuronal Differentiation | Axonal Projection | Electrophysiology | In Vivo Microscopic Imaging | Optogenetics | Vascularization | Behavior Tests | Others | |
| Pham, M. T. et al., 2018 [21] | + | ND | ND | ND | ND | ND | +, graft | ND | |
| Shi, Y. et al., 2020 [22] | + | + | ND | + | + | ND | +, graft | ND |
|
| Disease Modeling and Cell Study Platform In Vivo | |||||||||
| Cell Survival | Neuronal Differentiation | Axonal Projection | Electrophysiology | In Vivo Microscopic Imaging | Optogenetics | Vascularization | Behavior Tests | Others | |
| Huang et al. 2022 [33] | + | + | ND | ND | ND | ND | + |
| |
| Schafer et al., 2023 [34] | +, microglia | +, microglia | ND | ND | + | ND | ND | ND |
|
| Wang et al., 2024 [35] | +, astrocytes | +, astrocytes | ND | + | ND | ND | + | ND |
|
| Chen et al., 2024 [36] | + | ND | ND | ND | + | ND | ND | ND |
|
| Xu et al., 2024 [37] | + | + | ND | ND | + | ND | + | ND |
|
| Li et al., 2024 [38] | + | + | + | + | ND | ND | + |
|
|
| Conventional Culture | Brain Organoid-on-Chip | |
|---|---|---|
| Organoid | ||
| Necrotic core | Significant | Minimized |
| Neurogenesis and corticogenesis | Fair | Enhanced |
| Electrophysiological activity | Fair | Enhanced |
| Heterogeneity | High | Relatively lower |
| Batch variability | High | Relatively lower |
| System | ||
| Nutrients and gases exchange | Poor | Good |
| Precise microenvironments monitor and control | Hard | Feasible |
| Manual intervention/automatic level | Much/low | Less/high |
| Technical requirements | Relatively lower | High |
| System maintenance | Relatively easier | Complicated |
| Cost | Fair | Relatively higher in developing phase; may be lower in mature batch processing with precise control |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.-P.; Kokaia, Z. Brain Organoid Transplantation: A Comprehensive Guide to the Latest Advances and Practical Applications—A Systematic Review. Cells 2025, 14, 1074. https://doi.org/10.3390/cells14141074
Shen Y-P, Kokaia Z. Brain Organoid Transplantation: A Comprehensive Guide to the Latest Advances and Practical Applications—A Systematic Review. Cells. 2025; 14(14):1074. https://doi.org/10.3390/cells14141074
Chicago/Turabian StyleShen, Yu-Ping, and Zaal Kokaia. 2025. "Brain Organoid Transplantation: A Comprehensive Guide to the Latest Advances and Practical Applications—A Systematic Review" Cells 14, no. 14: 1074. https://doi.org/10.3390/cells14141074
APA StyleShen, Y.-P., & Kokaia, Z. (2025). Brain Organoid Transplantation: A Comprehensive Guide to the Latest Advances and Practical Applications—A Systematic Review. Cells, 14(14), 1074. https://doi.org/10.3390/cells14141074