
The Phosphodiesterase 4 Inhibitor Roflumilast Protects Microvascular Endothelial Cells from Irradiation-Induced Dysfunctions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Culture, Irradiation and Treatment of Microvascular Endothelial Cells
2.3. Adherens Junctions and Actin Cytoskeleton Stainings
2.4. Apoptosis Assay
2.5. Measurements of Endothelial Cell Surface Expression of Adhesion Molecules
2.6. Measurements of cAMP Concentration in Cell Lysate Samples
2.7. Measurements of Impedance in HPMEC Monolayers
2.8. Statistical Analysis
3. Results
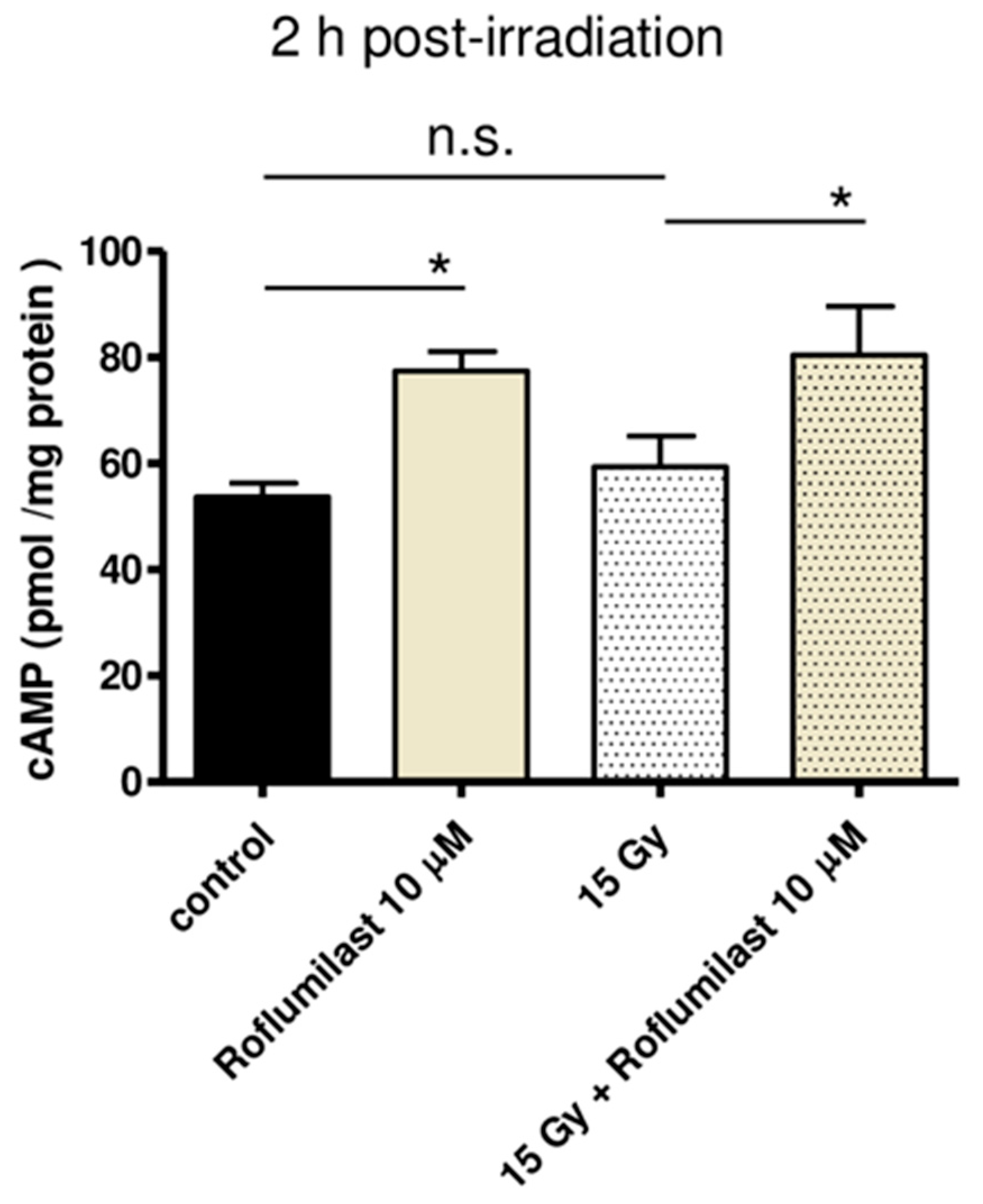
3.1. Effects of Roflumilast and Irradiation on Global cAMP Levels in HPMECs
3.2. Effects of Roflumilast and Irradiation on Impedance of HPMEC Monolayers
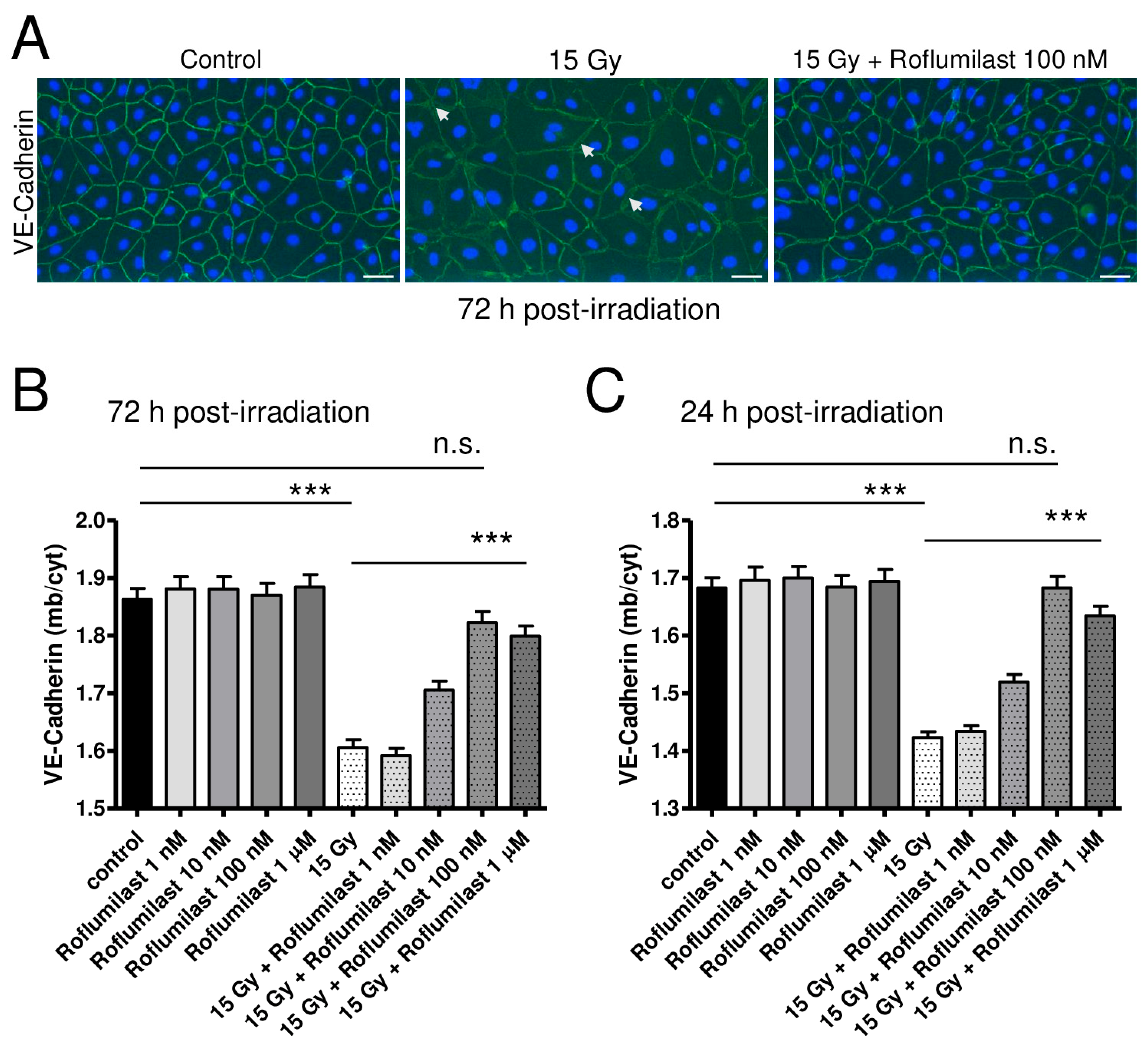
3.3. Effect of Roflumilast and Irradiation on Adherens Junction in HPMEC Monolayers
3.4. Effect of Roflumilast and Irradiation on the Actin Cytoskeleton in HPMEC Monolayers
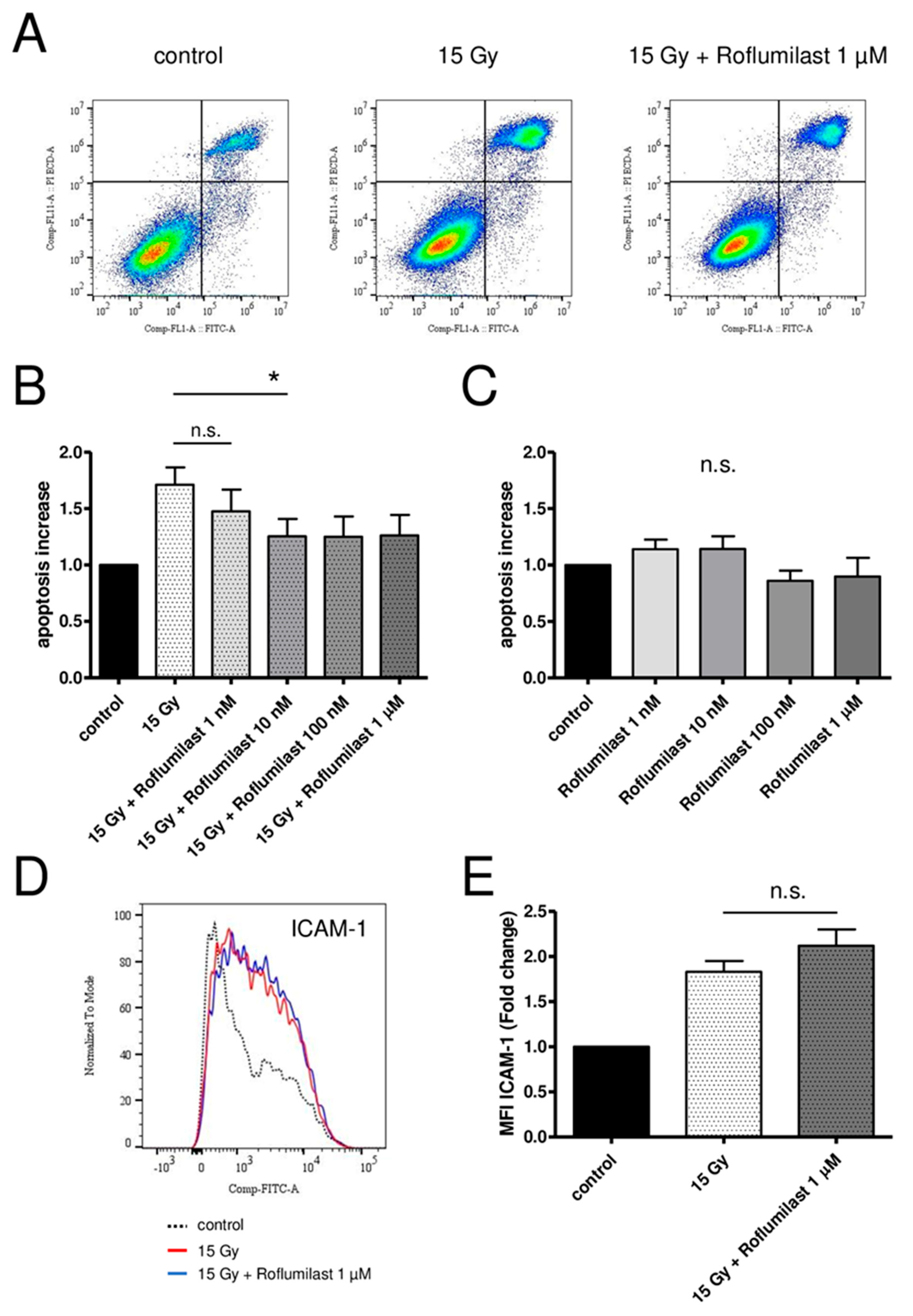
3.5. Effect of Roflumilast on Irradiation-Induced Apoptosis of HPMECs
3.6. Effect of Roflumilast on Irradiation-Induced ICAM-1 Overexpression in HPMECs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, J.P.; McBride, W.H. After the bomb drops: A new look at radiation-induced multiple organ dysfunction syndrome (MODS). nt. J. Radiat. Biol. 2011, 87, 851–868. [Google Scholar] [CrossRef] [PubMed]
- Baselet, B.; Sonveaux, P.; Baatout, S.; Aerts, A. Pathological effects of ionizing radiation: Endothelial activation and dysfunction. Cell. Mol. Life Sci. 2019, 76, 672–699. [Google Scholar] [CrossRef]
- Hallahan, D.E.; Virudachalam, S. Intercellular adhesion molecule 1 knockout abrogates radiation induced pulmonary inflammation. Proc. Natl. Acad. Sci. USA 1997, 94, 6432–6437. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, M.; Douwes, K.; Peter, R.; Degitz, K. Vascular activation of adhesion molecule mRNA and cell surface expression by ionizing radiation. Exp. Cell Res. 1998, 238, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Kouam, P.N.; Rezniczek, G.A.; Adamietz, I.A.; Buhler, H. Ionizing radiation increases the endothelial permeability and the transendothelial migration of tumor cells through ADAM10-activation and subsequent degradation of VE-cadherin. BMC Cancer 2019, 19, 958. [Google Scholar] [CrossRef]
- Paris, F.; Fuks, Z.; Kang, A.; Capodieci, P.; Juan, G.; Ehleiter, D.; Haimovitz-Friedman, A.; Cordon-Cardo, C.; Kolesnick, R. Endothelial apoptosis as the primary lesion initiating intestinal radiation damage in mice. Science 2001, 293, 293–297. [Google Scholar] [CrossRef]
- Quarmby, S.; Kumar, P.; Kumar, S. Radiation-induced normal tissue injury: Role of adhesion molecules in leukocyte-endothelial cell interactions. Int J. Cancer 1999, 82, 385–395. [Google Scholar] [CrossRef]
- Venkatesulu, B.P.; Mahadevan, L.S.; Aliru, M.L.; Yang, X.; Bodd, M.H.; Singh, P.K.; Yusuf, S.W.; Abe, J.I.; Krishnan, S. Radiation-Induced Endothelial Vascular Injury: A Review of Possible Mechanisms. JACC Basic Transl. Sci. 2018, 3, 563–572. [Google Scholar] [CrossRef]
- Wang, H.; Segaran, R.C.; Chan, L.Y.; Aladresi, A.A.M.; Chinnathambi, A.; Alharbi, S.A.; Sethi, G.; Tang, F.R. Gamma Radiation-Induced Disruption of Cellular Junctions in HUVECs Is Mediated through Affecting MAPK/NF-κB Inflammatory Pathways. Oxidative Med. Cell. Longev. 2019, 2019, 1486232. [Google Scholar] [CrossRef]
- Rho, S.S.; Ando, K.; Fukuhara, S. Dynamic Regulation of Vascular Permeability by Vascular Endothelial Cadherin-Mediated Endothelial Cell-Cell Junctions. J. Nippon. Med Sch. 2017, 84, 148–159. [Google Scholar] [CrossRef]
- Wakasugi, R.; Suzuki, K.; Kaneko-Kawano, T. Molecular Mechanisms Regulating Vascular Endothelial Permeability. Int. J. Mol. Sci. 2024, 25, 6415. [Google Scholar] [CrossRef] [PubMed]
- Radeva, M.Y.; Waschke, J. Mind the gap: Mechanisms regulating the endothelial barrier. Acta Physiol. 2018, 222, e12860. [Google Scholar] [CrossRef] [PubMed]
- Duong, C.N.; Vestweber, D. Mechanisms Ensuring Endothelial Junction Integrity Beyond VE-Cadherin. Front. Physiol. 2020, 11, 519. [Google Scholar] [CrossRef] [PubMed]
- van der Krogt, J.M.; van der Meulen, I.J.; van Buul, J.D. Spatiotemporal regulation of Rho GTPase signaling during endothelial barrier remodeling. Curr. Opin. Physiol. 2023, 34, 100676. [Google Scholar] [CrossRef]
- Kabacik, S.; Raj, K. Ionising radiation increases permeability of endothelium through ADAM10-mediated cleavage of VE-cadherin. Oncotarget 2017, 8, 82049–82063. [Google Scholar] [CrossRef]
- Narayanan, S.A.; Ford, J.; Zawieja, D.C. Impairment of lymphatic endothelial barrier function by X-ray irradiation. Int. J. Radiat. Biol. 2019, 95, 562–570. [Google Scholar] [CrossRef]
- Gabrys, D.; Greco, O.; Patel, G.; Prise, K.M.; Tozer, G.M.; Kanthou, C. Radiation effects on the cytoskeleton of endothelial cells and endothelial monolayer permeability. Int. J. Radiat. Oncol. Biol. Phys. 2007, 69, 1553–1562. [Google Scholar] [CrossRef]
- Guo, X.W.; Zhang, H.; Huang, J.Q.; Wang, S.N.; Lu, Y.; Cheng, B.; Dong, S.H.; Wang, Y.Y.; Li, F.S.; Li, Y.W. PIEZO1 Ion Channel Mediates Ionizing Radiation-Induced Pulmonary Endothelial Cell Ferroptosis via Ca2+/Calpain/VE-Cadherin Signaling. Front. Mol. Biosci. 2021, 8, 725274. [Google Scholar] [CrossRef]
- Na, K.; Cho, Y.; Choi, D.H.; Park, M.J.; Yang, J.H.; Chung, S. Gamma irradiation exposure for collapsed cell junctions and reduced angiogenesis of 3-D in vitro blood vessels. Sci. Rep. 2021, 11, 18230. [Google Scholar] [CrossRef]
- Boittin, F.X.; Guitard, N.; Toth, M.; Riccobono, D.; Thery, H.; Bobe, R. The Protein Kinase A Inhibitor KT5720 Prevents Endothelial Dysfunctions Induced by High-Dose Irradiation. Int. J. Mol. Sci. 2024, 25, 2269. [Google Scholar] [CrossRef]
- Sayner, S.L. Emerging themes of cAMP regulation of the pulmonary endothelial barrier. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 300, L667–L678. [Google Scholar] [CrossRef] [PubMed]
- Vielmuth, F.; Radeva, M.Y.; Yeruva, S.; Sigmund, A.M.; Waschke, J. cAMP: A master regulator of cadherin-mediated binding in endothelium, epithelium and myocardium. Acta Physiol. 2023, 238, e14006. [Google Scholar] [CrossRef]
- Spindler, V.; Peter, D.; Harms, G.S.; Asan, E.; Waschke, J. Ultrastructural analysis reveals cAMP-dependent enhancement of microvascular endothelial barrier functions via Rac1-mediated reorganization of intercellular junctions. Am. J. Pathol. 2011, 178, 2424–2436. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Hong, Z.; Huang, L.S.; Tsukasaki, Y.; Nepal, S.; Di, A.; Zhong, M.; Wu, W.; Ye, Z.; Gao, X.; et al. IL-1beta suppression of VE-cadherin transcription underlies sepsis-induced inflammatory lung injury. J. Clin. Invest. 2020, 130, 3684–3698. [Google Scholar] [CrossRef]
- Insel, P.A.; Zhang, L.; Murray, F.; Yokouchi, H.; Zambon, A.C. Cyclic AMP is both a pro-apoptotic and anti-apoptotic second messenger. Acta Physiol. 2012, 204, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kostin, S.; Flacke, J.P.; Reusch, H.P.; Ladilov, Y. Soluble adenylyl cyclase controls mitochondria-dependent apoptosis in coronary endothelial cells. J. Biol. Chem. 2009, 284, 14760–14768. [Google Scholar] [CrossRef]
- Sanz, M.J.; Cortijo, J.; Taha, M.A.; Cerda-Nicolas, M.; Schatton, E.; Burgbacher, B.; Klar, J.; Tenor, H.; Schudt, C.; Issekutz, A.C.; et al. Roflumilast inhibits leukocyte-endothelial cell interactions, expression of adhesion molecules and microvascular permeability. Br. J. Pharmacol. 2007, 152, 481–492. [Google Scholar] [CrossRef]
- Schick, M.A.; Wunder, C.; Wollborn, J.; Roewer, N.; Waschke, J.; Germer, C.T.; Schlegel, N. Phosphodiesterase-4 inhibition as a therapeutic approach to treat capillary leakage in systemic inflammation. J. Physiol. 2012, 590, 2693–2708. [Google Scholar] [CrossRef]
- Bogatcheva, N.V.; Zemskova, M.A.; Kovalenkov, Y.; Poirier, C.; Verin, A.D. Molecular mechanisms mediating protective effect of cAMP on lipopolysaccharide (LPS)-induced human lung microvascular endothelial cells (HLMVEC) hyperpermeability. J. Cell. Physiol. 2009, 221, 750–759. [Google Scholar] [CrossRef]
- Oishi, H.; Takano, K.; Tomita, K.; Takebe, M.; Yokoo, H.; Yamazaki, M.; Hattori, Y. Olprinone and colforsin daropate alleviate septic lung inflammation and apoptosis through CREB-independent activation of the Akt pathway. Am. J. Physiol. Cell. Mol. Physiol. 2012, 303, L130–L140. [Google Scholar] [CrossRef]
- Ma, H.; Shi, J.; Wang, C.; Guo, L.; Gong, Y.; Li, J.; Gong, Y.; Yun, F.; Zhao, H.; Li, E. Blockade of PDE4B limits lung vascular permeability and lung inflammation in LPS-induced acute lung injury. Biochem. Biophys. Res. Commun. 2014, 450, 1560–1567. [Google Scholar] [CrossRef]
- Cai, N.; Xu, B.; Li, X.; Qin, Y.; Li, M.; Chen, K.; Xu, J.; Wang, H. Roflumilast, a cyclic nucleotide phosphodiesterase 4 inhibitor, protects against cerebrovascular endothelial injury following cerebral ischemia/reperfusion by activating the Notch1/Hes1 pathway. Eur. J. Pharmacol. 2022, 926, 175027. [Google Scholar] [CrossRef]
- de Moraes-Souza, R.; Chahine, C.R.; Pera, C.I.; Mesquita, Y.; Sarto, R.; Lapenda, I.; Figueiredo, P.L.; Moury, L.; Herranz-Pinto, P. Efficacy and Safety of Topical Roflumilast for the Treatment of Psoriasis: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Clin. Drug Investig. 2024, 44, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Garbe, E.; Hoti, F.; Schink, T.; Svendsen, K.; Al-Eid, H.; Arkhammar, P.; Carlholm, M.; Fjallbrant, H.; Franzen, S.; Hedlund, C.; et al. Long-Term Safety of Roflumilast in Patients with Chronic Obstructive Pulmonary Disease, a Multinational Observational Database Cohort Study. Int. J. Chron. Obstruct. Pulmon. Dis. 2024, 19, 1879–1892. [Google Scholar] [CrossRef] [PubMed]
- Boittin, F.X.; Beddek, K.; Thery, H.; Pateux, J.; Cosler, G.; Riccobono, D.; Drouet, M.; Bobe, R. The immunosuppressant drug Cyclosporin A aggravates irradiation effects in endothelial cells. Biochem. Biophys. Res. Commun. 2022, 602, 127–134. [Google Scholar] [CrossRef] [PubMed]
- McRae, M.; LaFratta, L.M.; Nguyen, B.M.; Paris, J.J.; Hauser, K.F.; Conway, D.E. Characterization of cell-cell junction changes associated with the formation of a strong endothelial barrier. Tissue Barriers 2018, 6, e1405774. [Google Scholar] [CrossRef]
- Morandini, R.; Ghanem, G.; Portier-Lemarie, A.; Robaye, B.; Renaud, A.; Boeynaems, J.M. Action of cAMP on expression and release of adhesion molecules in human endothelial cells. Am. J. Physiol. Circ. Physiol. 1996, 270, H807–H816. [Google Scholar] [CrossRef]
- Blease, K.; Burke-Gaffney, A.; Hellewell, P.G. Modulation of cell adhesion molecule expression and function on human lung microvascular endothelial cells by inhibition of phosphodiesterases 3 and 4. Br. J. Pharmacol. 1998, 124, 229–237. [Google Scholar] [CrossRef]
- Rabe, K.F. Update on roflumilast, a phosphodiesterase 4 inhibitor for the treatment of chronic obstructive pulmonary disease. Br. J. Pharmacol. 2011, 163, 53–67. [Google Scholar] [CrossRef]
- Fan, T.; Wang, W.; Wang, Y.; Zeng, M.; Liu, Y.; Zhu, S.; Yang, L. PDE4 inhibitors: Potential protective effects in inflammation and vascular diseases. Front. Pharmacol. 2024, 15, 1407871. [Google Scholar] [CrossRef]
- Freeman, M.L. Gastrointestinal acute radiation syndrome: Current knowledge and perspectives. Cell Death Discov. 2025, 11, 235. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guitard, N.; Raffin, F.; Boittin, F.-X. The Phosphodiesterase 4 Inhibitor Roflumilast Protects Microvascular Endothelial Cells from Irradiation-Induced Dysfunctions. Cells 2025, 14, 1017. https://doi.org/10.3390/cells14131017
Guitard N, Raffin F, Boittin F-X. The Phosphodiesterase 4 Inhibitor Roflumilast Protects Microvascular Endothelial Cells from Irradiation-Induced Dysfunctions. Cells. 2025; 14(13):1017. https://doi.org/10.3390/cells14131017
Chicago/Turabian StyleGuitard, Nathalie, Florent Raffin, and François-Xavier Boittin. 2025. "The Phosphodiesterase 4 Inhibitor Roflumilast Protects Microvascular Endothelial Cells from Irradiation-Induced Dysfunctions" Cells 14, no. 13: 1017. https://doi.org/10.3390/cells14131017
APA StyleGuitard, N., Raffin, F., & Boittin, F.-X. (2025). The Phosphodiesterase 4 Inhibitor Roflumilast Protects Microvascular Endothelial Cells from Irradiation-Induced Dysfunctions. Cells, 14(13), 1017. https://doi.org/10.3390/cells14131017