
Regulation of Mitochondrial Metabolism by Mfn1 Gene Encoding Mitofusin Affects Cellular Proliferation and Histone Modification
 ,
,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Transfection
2.3. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Assay for Cell Proliferation Detection
2.4. 5-Ethynyl-2′-deoxyuridine (EdU) Detection of Cell Proliferation Ability
2.5. Annexin V-FITC Detection of Cell Apoptosis Ability
2.6. RNA Extraction and Quantitative RT-PCR
2.7. Mitochondrial Membrane Potential Assay
2.8. ATP Content Assay
2.9. Superoxide Dismutase (SOD) Activity Assay
2.10. Catalase (CAT) Activity Assay
2.11. Nicotinamide Adenine Dinucleotide Hydrogen (NADH/NAD+) Content Assay
2.12. Western Blot Analysis
2.13. Determination of Enzyme-Linked Immunosorbent Assay (ELISA) for α-KG, A-CoA, and SAM Content
2.14. RNA Sequencing Method
2.15. Metabolome Analysis
2.16. Statistical Analysis
3. Results
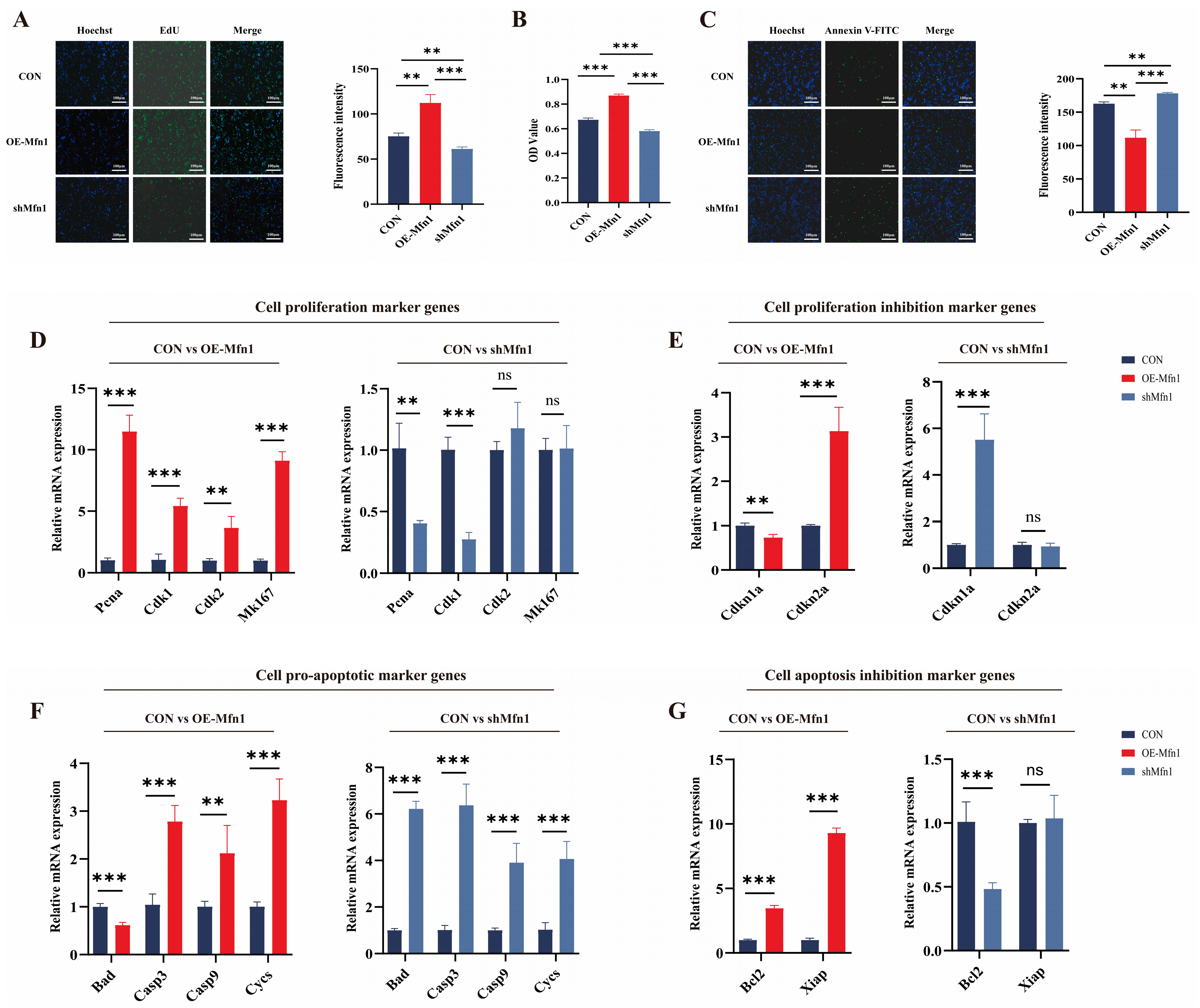
3.1. Changes in Mfn1 Gene Expression Affect Cell Proliferation and Apoptotic Capacity
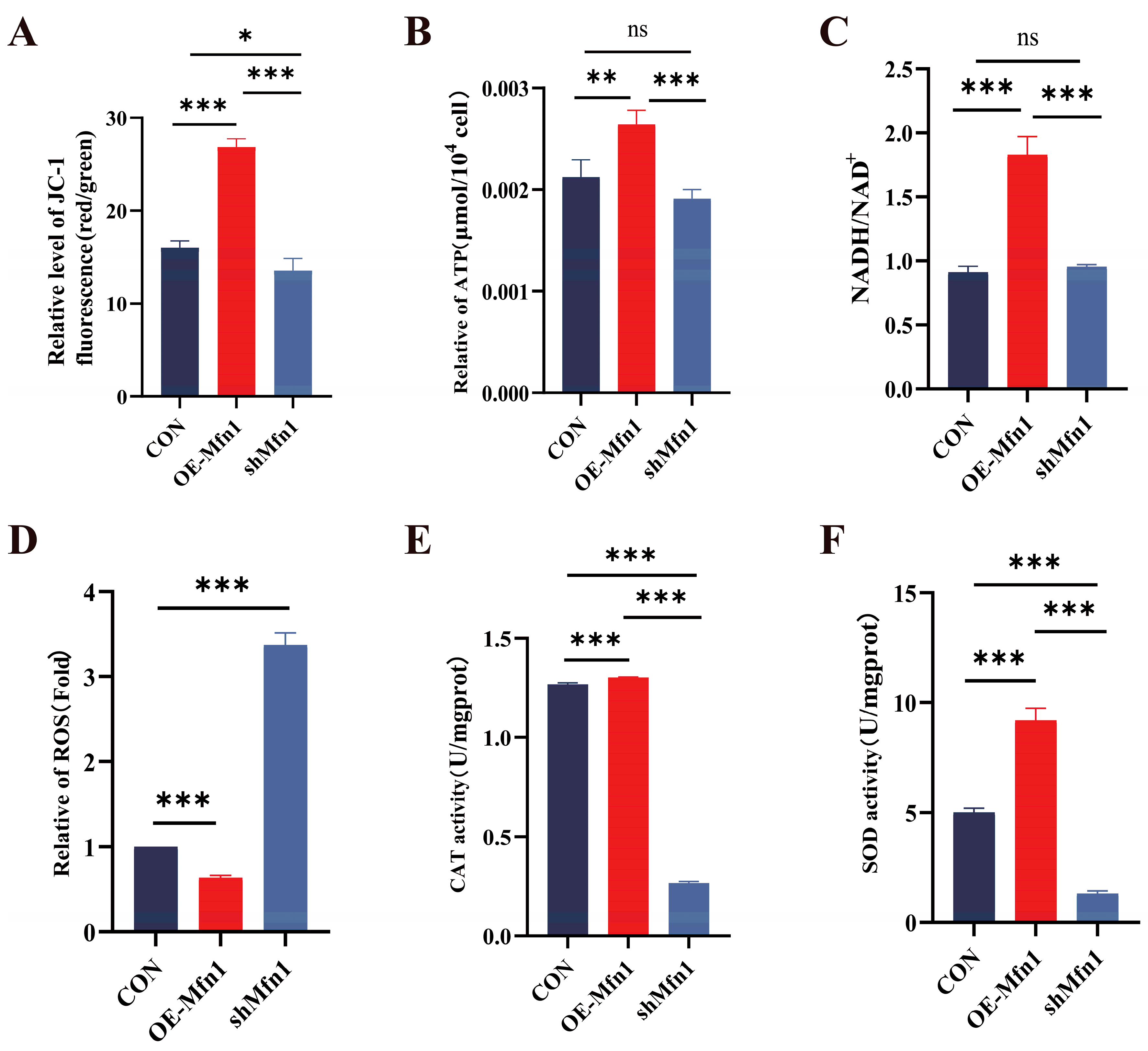
3.2. Mfn1 Gene Affects Mitochondrial Function
3.3. Overexpression and Downregulation of Mfn1 Gene Affects Expression of Many Genes
3.4. Metabolomic Analysis of OE-Mfn1 and shMfn1 Cells
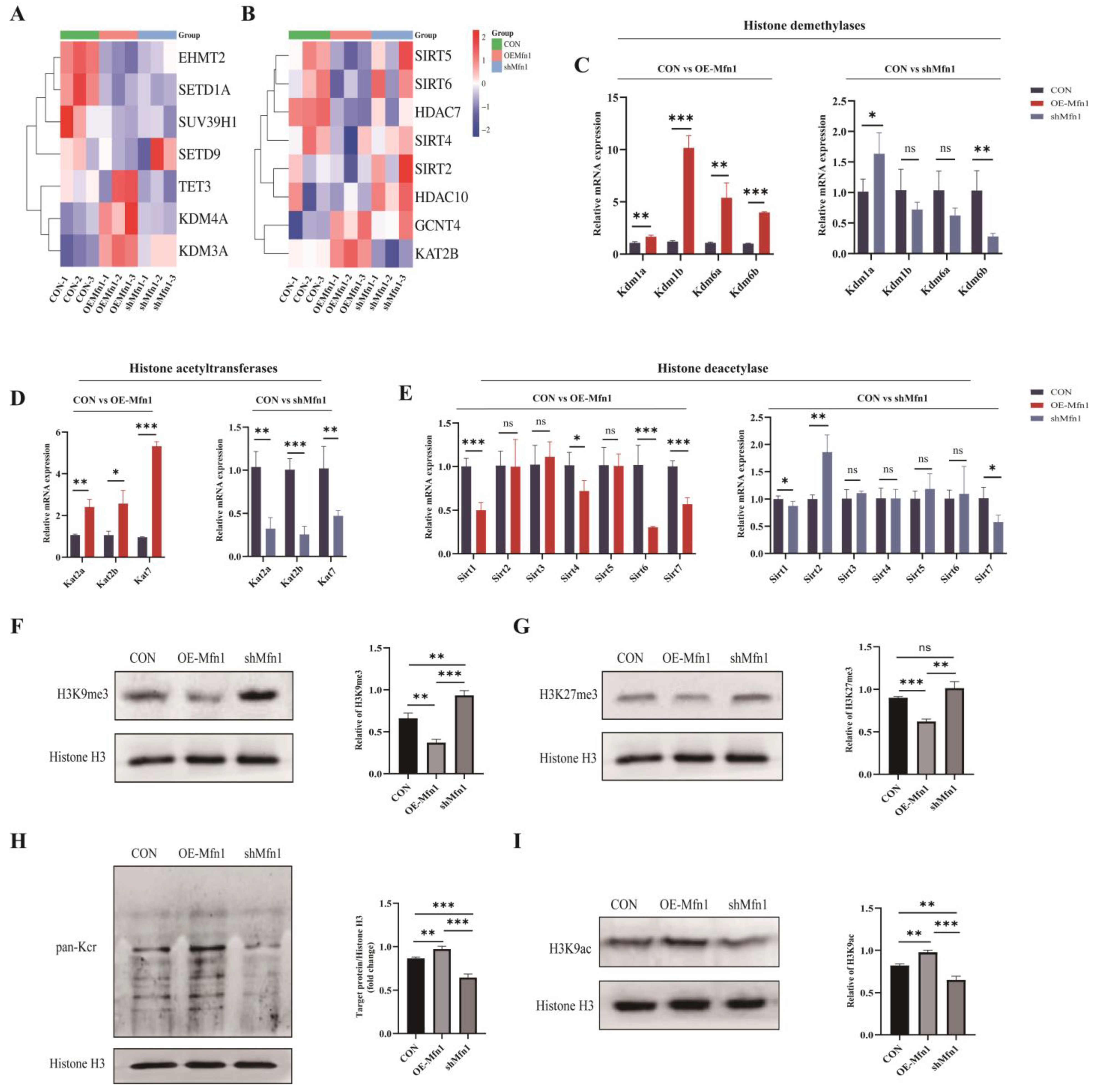
3.5. Mfn1 Gene Affects Protein Methylation and Acetylation
4. Discussion
5. Study Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Canovai, A.; Tribble, J.R.; Jöe, M.; Westerlund, D.Y.; Amato, R.; Trounce, I.A.; Monte, M.D.; Williams, P.A. Pyrroloquinoline quinone drives ATP synthesis in vitro and in vivo and provides retinal ganglion cell neuroprotection. Acta Neuropathol. Commun. 2023, 11, 146. [Google Scholar] [CrossRef]
- Ballard, A.; Zeng, R.; Zarei, A.; Shao, C.; Cox, L.; Yan, H.; Franco, A.; Dorn, G.W.; Faccio, R.; Veis, D.J. The tethering function of mitofusin2 controls osteoclast differentiation by modulating the Ca2+-NFATc1 axis. J. Biol. Chem. 2020, 295, 6629–6640. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Chen, P.; Xu, B.; Jin, Y.; Pan, Y.; Wu, T.; Du, Y.; Mao, J.; Wu, R. Abnormal expression of fission and fusion genes and the morphology of mitochondria in eutopic and ectopic endometrium. Eur. J. Med. Res. 2023, 28, 209. [Google Scholar] [CrossRef]
- Baek, A.; Son, S.; Baek, Y.M.; Kim, D.E. KRT8 (keratin 8) attenuates necrotic cell death by facilitating mitochondrial fission-mediated mitophagy through interaction with PLEC (plectin). Autophagy 2021, 17, 3939–3956. [Google Scholar] [CrossRef]
- Shannar, A.; Sarwar, M.S.; Kong, A.T. A new frontier in studying dietary phytochemicals in cancer and in health: Metabolic and epigenetic reprogramming. Prev. Nutr. Food. Sci. 2022, 27, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Yeung, B.H.Y.; Huang, J.; An, S.S.; Solway, J.; Mitzner, W.; Tang, W.Y. Role of isocitrate dehydrogenase 2 on DNA hydroxymethylation in human airway smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 2020, 63, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Singh, K.; Shing, V.; Gupta, A.K.; Huffstutler, R.D.; Lee, D.Y.; Sack, M.N. The mitochondrial thiolase ACAT1 regulates monocyte/macrophage type I interferon via epigenetic control. BioRxiv 2024. BioRxiv:01.29.577773. [Google Scholar] [CrossRef]
- Jurich, C.; Yang, Z.J. High-throughput computational investigation of protein electrostatics and cavity for SAM-dependent methyltransferases. Protein. Sci. 2023, 32, e4690. [Google Scholar] [CrossRef]
- Liu, X.; Si, W.; He, L.; Yang, J.; Peng, Y.; Ren, J.; Liu, X.; Jin, T.; Yu, H.; Zhang, Z.; et al. The existence of a nonclassical TCA cycle in the nucleus that wires the metabolic-epigenetic circuitry. Signal. Transduct. Target. Ther. 2021, 6, 375. [Google Scholar] [CrossRef]
- Mohammed, S.A.; Ambrosini, S.; Lüscher, T.; Paneni, F.; Costantino, S. Epigenetic control of mitochondrial function in the vasculature. Front. Cardiovasc. Med. 2020, 7, 28. [Google Scholar] [CrossRef]
- Zhou, Z.; Ibekwe, E.; Chornenkyy, Y. Metabolic alterations in cancer cells and the emerging role of oncometabolites as drivers of neoplastic change. Antioxidants 2018, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Jasra, I.T.; Cuesta-Gomez, N.; Verhoeff, K.; Marfil-Garza, B.A.; Dadheech, N.; Shapiro, A.M.J. Mitochondrial regulation in human pluripotent stem cells during reprogramming and β cell differentiation. Front. Endocrinol. 2023, 14, 1236472. [Google Scholar] [CrossRef]
- Wu, S.; Zhao, X.; Wu, M.; Yang, L.; Liu, X.; Li, D.; Xu, H.; Zhao, Y.; Su, X.; Wei, Z.; et al. Low expression of mitofusin 1 gene leads to mitochondrial dysfunction and embryonic genome activation failure in ovine-bovine inter-species cloned embryos. Int. J. Mol. Sci. 2022, 23, 10145. [Google Scholar] [CrossRef]
- Kunlayawutipong, T.; Apaijai, N.; Tepmalai, K.; Kongkarnka, S.; Leerapun, A.; Pinyopornpanish, K.; Soontornpun, A.; Chattipakorn, S.C.; Chattipakorn, N.; Pinyopornpanish, K. Imbalance of mitochondrial fusion in peripheral blood mononuclear cells is associated with liver fibrosis in patients with metabolic dysfunction-associated steatohepatitis. Heliyon 2024, 10, e27557. [Google Scholar] [CrossRef]
- Koshiba, T.; Detmer, S.A.; Kaiser, J.T.; Chen, H.; McCaffery, J.M.; Chan, D.C. Structural basis of mitochondrial tethering by mitofusin complexes. Science 2004, 305, 858–862. [Google Scholar] [CrossRef] [PubMed]
- Chin, R.M.; Fu, X.; Pai, M.Y.; Vergnes, L.; Hwang, H.; Deng, G.; Diep, S.; Lomenick, B.; Meli, V.S.; Monsalve, G.C.; et al. The metabolite α-ketoglutarate extends lifespan by inhibiting ATP synthase and TOR. Nature 2014, 510, 397–401. [Google Scholar] [CrossRef]
- Mentch, S.J.; Mehrmohamadi, M.; Huang, L.; Liu, X.; Gupta, D.; Mattocks, D.; Padilla, P.G.; Ables, G.; Bamman, M.M.; Thalacker-Mercer, A.E.; et al. Histone Methylation Dynamics and Gene Regulation Occur through the Sensing of One-Carbon Metabolism. Cell Metab. 2015, 22, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Shyh-Chang, N.; Locasale, J.W.; Lyssiotis, C.A.; Zheng, Y.; Teo, R.Y.; Ratanasirintrawoot, S.; Zhang, J.; Onder, T.; Unternaehrer, J.J.; Zhu, H.; et al. Influence of threonine metabolism on S-adenosylmethionine and histone methylation. Science 2013, 339, 222–226. [Google Scholar] [CrossRef]
- Sutendra, G.; Kinnaird, A.; Dromparis, P.; Paulin, R.; Stenson, T.H.; Haromy, A.; Hashimoto, K.; Zhang, N.; Flaim, E.; Michelakis, E.D. A nuclear pyruvate dehydrogenase complex is important for the generation of acetyl-CoA and histone acetylation. Cell 2014, 158, 84–97. [Google Scholar] [CrossRef]
- Martinez, S.; Hausinger, R.P. Catalytic mechanisms of fe(II)- and 2-oxoglutarate-dependent oxygenases. J. Biol. Chem. 2015, 290, 20702–20711. [Google Scholar] [CrossRef]
- Papanicolaou, K.N.; Ngoh, G.A.; Dabkowski, E.R.; O’Connell, K.A.; Ribeiro, R.; Stanley, W.C.; Walsh, K. Cardiomyocyte deletion of mitofusin-1 leads to mitochondrial fragmentation and improves tolerance to ROS-induced mitochondrial dysfunction and cell death. Am. J. Physiol. Heart. Circ. Physiol. 2012, 302, H167–H179. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.C.; Madeira, N.; Morais, S.; Macedo, A.; Cruz, M.T.; Pereira, C.M.F. Mitochondria fusion upon SERCA inhibition prevents activation of the NLRP3 inflammasome in human monocytes. Cells 2022, 11, 433. [Google Scholar] [CrossRef]
- Yan, L.; Qi, Y.; Huang, X.; Yu, C.; Lan, L.; Guo, X.; Rao, Z.; Hu, J.; Lou, Z. Structural basis for GTP hydrolysis and conformational change of MFN1 in mediating membrane fusion. Nat. Struct. Mol. Biol. 2018, 25, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, L.; Qi, Y.; Xu, H. Mitochondrial cAMP signaling. Cell. Mol. Life. Sci. 2016, 73, 4577–4590. [Google Scholar] [CrossRef]
- Fontana, F.; Giannitti, G.; Marchesi, S.; Limonta, P. The PI3K/Akt pathway and glucose metabolism: A dangerous liaison in cancer. Int. J. Biol. Sci. 2024, 20, 3113–3125. [Google Scholar] [CrossRef] [PubMed]
- Bahar, M.E.; Kim, H.J.; Kim, D.R. Targeting the RAS/RAF/MAPK pathway for cancer therapy: From mechanism to clinical studies. Signal. Transduct. Target. Ther. 2023, 8, 455. [Google Scholar] [CrossRef]
- Miao, J.; Liu, J.; Niu, J.; Zhang, Y.; Shen, W.; Luo, C.; Liu, Y.; Li, C.; Li, H.; Yang, P.; et al. Wnt/β-catenin/RAS signaling mediates age-related renal fibrosis and is associated with mitochondrial dysfunction. Aging. Cell 2019, 18, e13004. [Google Scholar] [CrossRef]
- Garza, S.; Chen, L.; Galano, M.; Cheung, G.; Sottas, C.; Li, L.; Li, Y.; Zirkin, B.R. Mitochondrial dynamics, Leydig cell function, and age-related testosterone deficiency. FASEB J. 2022, 36, e22637. [Google Scholar] [CrossRef]
- Yamada, S.; Kubo, Y.; Yamazaki, D.; Sekino, Y.; Kanda, Y. Chlorpyrifos inhibits neural induction via Mfn1-mediated mitochondrial dysfunction in human induced pluripotent stem cells. Sci. Rep. 2017, 7, 40925. [Google Scholar] [CrossRef]
- Xiong, W.; Ma, Z.; An, D.; Liu, Z.; Cai, W.; Bai, Y.; Zhan, Q.; Lai, W.; Zeng, Q.; Ren, H.; et al. Mitofusin 2 participates in mitophagy and mitochondrial fusion against angiotensin II-induced cardiomyocyte injury. Front. Physiol. 2019, 10, 411. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, H. Inhibitory effect of astaxanthin on oxidative stress-induced mitochondrial dysfunction-A mini-review. Nutrients 2018, 10, 1137. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.J.; Nie, Z.W.; Shin, K.T.; Zhou, W.; Cui, X.S. PINK1 regulates mitochondrial morphology via promoting mitochondrial fission in porcine preimplantation embryos. FASEB J. 2019, 33, 7882–7895. [Google Scholar] [CrossRef]
- Depaoli, M.R.; Karsten, F.; Madreiter-Sokolowski, C.T.; Klec, C.; Gottschalk, B.; Bischof, H.; Eroglu, E.; Waldeck-Weiermair, M.; Simmen, T.; Graier, W.F.; et al. Real-time imaging of mitochondrial ATP dynamics reveals the metabolic setting of single cells. Cell Rep. 2018, 25, 501–512.e3. [Google Scholar] [CrossRef]
- Jing, M.; Zhang, H.; Wei, M.; Tang, Y.; Xia, Y.; Chen, Y.; Shen, Z.; Chen, C. Reactive oxygen species partly mediate DNA Mmethylation in responses to different heavy metals in pokeweed. Front. Plant. Sci. 2022, 13, 845108. [Google Scholar] [CrossRef]
- Villanueva, M.T. Tumour metabolism: The sugar-free, full-fat diet. Nat. Rev. Cancer 2016, 16, 615. [Google Scholar] [CrossRef]
- Chen, S.; Whetstine, J.R.; Ghosh, S.; Hanover, J.A.; Gali, R.R.; Grosu, P.; Shi, Y. The conserved NAD(H)-dependent corepressor CTBP-1 regulates Caenorhabditis elegans life span. Proc. Natl. Acad. Sci. USA 2009, 106, 1496–1501. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Hwang, I.S.; Kwak, T.U.; Lim, J.H.; Hwang, S.; Cho, S.K. Low expression of mitofusin 1 is associated with mitochondrial dysfunction and apoptosis in porcine somatic cell nuclear transfer embryos. Anim. Sci. J. 2020, 91, e13430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Bener, M.B.; Jiang, Z.; Wang, T.; Esencan, E.; Iii, R.S.; Horvath, T.; Seli, E. Mitofusin 1 is required for female fertility and to maintain ovarian follicular reserve. Cell Death. Dis. 2019, 10, 560. [Google Scholar] [CrossRef]
- Londoño Gentile, T.; Lu, C.; Lodato, P.M.; Tse, S.; Olejiniczak, S.H.; Witze, E.S.; Thompson, C.B.; Wellen, K.E. DNMT1 is regulated by ATP-citrate lyase and maintains methylation patterns during adipocyte differentiation. Mol. Cell. Biol. 2013, 33, 3864–3878. [Google Scholar] [CrossRef]
- Phoyen, S.; Sanpavat, A.; Ma-On, C.; Stein, U.; Hirankarn, N.; Tangkijvanich, P.; Jindatip, D.; Whongsiri, P.; Boonla, C. H4K20me3 upregulated by reactive oxygen species is associated with tumor progression and poor prognosis in patients with hepatocellular carcinoma. Heliyon 2023, 9, e22589. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, L.; Lai, C.; Peng, X.; Yu, S.; Zhou, C.; Zhang, B.; Zhang, W. Dynamic alteration profile and new role of RNA m6A methylation in replicative and H2O2-induced premature senescence of human embryonic lung fibroblasts. Int. J. Mol. Sci. 2022, 23, 9271. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lee, J.-E.; Cho, Y.-W.; Xiao, Y.; Jin, Q.; Liu, C.; Ge, K. UTX regulates mesoderm differentiation of embryonic stem cells independent of H3K27 demethylase activity. Proc. Natl. Acad. Sci. USA 2012, 109, 15324–15329. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liang, W.; Yang, Z.; Chen, Z.; Du, Z.; Gong, S. SIRT3-mediated deacetylation protects inner hair cell synapses in a H2O2-induced oxidative stress model in vitro. Exp. Cell Res. 2022, 418, 113280. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Scott, I.; Webster, B.R.; Sack, M.N. The emerging characterization of lysine residue deacetylation on the modulation of mitochondrial function and cardiovascular biology. Circ. Res. 2009, 105, 830–841. [Google Scholar] [CrossRef]
- Moussaieff, A.; Rouleau, M.; Kitsberg, D.; Cohen, M.; Levy, G.; Barasch, D.; Nemirovski, A.; Shen-Orr, S.; Laevsky, I.; Amit, M.; et al. Glycolysis-mediated changes in acetyl-CoA and histone acetylation control the early differentiation of embryonic stem cells. Cell Metab. 2015, 21, 392–402. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Zhao, X.; Yun, Y.; Gao, Y.; Bo, C.; Song, L.; Bai, C.; Yang, L.; Li, G.; Su, G. Regulation of Mitochondrial Metabolism by Mfn1 Gene Encoding Mitofusin Affects Cellular Proliferation and Histone Modification. Cells 2025, 14, 1015. https://doi.org/10.3390/cells14131015
Xu H, Zhao X, Yun Y, Gao Y, Bo C, Song L, Bai C, Yang L, Li G, Su G. Regulation of Mitochondrial Metabolism by Mfn1 Gene Encoding Mitofusin Affects Cellular Proliferation and Histone Modification. Cells. 2025; 14(13):1015. https://doi.org/10.3390/cells14131015
Chicago/Turabian StyleXu, Han, Xiaoyu Zhao, Yuan Yun, Yuxin Gao, Chunjie Bo, Lishuang Song, Chunling Bai, Lei Yang, Guangpeng Li, and Guanghua Su. 2025. "Regulation of Mitochondrial Metabolism by Mfn1 Gene Encoding Mitofusin Affects Cellular Proliferation and Histone Modification" Cells 14, no. 13: 1015. https://doi.org/10.3390/cells14131015
APA StyleXu, H., Zhao, X., Yun, Y., Gao, Y., Bo, C., Song, L., Bai, C., Yang, L., Li, G., & Su, G. (2025). Regulation of Mitochondrial Metabolism by Mfn1 Gene Encoding Mitofusin Affects Cellular Proliferation and Histone Modification. Cells, 14(13), 1015. https://doi.org/10.3390/cells14131015