
Lovastatin Targets the USP14–Survivin Axis to Suppress Triple-Negative Breast Cancer via Ubiquitin-Mediated Proteasomal Degradation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Reagents and Antibodies
2.3. Plasmid Constructs and Lentiviral Infection
2.4. Cell Viability Analysis
2.5. Soft Agar Assay
2.6. Flow Cytometry Analysis
2.7. Western Blot Assay
2.8. Co-Immunoprecipitation
2.9. Subcellular Fraction Isolation
2.10. Immunofluorescence
2.11. Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
2.12. Caspase 3 Activity Assay
2.13. Ubiquitination Analysis
2.14. Cycloheximide Pulse-Chase Assay
2.15. Xenograft Tumor Model
2.16. Immunohistochemistry
2.17. Bioinformatics Analysis
2.18. Statistical Analysis
3. Results
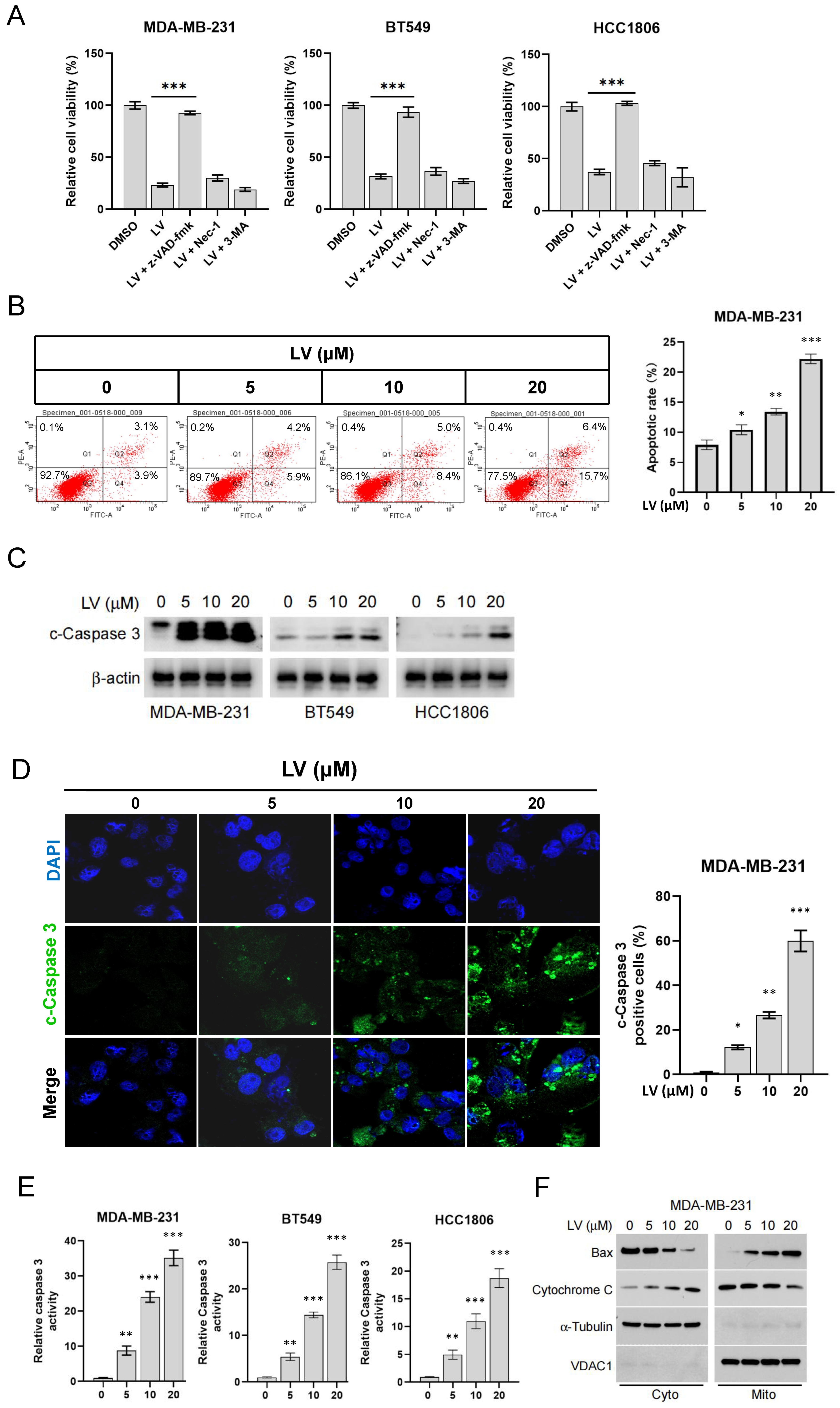
3.1. LV Induces Apoptosis in TNBC Cells Through Activation of the Intrinsic Apoptotic Pathway
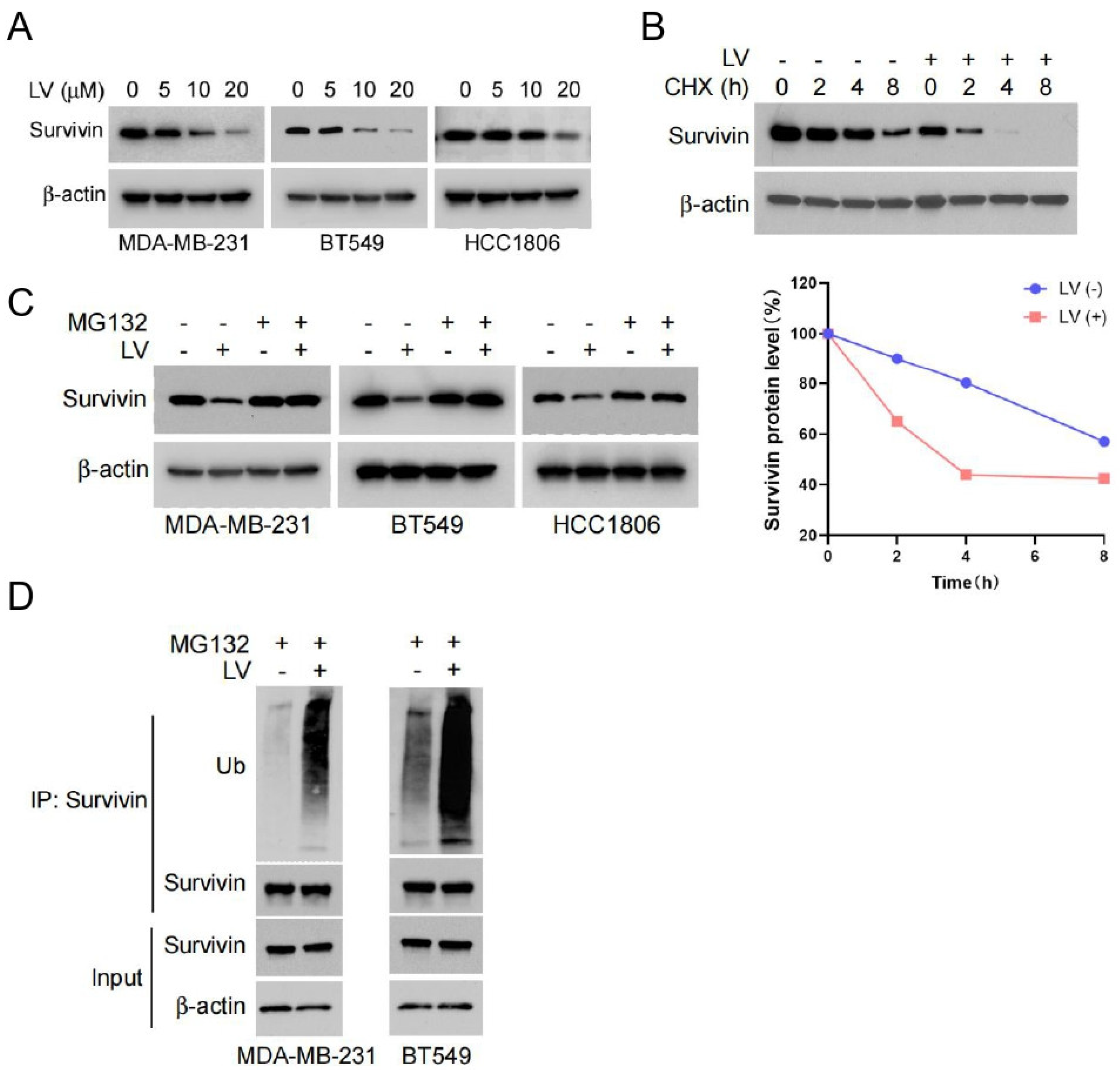
3.2. LV Induces the Degradation of Survivin via the Ubiquitin–Proteasome Pathway in TNBC Cells
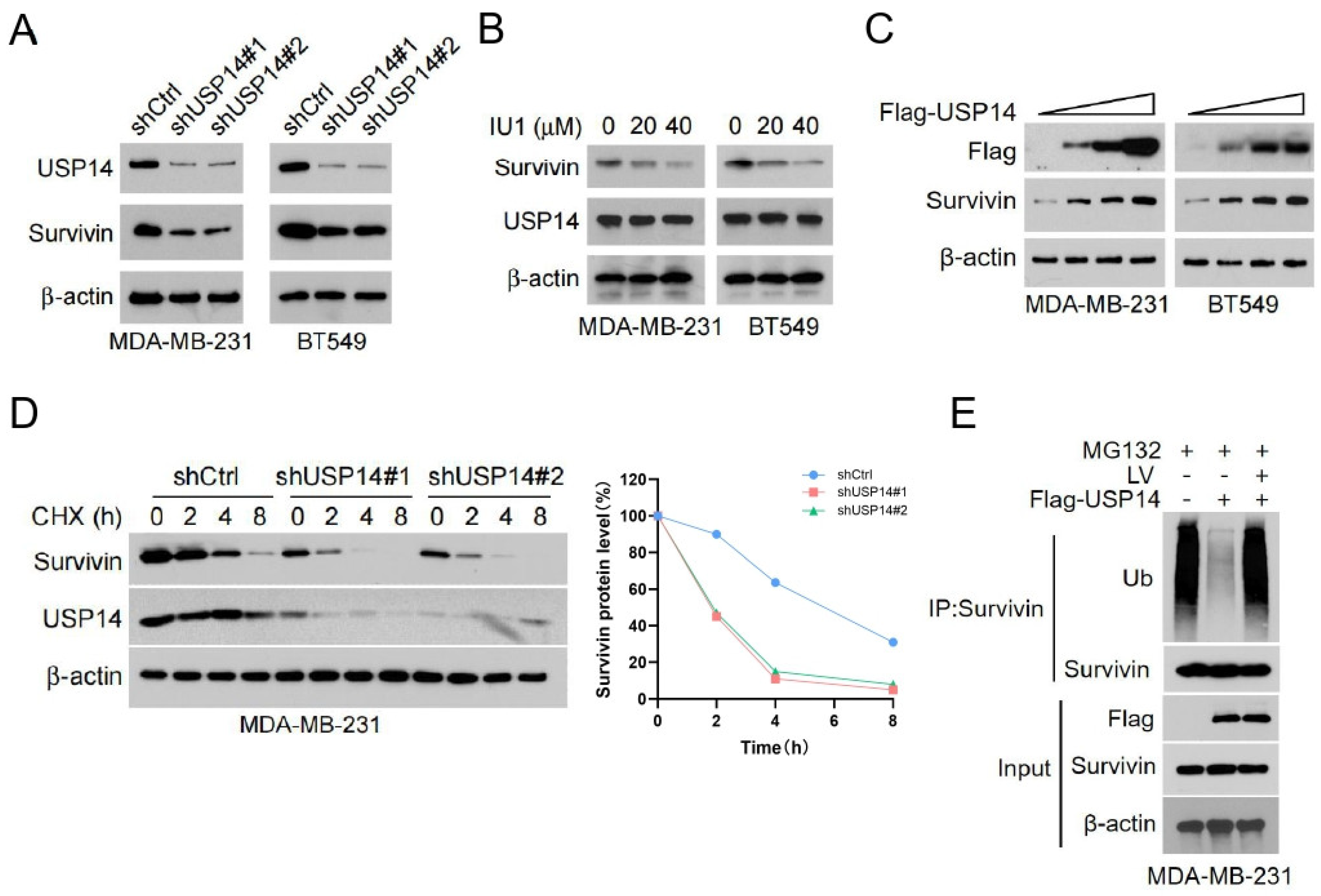
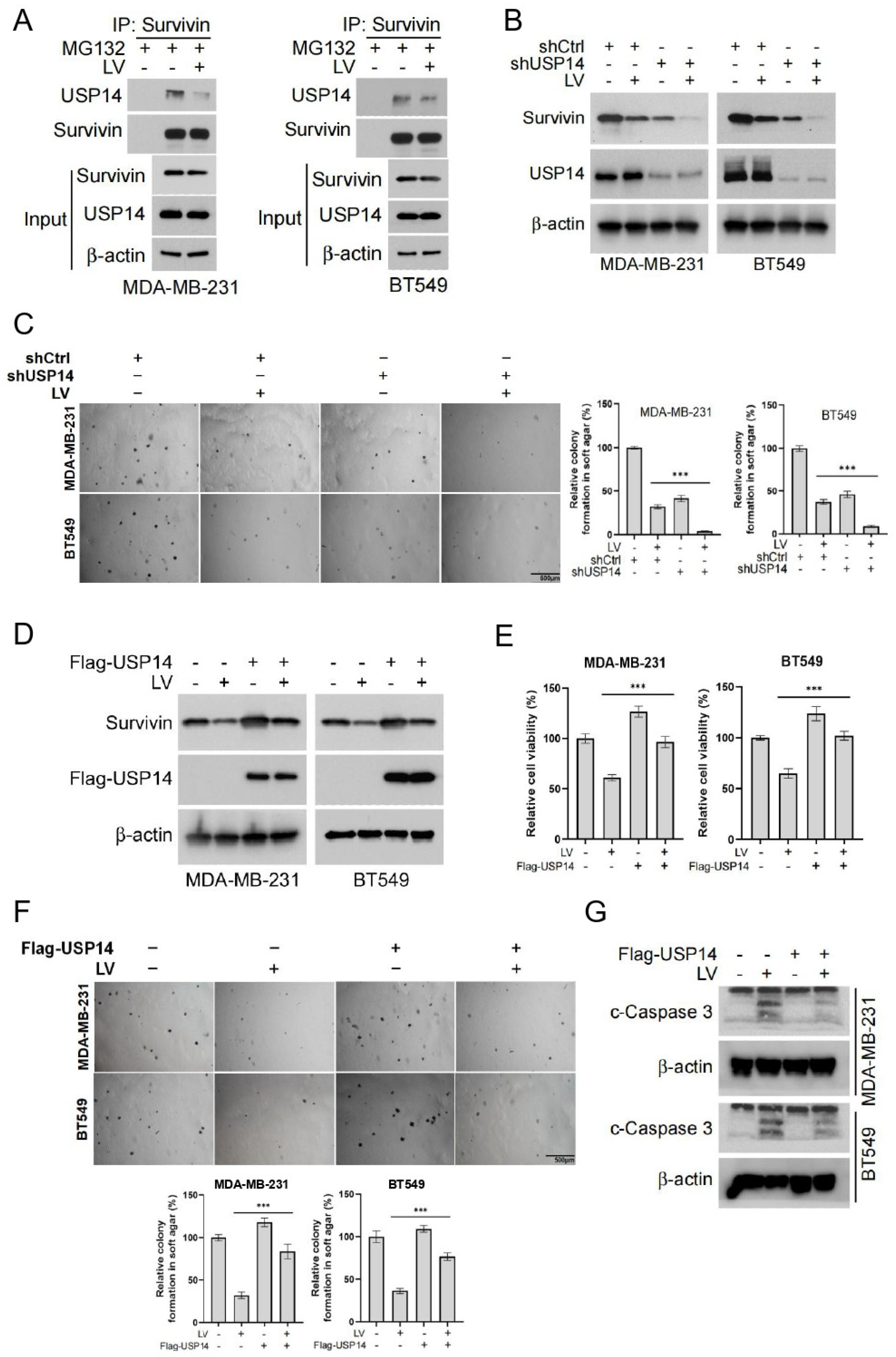
3.3. LV Destabilizes Survivin by Disrupting the Interaction Between USP14 and Survivin in TNBC Cells
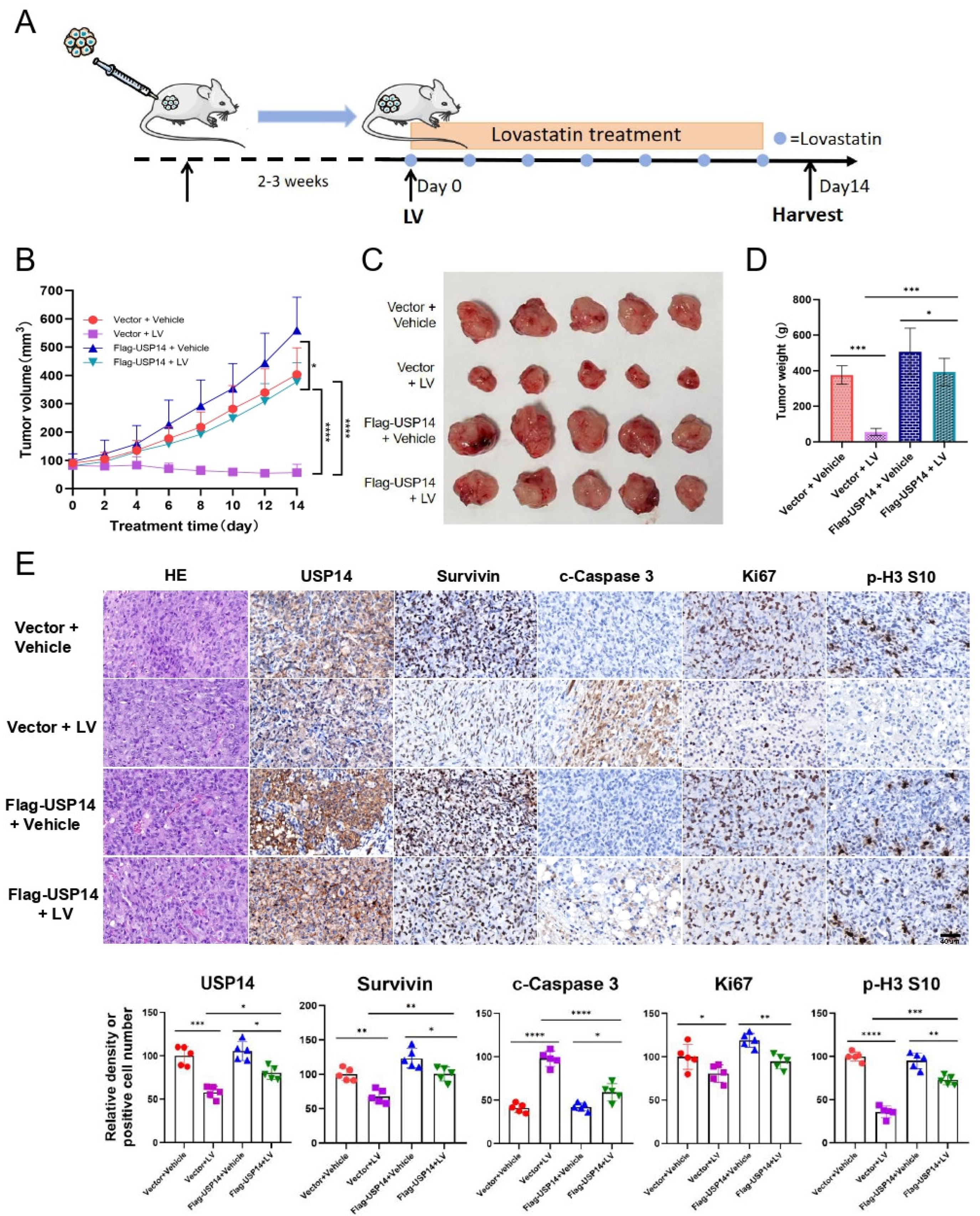
3.4. The Overexpression of USP14 Abrogates the Anti-Tumor Activity of LV In Vivo
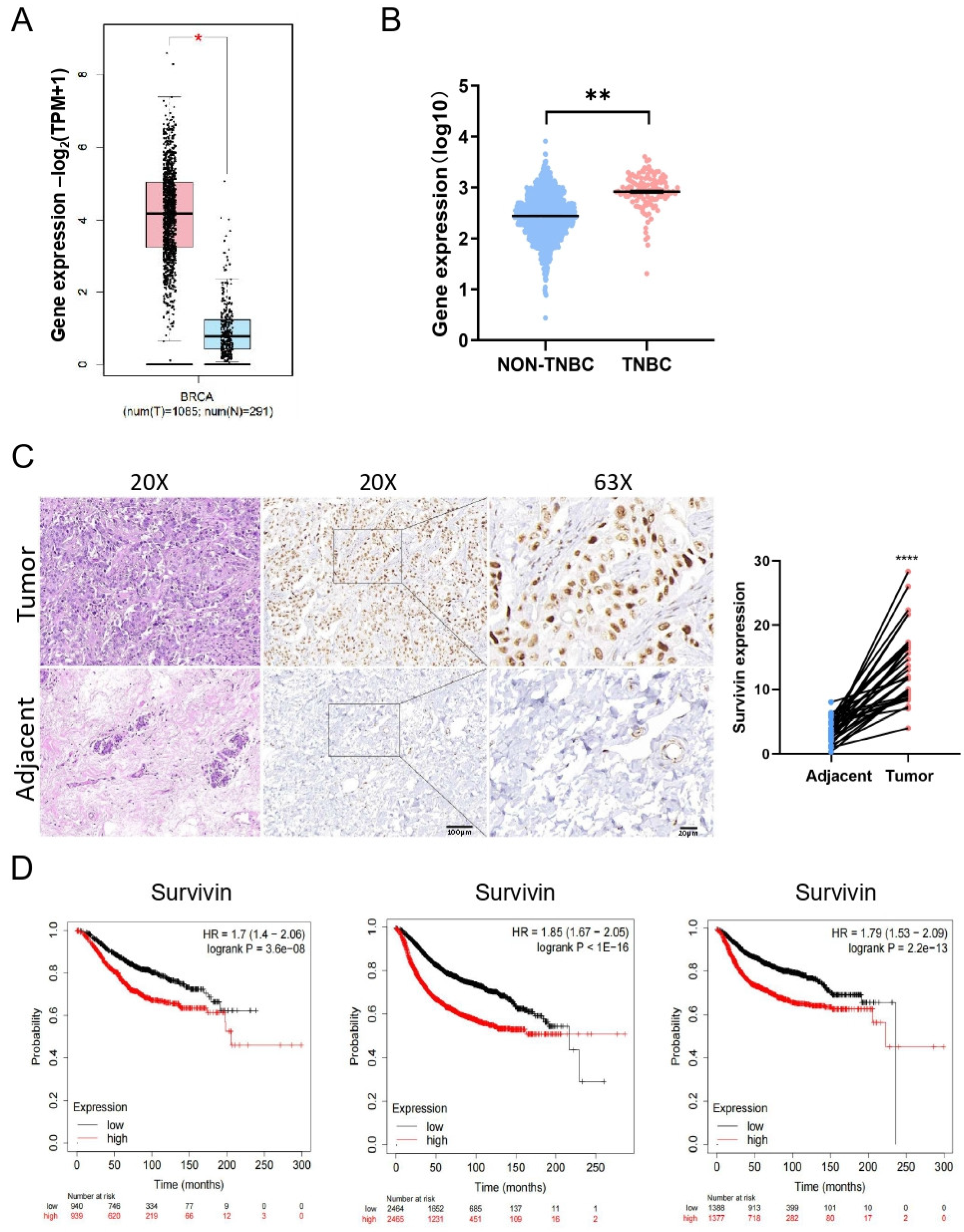
3.5. Higher Levels of Survivin Predict Poorer Prognosis in TNBC Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, U.; Boyle, P.; Goldhirsch, A.; Orecchia, R.; Viale, G. Breast cancer. Lancet 2005, 365, 1727–1741. [Google Scholar] [CrossRef]
- Liedtke, C.; Mazouni, C.; Hess, K.R.; André, F.; Tordai, A.; Mejia, J.A.; Symmans, W.F.; Gonzalez-Angulo, A.M.; Hennessy, B.; Green, M.; et al. Response to neoadjuvant therapy and long-term survival in patients with triple-negative breast cancer. J. Clin. Oncol. 2008, 26, 1275–1281. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Merkher, Y.; Chen, L.; Liu, N.; Leonov, S.; Chen, Y. Recent advances in therapeutic strategies for triple-negative breast cancer. J. Hematol. Oncol. 2022, 15, 121–150. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xu, P.; Wang, J.; Ji, H.; Zhang, L.; Tang, Z. Advancements in clinical research and emerging therapies for triple-negative breast cancer treatment. Eur. J. Pharmacol. 2024, 988, 177202. [Google Scholar] [CrossRef]
- Jia, H.; Truica, C.I.; Wang, B.; Wang, Y.; Ren, X.; Harvey, H.A.; Song, J.; Yang, J.-M. Immunotherapy for triple-negative breast cancer: Existing challenges and exciting prospects. Drug Resist. Updat. 2017, 32, 1–15. [Google Scholar] [CrossRef]
- Bianchini, G.; De Angelis, C.; Licata, L.; Gianni, L. Treatment landscape of triple-negative breast cancer—Expanded options, evolving needs. Nat. Rev. Clin. Oncol. 2022, 19, 91–113. [Google Scholar] [CrossRef] [PubMed]
- Mita, A.C.; Mita, M.M.; Nawrocki, S.T.; Giles, F.J. Survivin: Key Regulator of mitosis and apoptosis and novel target for cancer therapeutics. Clin. Cancer Res. 2008, 14, 5000–5005. [Google Scholar] [CrossRef]
- Frazzi, R. BIRC3 and BIRC5: Multi-faceted inhibitors in cancer. Cell Biosci. 2021, 11, 8–21. [Google Scholar] [CrossRef]
- Garlapati, C.; Joshi, S.; Bhattarai, S.; Krishnamurthy, J.; Turaga, R.C.; Nguyen, T.; Li, X.; Aneja, R. PLK1 and AURKB phosphorylate survivin differentially to affect proliferation in racially distinct triple-negative breast cancer. Cell Death Dis. 2023, 14, 12. [Google Scholar] [CrossRef]
- Jia, Y.; Hu, R.; Li, P.; Zheng, Y.; Wang, Y.; Ma, X. DEC1 is required for anti-apoptotic activity of gastric cancer cells under hypoxia by promoting Survivin expression. Gastric Cancer 2018, 21, 632–642. [Google Scholar] [CrossRef]
- Li, X.; Liang, Q.; Zhou, L.; Deng, G.; Xiao, Y.; Gan, Y.; Han, S.; Liao, J.; Wang, R.; Qing, X.; et al. Survivin degradation by bergenin overcomes pemetrexed resistance. Cell. Oncol. 2023, 46, 1837–1853. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wang, Z.; Zhang, H.; Wang, Y.; Li, D.-Q. Survivin-positive circulating tumor cells as a marker for metastasis of hepatocellular carcinoma. World J. Gastroenterol. 2021, 27, 7546–7562. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Wang, Q.; Wu, Z.; Tian, X.; Yan, H.; Wang, B.; Dong, P.; Watari, H.; Pfeffer, L.M.; Guo, Y.; et al. Ovarian Primary and Metastatic Tumors Suppressed by Survivin Knockout or a Novel Survivin Inhibitor. Mol. Cancer Ther. 2019, 18, 2233–2245. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Dao, M.Q.; La, H.T. Performance of survivin mRNA as a biomarker for breast cancer among Vietnamese women. Heliyon 2019, 5, e01371. [Google Scholar] [CrossRef]
- Xu, C.; Yamamoto-Ibusuki, M.; Yamamoto, Y.; Yamamoto, S.; Fujiwara, S.; Murakami, K.; Okumura, Y.; Yamaguchi, L.; Fujiki, Y.; Iwase, H. High survivin mRNA expression is a predictor of poor prognosis in breast cancer: A comparative study at the mRNA and protein level. Breast Cancer 2014, 21, 482–490. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, X.; Shen, C.; Shi, Y. MicroRNA-203 suppresses cell proliferation and migration by targeting BIRC5 and LASP1 in human triple-negative breast cancer cells. J. Exp. Clin. Cancer Res. 2012, 31, 58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, X.; Zhao, S.; Wang, Y.; Di, W.; Zhao, G.; Yang, M.; Zhang, Q. Prognostic value of survivin and EGFR protein expression in triple-negative breast cancer (TNBC) patients. Target. Oncol. 2014, 9, 349–357. [Google Scholar] [CrossRef]
- Wu, W.; Warner, M.; Wang, L.; He, W.-W.; Zhao, R.; Guan, X.; Botero, C.; Huang, B.; Ion, C.; Coombes, C.; et al. Drivers and suppressors of triple-negative breast cancer. Proc. Natl. Acad. Sci. USA 2021, 118, e2104162118. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Z.; Wen, Q.; Xiong, Z.; Cao, X.; Zheng, Z.; Zhang, Y.; Huang, Z. An overview on the biosynthesis and metabolic regulation of monacolin K/lovastatin. Food Funct. 2020, 11, 5738–5748. [Google Scholar] [CrossRef]
- Xiong, Z.; Cao, X.; Wen, Q.; Chen, Z.; Cheng, Z.; Huang, X.; Zhang, Y.; Long, C.; Zhang, Y.; Huang, Z. An overview of the bioactivity of monacolin K / lovastatin. Food Chem. Toxicol. 2019, 131, 110585. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Zhu, G.; Shang, J.; Chen, X.; Zhang, C.; Ji, X.; Zhang, Q.; Wei, Y. An overview on the biological activity and anti-cancer mechanism of lovastatin. Cell. Signal. 2021, 87, 110122. [Google Scholar] [CrossRef] [PubMed]
- Dimitroulakos, J.; Ye, L.Y.; Benzaquen, M.; Moore, M.J.; Kamel-Reid, S.; Freedman, M.H.; Yeger, H.; Penn, L.Z. Differential sensitivity of various pediatric cancers and squamous cell carcinomas to lovastatin-induced apoptosis: Therapeutic implications. Clin. Cancer Res. 2001, 7, 158–167. [Google Scholar]
- Song, L.; Tao, X.; Lin, L.; Chen, C.; Yao, H.; He, G.; Zou, G.; Cao, Z.; Yan, S.; Lu, L.; et al. Cerasomal Lovastatin Nanohybrids for Efficient Inhibition of Triple-Negative Breast Cancer Stem Cells To Improve Therapeutic Efficacy. ACS Appl. Mater. Interfaces 2018, 10, 7022–7030. [Google Scholar] [CrossRef]
- Lee, B.-H.; Lu, Y.; Prado, M.A.; Shi, Y.; Tian, G.; Sun, S.; Elsasser, S.; Gygi, S.P.; King, R.W.; Finley, D. USP14 deubiquitinates proteasome-bound substrates that are ubiquitinated at multiple sites. Nature 2016, 532, 398–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ning, S.; Yu, B.; Wang, Y. USP14: Structure, Function, and Target Inhibition. Front Pharmacol. 2021, 12, 801328. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Tang, Q.; Efe, G.; Chiarella, A.M.; Leung, J.; Chen, M.; Yamazoe, T.; Su, Z.; Pitarresi, J.R.; Li, J.; Islam, M.; et al. Mutant p53 regulates Survivin to foster lung metastasis. Genes Dev. 2021, 35, 528–541. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Wong, B.C. Targeting apoptosis as an approach for gastrointestinal cancer therapy. Drug Resist. Updat. 2009, 12, 55–64. [Google Scholar] [CrossRef]
- Zhou, Y.; Tang, Y.; Luo, J.; Yang, Y.; Zang, H.; Ma, J.; Fan, S.; Wen, Q. High expression of HSP60 and survivin predicts poor prognosis for oral squamous cell carcinoma patients. BMC Oral Health 2023, 23, 629–638. [Google Scholar] [CrossRef]
- Li, F.; Aljahdali, I.; Ling, X. Cancer therapeutics using survivin BIRC5 as a target: What can we do after over two decades of study? J. Exp. Clin. Cancer Res. 2019, 38, 368. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, D.; Manero-Rupérez, N.; Quesada, R.; Korrodi-Gregório, L.; Soto-Cerrato, V. Therapeutic strategies involving survivin inhibition in cancer. Med. Res. Rev. 2019, 39, 887–909. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Qing, X.; Li, X.; Gan, Y.; Wang, R.; Han, S.; Li, W.; Song, W. TRAF4 regulates ubiquitination-modulated survivin turnover and confers radioresistance. Int. J. Biol. Sci. 2024, 20, 182–199. [Google Scholar] [CrossRef]
- Cao, Y.; Tang, H.; Wang, G.; Li, P.; Song, Z.; Li, W.; Sun, X.; Zhong, X.; Yu, Q.; Zhu, S.; et al. Targeting survivin with Tanshinone IIA inhibits tumor growth and overcomes chemoresistance in colorectal cancer. Cell Death Discov. 2023, 9, 351. [Google Scholar] [CrossRef]
- Lee, T.G.; Woo, S.M.; Seo, S.U.; Kim, S.; Park, J.-W.; Chang, Y.-C.; Kwon, T.K. Inhibition of USP2 Enhances TRAIL-Mediated Cancer Cell Death through Downregulation of Survivin. Int. J. Mol. Sci. 2023, 24, 12816. [Google Scholar] [CrossRef]
- Sun, X.; Angelastro, J.M.; Merino, D.; Zhou, Q.; Siegelin, M.D.; Greene, L.A. Dominant-negative ATF5 rapidly depletes survivin in tumor cells. Cell Death Dis. 2019, 10, 709. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, A.P.; Tyagi, A.; Poondla, N.; Sarodaya, N.; Karapurkar, J.K.; Kaushal, K.; Park, C.-H.; Hong, S.-H.; Kim, K.-S.; Ramakrishna, S. Dual role of deubiquitinating enzyme USP19 regulates mitotic progression and tumorigenesis by stabilizing survivin. Mol. Ther. 2022, 30, 3414–3429. [Google Scholar] [CrossRef]
- Liu, H.; Gan, Q.; Lai, Y.; Pan, Z.; Jin, Q.; Li, J.; Wang, N.; Jiao, S.; Chai, Y. USP14 increases the sensitivity of retinoblastoma to cisplatin by mediating the ferroptosis. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2024, 397, 8671–8680. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Zheng, C.; Ding, S.; Wang, Z.; Yang, Y.; Wang, Y.; He, G.; Fu, S.; Deng, X. Lovastatin Targets the USP14–Survivin Axis to Suppress Triple-Negative Breast Cancer via Ubiquitin-Mediated Proteasomal Degradation. Cells 2025, 14, 816. https://doi.org/10.3390/cells14110816
Zhou L, Zheng C, Ding S, Wang Z, Yang Y, Wang Y, He G, Fu S, Deng X. Lovastatin Targets the USP14–Survivin Axis to Suppress Triple-Negative Breast Cancer via Ubiquitin-Mediated Proteasomal Degradation. Cells. 2025; 14(11):816. https://doi.org/10.3390/cells14110816
Chicago/Turabian StyleZhou, Li, Chanjuan Zheng, Siyu Ding, Zhiyu Wang, Yiyuan Yang, Yian Wang, Guangchun He, Shujun Fu, and Xiyun Deng. 2025. "Lovastatin Targets the USP14–Survivin Axis to Suppress Triple-Negative Breast Cancer via Ubiquitin-Mediated Proteasomal Degradation" Cells 14, no. 11: 816. https://doi.org/10.3390/cells14110816
APA StyleZhou, L., Zheng, C., Ding, S., Wang, Z., Yang, Y., Wang, Y., He, G., Fu, S., & Deng, X. (2025). Lovastatin Targets the USP14–Survivin Axis to Suppress Triple-Negative Breast Cancer via Ubiquitin-Mediated Proteasomal Degradation. Cells, 14(11), 816. https://doi.org/10.3390/cells14110816