

Expression and Regulation of Hypoxia-Inducible Factor Signalling in Acute Lung Inflammation

 , ,
, ,  ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
Structure and Regulation of Hypoxia-Inducible Factors (HIFs)
2. HIF Expression and Function in Experimental Acute Lung Injury Models
2.1. HIF Expression in Experimental Acute Lung Injury Models
2.2. HIF Regulation and Inflammation in Experimental Acute Lung Injury Models
2.3. HIF Regulation and Metabolism in Experimental Acute Lung Injury Models
2.4. HIF Regulation and Vascular Remodelling in Experimental Acute Lung Injury Models
3. HIF Expression in Clinical Studies in Critically Ill Patients with ARDS
4. Targeting HIFs as Novel Therapeutic Strategies
5. Unravelling the Implication of HIF in Acute Lung Inflammation
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Crawford, A.M.; Shiferaw, A.A.; Ntambwe, P.; Milan, A.O.; Khalid, K.; Rubio, R.; Nizeyimana, F.; Ariza, F.; Mohammed, A.D.; Baker, T.; et al. Global critical care: A call to action. Crit. Care 2023, 27, 28. [Google Scholar] [CrossRef]
- McKenna, H.T.; Murray, A.J.; Martin, D.S. Human adaptation to hypoxia in critical illness. J. Appl. Physiol. 2020, 129, 656–663. [Google Scholar] [CrossRef]
- SRLF Trial Group. Hypoxemia in the ICU: Prevalence, treatment, and outcome. Ann. Intensive Care 2018, 8, 82. [Google Scholar] [CrossRef]
- Mirchandani, A.S.; Jenkins, S.J.; Bain, C.C.; Sanchez-Garcia, M.A.; Lawson, H.; Coelho, P.; Murphy, F.; Griffith, D.M.; Zhang, A.; Morrison, T.; et al. Author Correction: Hypoxia shapes the immune landscape in lung injury and promotes the persistence of inflammation. Nat. Immunol. 2022, 23, 1394. [Google Scholar] [CrossRef]
- Bos, L.D.J.; Ware, L.B. Acute respiratory distress syndrome: Causes, pathophysiology, and phenotypes. Lancet 2022, 400, 1145–1156. [Google Scholar] [CrossRef]
- Bos, L.D.J.; de Grooth, H.J.; Tuinman, P.R. A structured diagnostic algorithm for patients with ARDS. Crit. Care 2023, 27, 94. [Google Scholar] [CrossRef] [PubMed]
- Meduri, G.U.; Annane, D.; Confalonieri, M.; Chrousos, G.P.; Rochwerg, B.; Busby, A.; Ruaro, B.; Meibohm, B. Pharmacological principles guiding prolonged glucocorticoid treatment in ARDS. Intensive Care Med. 2020, 46, 2284–2296. [Google Scholar] [CrossRef]
- Vassiliou, A.G.; Kotanidou, A.; Dimopoulou, I.; Orfanos, S.E. Endothelial Damage in Acute Respiratory Distress Syndrome. Int. J. Mol. Sci. 2020, 21, 8793. [Google Scholar] [CrossRef] [PubMed]
- The ARDS Definition Task Force; Ranieri, V.M.; Rubenfeld, G.D.; Thompson, B.T.; Ferguson, N.D.; Caldwell, E.; Fan, E.; Camporota, L.; Slutsky, A.S. Acute respiratory distress syndrome: The Berlin Definition. JAMA 2012, 307, 2526–2533. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Hypoxia-inducible factor 1 (HIF-1) pathway. Sci. STKE 2007, 2007, cm8. [Google Scholar] [CrossRef]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-inducible factors and the response to hypoxic stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Du, F.; Shen, G.; Zheng, F.; Xu, B. The role of hypoxia-inducible factor-2 in digestive system cancers. Cell Death Dis. 2015, 6, e1600. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, L.A.; Yun, X.; Sikka, G. Revisiting the role of hypoxia-inducible factors in pulmonary hypertension. Curr. Opin. Physiol. 2019, 7, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Potluri, N.; Lu, J.; Kim, Y.; Rastinejad, F. Structural integration in hypoxia-inducible factors. Nature 2015, 524, 303–308. [Google Scholar] [CrossRef]
- Taylor, C.T.; Scholz, C.C. The effect of HIF on metabolism and immunity. Nat. Rev. Nephrol. 2022, 18, 573–587. [Google Scholar] [CrossRef]
- Semenza, G.L. Regulation of mammalian O2 homeostasis by hypoxia-inducible factor 1. Annu. Rev. Cell Dev. Biol. 1999, 15, 551–578. [Google Scholar] [CrossRef]
- Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J. Activation of the HIF pathway in cancer. Curr. Opin. Genet. Dev. 2001, 11, 293–299. [Google Scholar] [CrossRef]
- Tanimoto, K.; Makino, Y.; Pereira, T.; Poellinger, L. Mechanism of regulation of the hypoxia-inducible factor-1 alphaby the von Hippel-Lindau tumor suppressor protein. EMBO J. 2000, 19, 4298–4309. [Google Scholar] [CrossRef]
- Watts, E.R.; Walmsley, S.R. Inflammation and Hypoxia: HIF and PHD Isoform Selectivity. Trends Mol. Med. 2019, 25, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, K.S.; McNeill, L.A.; Riordan, M.V.; Tian, Y.M.; Bullock, A.N.; Welford, R.W.; Elkins, J.M.; Oldham, N.J.; Bhattacharya, S.; Gleadle, J.M.; et al. Hypoxia-inducible factor (HIF) asparagine hydroxylase is identical to factor inhibiting HIF (FIH) and is related to the cupin structural family. J. Biol. Chem. 2002, 277, 26351–26355. [Google Scholar] [CrossRef] [PubMed]
- Lando, D.; Peet, D.J.; Whelan, D.A.; Gorman, J.J.; Whitelaw, M.L. Asparagine hydroxylation of the HIF transactivation domain a hypoxic switch. Science 2002, 295, 858–861. [Google Scholar] [CrossRef] [PubMed]
- Mahon, P.C.; Hirota, K.; Semenza, G.L. FIH-1: A novel protein that interacts with HIF-1alpha and VHL to mediate repression of HIF-1 transcriptional activity. Genes Dev. 2001, 15, 2675–2686. [Google Scholar] [CrossRef]
- Volkova, Y.L.; Pickel, C.; Jucht, A.E.; Wenger, R.H.; Scholz, C.C. The Asparagine Hydroxylase FIH: A Unique Oxygen Sensor. Antioxid. Redox Signal. 2022, 37, 913–935. [Google Scholar] [CrossRef]
- Malkov, M.I.; Lee, C.T.; Taylor, C.T. Regulation of the Hypoxia-Inducible Factor (HIF) by Pro-Inflammatory Cytokines. Cells 2021, 10, 2340. [Google Scholar] [CrossRef]
- Luo, Z.; Tian, M.; Yang, G.; Tan, Q.; Chen, Y.; Li, G.; Zhang, Q.; Li, Y.; Wan, P.; Wu, J. Hypoxia signaling in human health and diseases: Implications and prospects for therapeutics. Signal Transduct. Target. Ther. 2022, 7, 218. [Google Scholar] [CrossRef] [PubMed]
- Bandarra, D.; Rocha, S. Tale of two transcription factors: NF-κΒ and HIF crosstalk. OA Mol. Cell Biol. 2013, 1, 1–7. [Google Scholar] [CrossRef]
- Frede, S.; Stockmann, C.; Freitag, P.; Fandrey, J. Bacterial lipopolysaccharide induces HIF-1 activation in human monocytes via p44/42 MAPK and NF-κB. Biochem. J. 2006, 396, 517–527. [Google Scholar] [CrossRef]
- Rius, J.; Guma, M.; Schachtrup, C.; Akassoglou, K.; Zinkernagel, A.S.; Nizet, V.; Johnson, R.S.; Haddad, G.G.; Karin, M. NF-kappaB links innate immunity to the hypoxic response through transcriptional regulation of HIF-1alpha. Nature 2008, 453, 807–811. [Google Scholar] [CrossRef] [PubMed]
- van Uden, P.; Kenneth, N.S.; Rocha, S. Regulation of hypoxia-inducible factor-1alpha by NF-kappaB. Biochem. J. 2008, 412, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Bandarra, D.; Biddlestone, J.; Mudie, S.; Muller, H.A.; Rocha, S. HIF-1α restricts NF-κB-dependent gene expression to control innate immunity signals. Dis. Model. Mech. 2015, 8, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Seagroves, T.N.; Ryan, H.E.; Lu, H.; Wouters, B.G.; Knapp, M.; Thibault, P.; Laderoute, K.; Johnson, R.S. Transcription factor HIF-1 is a necessary mediator of the pasteur effect in mammalian cells. Mol. Cell Biol. 2001, 21, 3436–3444. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, R.; Zhang, H.; Kim, J.W.; Shimoda, L.; Dang, C.V.; Semenza, G.L. HIF-1 regulates cytochrome oxidase subunits to optimize efficiency of respiration in hypoxic cells. Cell 2007, 129, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Sowter, H.M.; Ratcliffe, P.J.; Watson, P.; Greenberg, A.H.; Harris, A.L. HIF-1-dependent regulation of hypoxic induction of the cell death factors BNIP3 and NIX in human tumors. Cancer Res. 2001, 61, 6669–6673. [Google Scholar]
- Zhang, H.; Gao, P.; Fukuda, R.; Kumar, G.; Krishnamachary, B.; Zeller, K.I.; Dang, C.V.; Semenza, G.L. HIF-1 inhibits mitochondrial biogenesis and cellular respiration in VHL-deficient renal cell carcinoma by repression of C-MYC activity. Cancer Cell 2007, 11, 407–420. [Google Scholar] [CrossRef]
- Lotsios, N.S.; Keskinidou, C.; Jahaj, E.; Mastora, Z.; Dimopoulou, I.; Orfanos, S.E.; Vassilaki, N.; Vassiliou, A.G.; Kotanidou, A. Prognostic Value of HIF-1α-Induced Genes in Sepsis/Septic Shock. Med. Sci. 2023, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, H.; Lian, G.; Zhang, S.Y.; Wang, X.; Jiang, C. HIF1α-Induced Glycolysis Metabolism Is Essential to the Activation of Inflammatory Macrophages. Mediat. Inflamm. 2017, 2017, 9029327. [Google Scholar] [CrossRef]
- Takeda, N.; O’Dea, E.L.; Doedens, A.; Kim, J.W.; Weidemann, A.; Stockmann, C.; Asagiri, M.; Simon, M.C.; Hoffmann, A.; Johnson, R.S. Differential activation and antagonistic function of HIF-α isoforms in macrophages are essential for NO homeostasis. Genes Dev. 2010, 24, 491–501. [Google Scholar] [CrossRef]
- Vollger, L.; Akong-Moore, K.; Cox, L.; Goldmann, O.; Wang, Y.; Schafer, S.T.; Naim, H.Y.; Nizet, V.; von Kockritz-Blickwede, M. Iron-chelating agent desferrioxamine stimulates formation of neutrophil extracellular traps (NETs) in human blood-derived neutrophils. Biosci. Rep. 2016, 36, e00333. [Google Scholar] [CrossRef] [PubMed]
- McInturff, A.M.; Cody, M.J.; Elliott, E.A.; Glenn, J.W.; Rowley, J.W.; Rondina, M.T.; Yost, C.C. Mammalian target of rapamycin regulates neutrophil extracellular trap formation via induction of hypoxia-inducible factor 1 α. Blood 2012, 120, 3118–3125. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, S.R.; Cowburn, A.S.; Clatworthy, M.R.; Morrell, N.W.; Roper, E.C.; Singleton, V.; Maxwell, P.; Whyte, M.K.; Chilvers, E.R. Neutrophils from patients with heterozygous germline mutations in the von Hippel Lindau protein (pVHL) display delayed apoptosis and enhanced bacterial phagocytosis. Blood 2006, 108, 3176–3178. [Google Scholar] [CrossRef]
- Mecklenburgh, K.I.; Walmsley, S.R.; Cowburn, A.S.; Wiesener, M.; Reed, B.J.; Upton, P.D.; Deighton, J.; Greening, A.P.; Chilvers, E.R. Involvement of a ferroprotein sensor in hypoxia-mediated inhibition of neutrophil apoptosis. Blood 2002, 100, 3008–3016. [Google Scholar] [CrossRef]
- Jantsch, J.; Chakravortty, D.; Turza, N.; Prechtel, A.T.; Buchholz, B.; Gerlach, R.G.; Volke, M.; Glasner, J.; Warnecke, C.; Wiesener, M.S.; et al. Hypoxia and hypoxia-inducible factor-1 alpha modulate lipopolysaccharide-induced dendritic cell activation and function. J. Immunol. 2008, 180, 4697–4705. [Google Scholar] [CrossRef] [PubMed]
- Paardekooper, L.M.; Bendix, M.B.; Ottria, A.; de Haer, L.W.; Ter Beest, M.; Radstake, T.; Marut, W.; van den Bogaart, G. Hypoxia potentiates monocyte-derived dendritic cells for release of tumor necrosis factor α via MAP3K8. Biosci. Rep. 2018, 38, BSR20182019. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, X.; Chen, K.; Cheng, Y.; Liu, S.; Xia, M.; Chen, Y.; Zhu, H.; Li, Z.; Cao, X. CCR7 Chemokine Receptor-Inducible lnc-Dpf3 Restrains Dendritic Cell Migration by Inhibiting HIF-1α-Mediated Glycolysis. Immunity 2019, 50, 600–615.e15. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Frevert, C.W.; Martin, T.R. Animal models of acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 295, L379–L399. [Google Scholar] [CrossRef] [PubMed]
- Keskinidou, C.; Vassiliou, A.G.; Dimopoulou, I.; Kotanidou, A.; Orfanos, S.E. Mechanistic Understanding of Lung Inflammation: Recent Advances and Emerging Techniques. J. Inflamm. Res. 2022, 15, 3501–3546. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Zhang, M.; Fu, D.; Luo, H.; Yang, X. SPP1 exacerbates ARDS via elevating Th17/Treg and M1/M2 ratios through suppression of ubiquitination-dependent HIF-1α degradation. Cytokine 2023, 164, 156107. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.J.; Liu, T.; Yang, H.H.; Duan, J.X.; Yang, J.T.; Guan, X.X.; Xiong, J.B.; Zhang, Y.F.; Zhang, C.Y.; Zhou, Y.; et al. TREM-1 governs NLRP3 inflammasome activation of macrophages by firing up glycolysis in acute lung injury. Int. J. Biol. Sci. 2023, 19, 242–257. [Google Scholar] [CrossRef]
- Liu, F.C.; Yang, Y.H.; Liao, C.C.; Lee, H.C. Xanthoxylin Attenuates Lipopolysaccharide-Induced Lung Injury through Modulation of Akt/HIF-1α/NF-κB and Nrf2 Pathways. Int. J. Mol. Sci. 2024, 25, 8742. [Google Scholar] [CrossRef]
- Shang, L.; Zhang, M.; Li, J.; Zhou, F.; Wang, S.; Chen, L.; Yang, S. Dachengqi decoction alleviates acute lung injury by suppressing HIF-1α-mediated glycolysis. J. Ethnopharmacol. 2024, 321, 117410. [Google Scholar] [CrossRef]
- Tsai, Y.M.; Lin, Y.C.; Chen, C.Y.; Chien, H.C.; Chang, H.; Chiang, M.H. Leveraging Hypotension Prediction Index to Forecast LPS-Induced Acute Lung Injury and Inflammation in a Porcine Model: Exploring the Role of Hypoxia-Inducible Factor in Circulatory Shock. Biomedicines 2024, 12, 1665. [Google Scholar] [CrossRef]
- Wang, Y.M.; Qi, X.; Gong, F.C.; Chen, Y.; Yang, Z.T.; Mao, E.Q.; Chen, E.Z. Protective and predictive role of Mucin1 in sepsis-induced ALI/ARDS. Int. Immunopharmacol. 2020, 83, 106438. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.J.; Mirchandani, A.S.; Lynch, R.W.; Murphy, F.; Delaney, L.; Small, D.; Coelho, P.; Watts, E.R.; Sadiku, P.; Griffith, D.; et al. IL4Rα Signaling Abrogates Hypoxic Neutrophil Survival and Limits Acute Lung Injury Responses In Vivo. Am. J. Respir. Crit. Care Med. 2019, 200, 235–246. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, X.; Zhao, D.X.; Yin, J.; Hu, G.; Evans, C.E.; Zhao, Y. Endothelial Hypoxia-Inducible Factor-1α Is Required for Vascular Repair and Resolution of Inflammatory Lung Injury through Forkhead Box Protein M1. Am. J. Pathol. 2019, 189, 1664–1679. [Google Scholar] [CrossRef] [PubMed]
- Miao, R.F.; Tu, J. LncRNA CDKN2B-AS1 interacts with LIN28B to exacerbate sepsis-induced acute lung injury by inducing HIF-1α/NLRP3-mediated pyroptosis. Kaohsiung J. Med. Sci. 2023, 39, 883–895. [Google Scholar] [CrossRef]
- Sherman, M.A.; Suresh, M.V.; Dolgachev, V.A.; McCandless, L.K.; Xue, X.; Ziru, L.; Machado-Aranda, D.; Shah, Y.M.; Raghavendran, K. Molecular Characterization of Hypoxic Alveolar Epithelial Cells After Lung Contusion Indicates an Important Role for HIF-1α. Ann. Surg. 2018, 267, 382–391. [Google Scholar] [CrossRef]
- Suresh, M.V.; Balijepalli, S.; Zhang, B.; Singh, V.V.; Swamy, S.; Panicker, S.; Dolgachev, V.A.; Subramanian, C.; Ramakrishnan, S.K.; Thomas, B.; et al. Hypoxia-Inducible Factor (HIF)-1α Promotes Inflammation and Injury Following Aspiration-Induced Lung Injury in Mice. Shock 2019, 52, 612–621. [Google Scholar] [CrossRef]
- Liu, C.; Pei, S.; Dai, H.; Liu, Z.; Ye, M.; Liu, H.; He, X.; Wu, S.; Qin, Y.; Lin, F. Downregulation of SIRT3 Aggravates Lung Ischemia Reperfusion Injury by Increasing Mitochondrial Fission and Oxidative Stress through HIF-1α-Dependent Mechanisms. Oxidative Med. Cell Longev. 2022, 2022, 9041914. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Yang, Y. Parecoxib sodium alleviates ischemia reperfusion-induced pulmonary injury via inhibiting ERK/NF-κB and further activating the HIF-1α pathway. Immun. Inflamm. Dis. 2022, 10, e684. [Google Scholar] [CrossRef]
- Lu, Z.L.; Song, C.K.; Zou, S.S.; Pan, S.Z.; Lai, K.; Li, N.; Geng, Q. Hydroxycitric Acid Alleviated Lung Ischemia-Reperfusion Injury by Inhibiting Oxidative Stress and Ferroptosis through the Hif-1α Pathway. Curr. Issues Mol. Biol. 2023, 45, 9868–9886. [Google Scholar] [CrossRef]
- Grover, T.R.; Asikainen, T.M.; Kinsella, J.P.; Abman, S.H.; White, C.W. Hypoxia-inducible factors HIF-1α and HIF-2α are decreased in an experimental model of severe respiratory distress syndrome in preterm lambs. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 292, L1345–L1351. [Google Scholar] [CrossRef] [PubMed]
- Eckle, T.; Brodsky, K.; Bonney, M.; Packard, T.; Han, J.; Borchers, C.H.; Mariani, T.J.; Kominsky, D.J.; Mittelbronn, M.; Eltzschig, H.K. HIF1A reduces acute lung injury by optimizing carbohydrate metabolism in the alveolar epithelium. PLoS Biol. 2013, 11, e1001665. [Google Scholar] [CrossRef] [PubMed]
- Suresh, M.V.; Yalamanchili, G.; Rao, T.C.; Aktay, S.; Kralovich, A.; Shah, Y.M.; Raghavendran, K. Hypoxia-inducible factor (HIF)-1α-induced regulation of lung injury in pulmonary aspiration is mediated through NF-kB. FASEB BioAdv. 2022, 4, 309–328. [Google Scholar] [CrossRef]
- Berg, N.K.; Li, J.; Kim, B.; Mills, T.; Pei, G.; Zhao, Z.; Li, X.; Zhang, X.; Ruan, W.; Eltzschig, H.K.; et al. Hypoxia-inducible factor-dependent induction of myeloid-derived netrin-1 attenuates natural killer cell infiltration during endotoxin-induced lung injury. FASEB J. 2021, 35, e21334. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Fan, C.; Ji, Y.; Su, Q.; Zhao, F.; Xie, C.; Chen, X.; Zhang, Y.; Chen, Y. SENP3 facilitates M1 macrophage polarization via the HIF-1α/PKM2 axis in lipopolysaccharide-induced acute lung injury. Innate Immun. 2023, 29, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Tojo, K.; Tamada, N.; Nagamine, Y.; Yazawa, T.; Ota, S.; Goto, T. Enhancement of glycolysis by inhibition of oxygen-sensing prolyl hydroxylases protects alveolar epithelial cells from acute lung injury. FASEB J. 2018, 32, 2258–2268. [Google Scholar] [CrossRef] [PubMed]
- Woods, P.S.; Kimmig, L.M.; Sun, K.A.; Meliton, A.Y.; Shamaa, O.R.; Tian, Y.; Cetin-Atalay, R.; Sharp, W.W.; Hamanaka, R.B.; Mutlu, G.M. HIF-1α induces glycolytic reprograming in tissue-resident alveolar macrophages to promote cell survival during acute lung injury. eLife 2022, 11, e77457. [Google Scholar] [CrossRef]
- McClendon, J.; Jansing, N.L.; Redente, E.F.; Gandjeva, A.; Ito, Y.; Colgan, S.P.; Ahmad, A.; Riches, D.W.H.; Chapman, H.A.; Mason, R.J.; et al. Hypoxia-Inducible Factor 1α Signaling Promotes Repair of the Alveolar Epithelium after Acute Lung Injury. Am. J. Pathol. 2017, 187, 1772–1786. [Google Scholar] [CrossRef]
- Liu, M.; Liu, Q.; Pei, Y.; Gong, M.; Cui, X.; Pan, J.; Zhang, Y.; Liu, Y.; Liu, Y.; Yuan, X.; et al. Aqp-1 Gene Knockout Attenuates Hypoxic Pulmonary Hypertension of Mice. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 48–62. [Google Scholar] [CrossRef]
- Keskinidou, C.; Lotsios, N.S.; Vassiliou, A.G.; Dimopoulou, I.; Kotanidou, A.; Orfanos, S.E. The Interplay between Aquaporin-1 and the Hypoxia-Inducible Factor 1α in a Lipopolysaccharide-Induced Lung Injury Model in Human Pulmonary Microvascular Endothelial Cells. Int. J. Mol. Sci. 2022, 23, 10588. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, S.; Chen, M.; Liao, W.; Er, S.Y.; Wong, W.S.; Ge, R. Isthmin is a novel vascular permeability inducer that functions through cell-surface GRP78-mediated Src activation. Cardiovasc. Res. 2015, 107, 131–142. [Google Scholar] [CrossRef]
- Li, J.; Xia, Y.; Huang, Z.; Zhao, Y.; Xiong, R.; Li, X.; Huang, Q.; Shan, F. Novel HIF-1-target gene isthmin1 contributes to hypoxia-induced hyperpermeability of pulmonary microvascular endothelial cells monolayers. Am. J. Physiol. Cell Physiol. 2021, 321, C671–C680. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.R.; Li, Y.Q.; Chen, G.S.; MA, D.Y. Relationships between serum HIF-1α and VEGF levels and clinical prognosis of acute respiratory distress syndrome. J. Trop. Med. 2019, 19, 886–889. [Google Scholar]
- Hussain, L. Assessment of Serum Hypoxia-Inducible Factor-1 and Chemerin Levels as Potential Markers of Severity in COVID-19 Patients. HIV Nurs. 2022, 22, 993–1000. [Google Scholar]
- Tian, M.; Liu, W.; Li, X.; Zhao, P.; Shereen, M.A.; Zhu, C.; Huang, S.; Liu, S.; Yu, X.; Yue, M.; et al. HIF-1α promotes SARS-CoV-2 infection and aggravates inflammatory responses to COVID-19. Signal Transduct. Target. Ther. 2021, 6, 308. [Google Scholar] [CrossRef]
- Yilmaz, S.; Kuskucu, A.; Horoz, O.O.; Suakar, O.; Imamova, N.; Gongor, G.; Yildizdas, D. Polymorphism of hypoxia-inducible factor-1 α gene in pediatric acute respiratory distress syndrome. J. Acute Dis. 2019, 8, 67–71. [Google Scholar] [CrossRef]
- Yang, X.; Zhu, H.C.; Zhang, C.; Qin, Q.; Liu, J.; Xu, L.P.; Zhao, L.J.; Zhang, Q.; Cai, J.; Ma, J.X.; et al. HIF-1α 1772 C/T and 1790 G/A polymorphisms are significantly associated with higher cancer risk: An updated meta-analysis from 34 case-control studies. PLoS ONE 2013, 8, e80396. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.T.; Li, B.; Chen, M.; Jia, H.R.; Zhang, H.X. Associations between HIF-1α polymorphisms C1772T and G1790A and susceptibility to chronic obstructive pulmonary disease. Genet. Mol. Res. 2015, 14, 17341–17347. [Google Scholar] [CrossRef]
- Sipahioglu, H.; Sumeyra, K.; Akalin, H.; Karasu, N.; Sipahioglu, M.; Eren, E.E.; Onuk, S.; Elay, G.; Saatci, C.; Yuksel, R.C.; et al. HIF-1α is Associated with Improved Survival in ARDS due to COVID-19: A Prospective Study. Research Square 2024. [Google Scholar] [CrossRef]
- Dotsch, A.; Eisele, L.; Rabeling, M.; Rump, K.; Walstein, K.; Bick, A.; Cox, L.; Engler, A.; Bachmann, H.S.; Jockel, K.H.; et al. Hypoxia Inducible Factor-2 Alpha and Prolinhydroxylase 2 Polymorphisms in Patients with Acute Respiratory Distress Syndrome (ARDS). Int. J. Mol. Sci. 2017, 18, 1266. [Google Scholar] [CrossRef]
- Hocker, A.; Rabeling, M.; Bick, A.; Cox, L.; Kreuzer, M.; Engler, A.; Walstein, K.; Bachmann, H.S.; Jockel, K.H.; Eisele, L.; et al. Hypoxia inducible factor-1 alpha and prolinhydroxlase 2 polymorphisms in patients with severe sepsis: A prospective observational trial. BMC Anesthesiol. 2016, 16, 61. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Gao, W.; Chen, R.; Chen, S.; Gu, X.; Wang, F.; Li, Q. CircRNAs in BALF exosomes and plasma as diagnostic biomarkers in patients with acute respiratory distress syndrome caused by severe pneumonia. Front. Cell Infect. Microbiol. 2023, 13, 1194495. [Google Scholar] [CrossRef]
- Khan, Y.A.; Fan, E.; Ferguson, N.D. Precision Medicine and Heterogeneity of Treatment Effect in Therapies for ARDS. Chest 2021, 160, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Ruan, W.; Bobrow, B.; Carmeliet, P.; Eltzschig, H.K. Targeting hypoxia-inducible factors: Therapeutic opportunities and challenges. Nat. Rev. Drug Discov. 2024, 23, 175–200. [Google Scholar] [CrossRef]
- Ruan, H.; Zhang, Q.; Zhang, Y.-p.; Li, S.-s.; Ran, X. Unraveling the role of HIF-1α in sepsis: From pathophysiology to potential therapeutics—A narrative review. Crit. Care 2024, 28, 100. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Wang, F.; Yang, H.; Wang, Z. Action Sites and Clinical Application of HIF-1α Inhibitors. Molecules 2022, 27, 3426. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xiang, D.; Zhang, H.; Yao, H.; Wang, Y. Hypoxia-Inducible Factor-1: A Potential Target to Treat Acute Lung Injury. Oxidative Med. Cell. Longev. 2020, 2020, 8871476. [Google Scholar] [CrossRef]
- Suresh, M.V.; Balijepalli, S.; Solanki, S.; Aktay, S.; Choudhary, K.; Shah, Y.M.; Raghavendran, K. Hypoxia-Inducible Factor 1α and Its Role in Lung Injury: Adaptive or Maladaptive. Inflammation 2023, 46, 491–508. [Google Scholar] [CrossRef]
- Han, F.; Wu, G.; Han, S.; Li, Z.; Jia, Y.; Bai, L.; Li, X.; Wang, K.; Yang, F.; Zhang, J.; et al. Hypoxia-inducible factor prolyl-hydroxylase inhibitor roxadustat (FG-4592) alleviates sepsis-induced acute lung injury. Respir. Physiol. Neurobiol. 2020, 281, 103506. [Google Scholar] [CrossRef]
- Gong, H.; Rehman, J.; Tang, H.; Wary, K.; Mittal, M.; Chaturvedi, P.; Zhao, Y.Y.; Komarova, Y.A.; Vogel, S.M.; Malik, A.B. HIF2α signaling inhibits adherens junctional disruption in acute lung injury. J. Clin. Investig. 2015, 125, 652–664. [Google Scholar] [CrossRef]
- Li, J.; Haase, V.H.; Hao, C.M. Updates on Hypoxia-Inducible Factor Prolyl Hydroxylase Inhibitors in the Treatment of Renal Anemia. Kidney Dis. 2023, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- NCT04463602, Desidustat in the Management of COVID-19 Patients. Available online: https://clinicaltrials.gov/show/NCT04463602 (accessed on 30 November 2024).
- NCT04478071, Vadadustat for the Prevention and Treatment of Acute Respiratory Distress Syndrome (ARDS) in Hospitalized Patients With Coronavirus Disease 2019 (COVID-19). Available online: https://clinicaltrials.gov/show/NCT04478071 (accessed on 30 November 2024).
- Poloznikov, A.A.; Nersisyan, S.A.; Hushpulian, D.M.; Kazakov, E.H.; Tonevitsky, A.G.; Kazakov, S.V.; Vechorko, V.I.; Nikulin, S.V.; Makarova, J.A.; Gazaryan, I.G. HIF Prolyl Hydroxylase Inhibitors for COVID-19 Treatment: Pros and Cons. Front. Pharmacol. 2021, 11, 621054. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.E.; Peng, Y.; Zhu, M.M.; Dai, Z.; Zhang, X.; Zhao, Y.Y. Rabeprazole Promotes Vascular Repair and Resolution of Sepsis-Induced Inflammatory Lung Injury through HIF-1α. Cells 2022, 11, 1425. [Google Scholar] [CrossRef] [PubMed]
- Antunes, K.H.; Fachi, J.L.; de Paula, R.; da Silva, E.F.; Pral, L.P.; Dos Santos, A.; Dias, G.B.M.; Vargas, J.E.; Puga, R.; Mayer, F.Q.; et al. Microbiota-derived acetate protects against respiratory syncytial virus infection through a GPR43-type 1 interferon response. Nat. Commun. 2019, 10, 3273. [Google Scholar] [CrossRef]
- Galvão, I.; Tavares, L.P.; Corrêa, R.O.; Fachi, J.L.; Rocha, V.M.; Rungue, M.; Garcia, C.C.; Cassali, G.; Ferreira, C.M.; Martins, F.S.; et al. The Metabolic Sensor GPR43 Receptor Plays a Role in the Control of Klebsiella pneumoniae Infection in the Lung. Front. Immunol. 2018, 9, 142. [Google Scholar] [CrossRef]
- Machado, M.G.; Patente, T.A.; Rouillé, Y.; Heumel, S.; Melo, E.M.; Deruyter, L.; Pourcet, B.; Sencio, V.; Teixeira, M.M.; Trottein, F. Acetate Improves the Killing of Streptococcus pneumoniae by Alveolar Macrophages via NLRP3 Inflammasome and Glycolysis-HIF-1α Axis. Front. Immunol. 2022, 13, 773261. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xu, Y.; Lu, B.; Huang, Y.; Xu, S.; Xie, J.; Liu, M.; Che, D.; Ma, L.; Tao, J.; et al. Inositol Alleviates Pulmonary Fibrosis by Promoting Autophagy via Inhibiting the HIF-1α-SLUG Axis in Acute Respiratory Distress Syndrome. Oxidative Med. Cell. Longev. 2022, 2022, 1030238. [Google Scholar] [CrossRef] [PubMed]
- Stompor-Gorący, M. The Health Benefits of Emodin, a Natural Anthraquinone Derived from Rhubarb-A Summary Update. Int. J. Mol. Sci. 2021, 22, 9522. [Google Scholar] [CrossRef]
- Li, X.; Shan, C.; Wu, Z.; Yu, H.; Yang, A.; Tan, B. Emodin alleviated pulmonary inflammation in rats with LPS-induced acute lung injury through inhibiting the mTOR/HIF-1α/VEGF signaling pathway. Inflamm. Res. 2020, 69, 365–373. [Google Scholar] [CrossRef]
- Chen, Q.; Shao, X.; He, Y.; Lu, E.; Zhu, L.; Tang, W. Norisoboldine Attenuates Sepsis-Induced Acute Lung Injury by Modulating Macrophage Polarization via PKM2/HIF-1α/PGC-1α Pathway. Biol. Pharm. Bull. 2021, 44, 1536–1547. [Google Scholar] [CrossRef]
- Figarella, K.; Kim, J.; Ruan, W.; Mills, T.; Eltzschig, H.K.; Yuan, X. Hypoxia-adenosine axis as therapeutic targets for acute respiratory distress syndrome. Front. Immunol. 2024, 15, 1328565. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.E. Hypoxia-Inducible Factor Signaling in Inflammatory Lung Injury and Repair. Cells 2022, 11, 183. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
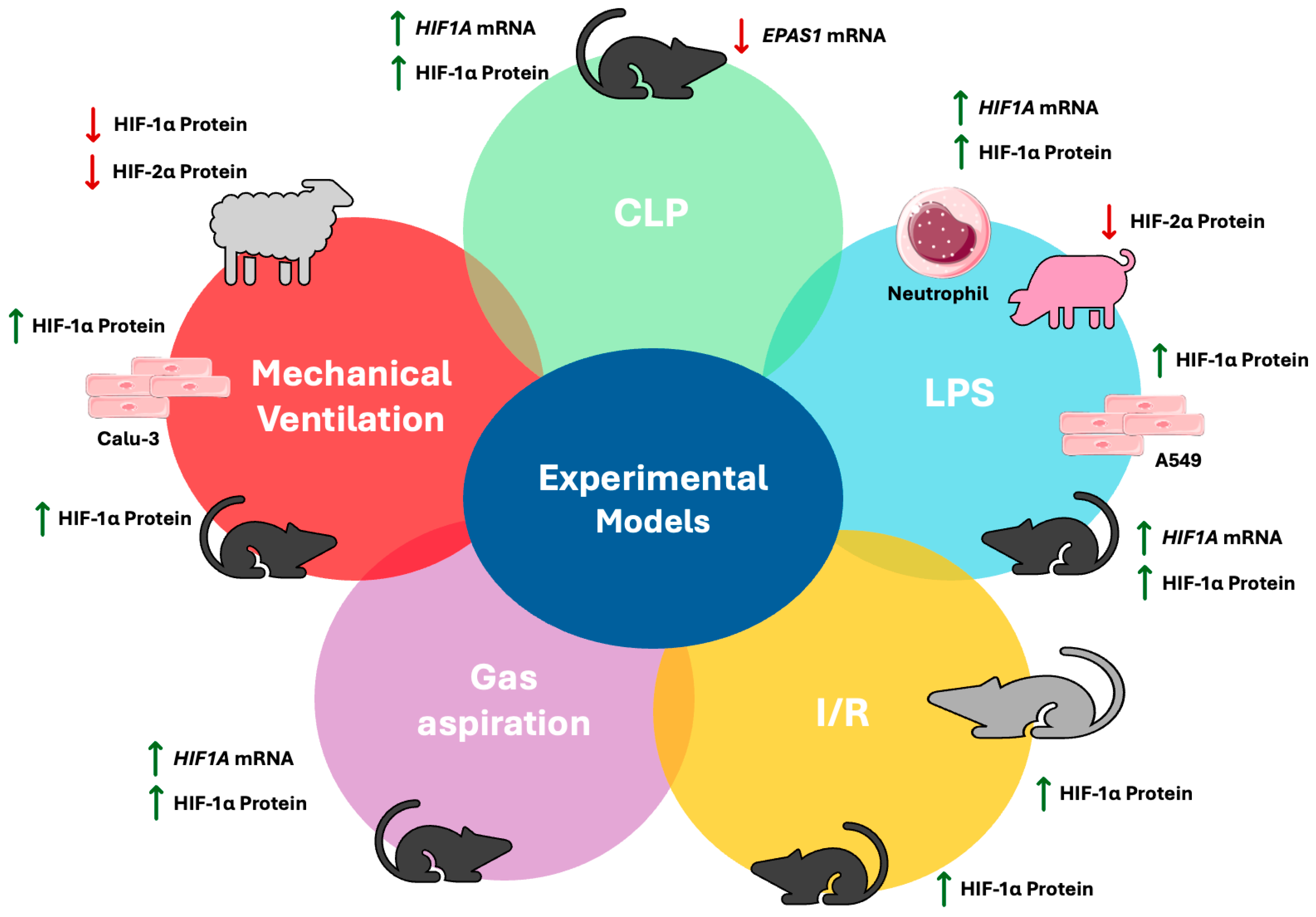
| HIF Isoform | Experimental Model | HIF Expression | References |
|---|---|---|---|
| HIF-1α | LPS mice | Increased mRNA and protein expression in lung tissue | [47] |
| LPS mice | Increased protein expression in lung tissue | [48,49,50] | |
| LPS pigs | No changes in serum | [51] | |
| LPS A549 cells | Increased protein expression | [52] | |
| LPS isolated human neutrophils | Increased mRNA and protein expression | [53] | |
| CLP mice | Increased mRNA and protein expression in lung tissue | [54] | |
| CLP mice | Increased protein expression in lung tissue | [52,55] | |
| HCl aspiration mice | Increased mRNA expression in lung lysates | [56] | |
| HCl aspiration mice | Increased protein expression in lung lysates | [57] | |
| Ischemia reperfusion mice | Increased protein expression in lung tissue | [58,60] | |
| Ischemia reperfusion rats | Increased protein expression in pulmonary tissue | [59] | |
| Mechanical ventilation preterm lambs | Decreased protein expression in lung samples | [61] | |
| Mechanical ventilation mice | Increased protein expression in lung tissue | [62] | |
| Cyclic mechanical stretching Calu-3 cells | Increased protein expression | [62] | |
| HIF-2α | LPS pigs | Decreased protein expression in serum | [51] |
| CLP mice | Decreased mRNA expression in lung tissue | [54] | |
| Mechanical ventilation preterm lambs | Decreased protein expression in lung samples | [61] |
| HIF Isoform | Overview of the Study | Findings on HIF Expression | Reference |
|---|---|---|---|
| To explore the relationship of HIF-1α with the prognosis of ARDS. HIF-1α levels were measured in the sera of 98 ARDS patients, of whom 50 patients survived, and in 30 healthy volunteers | Increased protein expression in ARDS patients vs healthy controls. Increased protein expression in non-survivors vs survivors | [73] | |
| HIF-1α | To examine whether HIF-1 is an early indicator of COVID-19 severity. HIF-1 was measured in the sera of 17 critically ill, 33 severe, and 40 mild/moderate COVID-19-induced ARDS patients and in 90 healthy volunteers | COVID-19 patients had significantly higher levels of HIF-1 than healthy volunteers. A progressive increase in HIF-1 serum levels was found according to ARDS severity in COVID-19 patients | [74] |
| Whole-transcriptome RNA sequencing analysis was performed to explore altered gene expression mediated by SARS-CoV-2 infection. Isolated PBMCs from 11 COVID-19 patients and 9 healthy individuals | Increased mRNA expression in COVID-19 patients | [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lotsios, N.S.; Keskinidou, C.; Karagiannis, S.P.; Papavassiliou, K.A.; Papavassiliou, A.G.; Kotanidou, A.; Dimopoulou, I.; Orfanos, S.E.; Vassiliou, A.G. Expression and Regulation of Hypoxia-Inducible Factor Signalling in Acute Lung Inflammation. Cells 2025, 14, 29. https://doi.org/10.3390/cells14010029
Lotsios NS, Keskinidou C, Karagiannis SP, Papavassiliou KA, Papavassiliou AG, Kotanidou A, Dimopoulou I, Orfanos SE, Vassiliou AG. Expression and Regulation of Hypoxia-Inducible Factor Signalling in Acute Lung Inflammation. Cells. 2025; 14(1):29. https://doi.org/10.3390/cells14010029
Chicago/Turabian StyleLotsios, Nikolaos S., Chrysi Keskinidou, Sotirios P. Karagiannis, Kostas A. Papavassiliou, Athanasios G. Papavassiliou, Anastasia Kotanidou, Ioanna Dimopoulou, Stylianos E. Orfanos, and Alice G. Vassiliou. 2025. "Expression and Regulation of Hypoxia-Inducible Factor Signalling in Acute Lung Inflammation" Cells 14, no. 1: 29. https://doi.org/10.3390/cells14010029
APA StyleLotsios, N. S., Keskinidou, C., Karagiannis, S. P., Papavassiliou, K. A., Papavassiliou, A. G., Kotanidou, A., Dimopoulou, I., Orfanos, S. E., & Vassiliou, A. G. (2025). Expression and Regulation of Hypoxia-Inducible Factor Signalling in Acute Lung Inflammation. Cells, 14(1), 29. https://doi.org/10.3390/cells14010029