
The Origin and Regulation of Neuromesodermal Progenitors (NMPs) in Embryos


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. The Tissue Organization of the Sinus Rhomboidalis, Where NMPs Undergo a Neural vs. Mesodermal Dichotomous Fate Choice
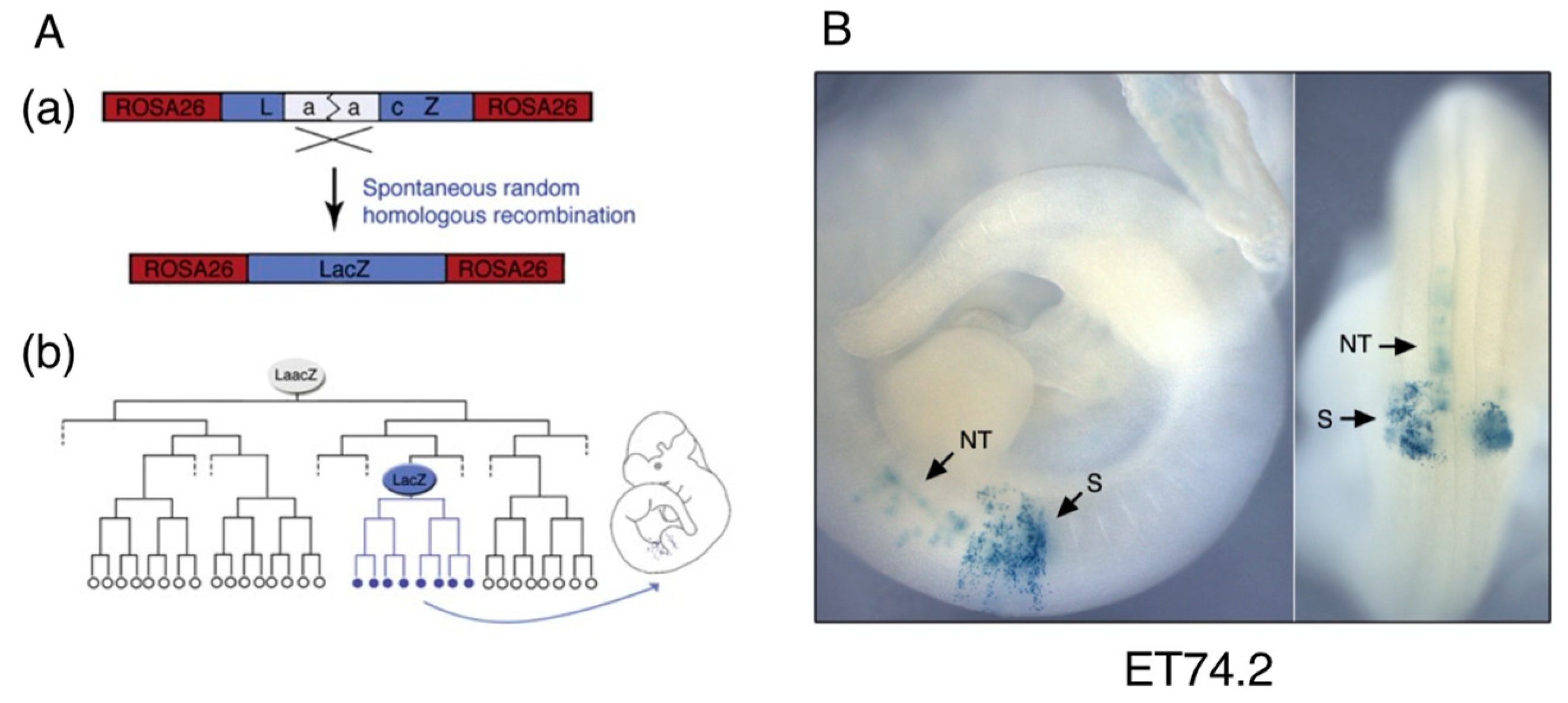
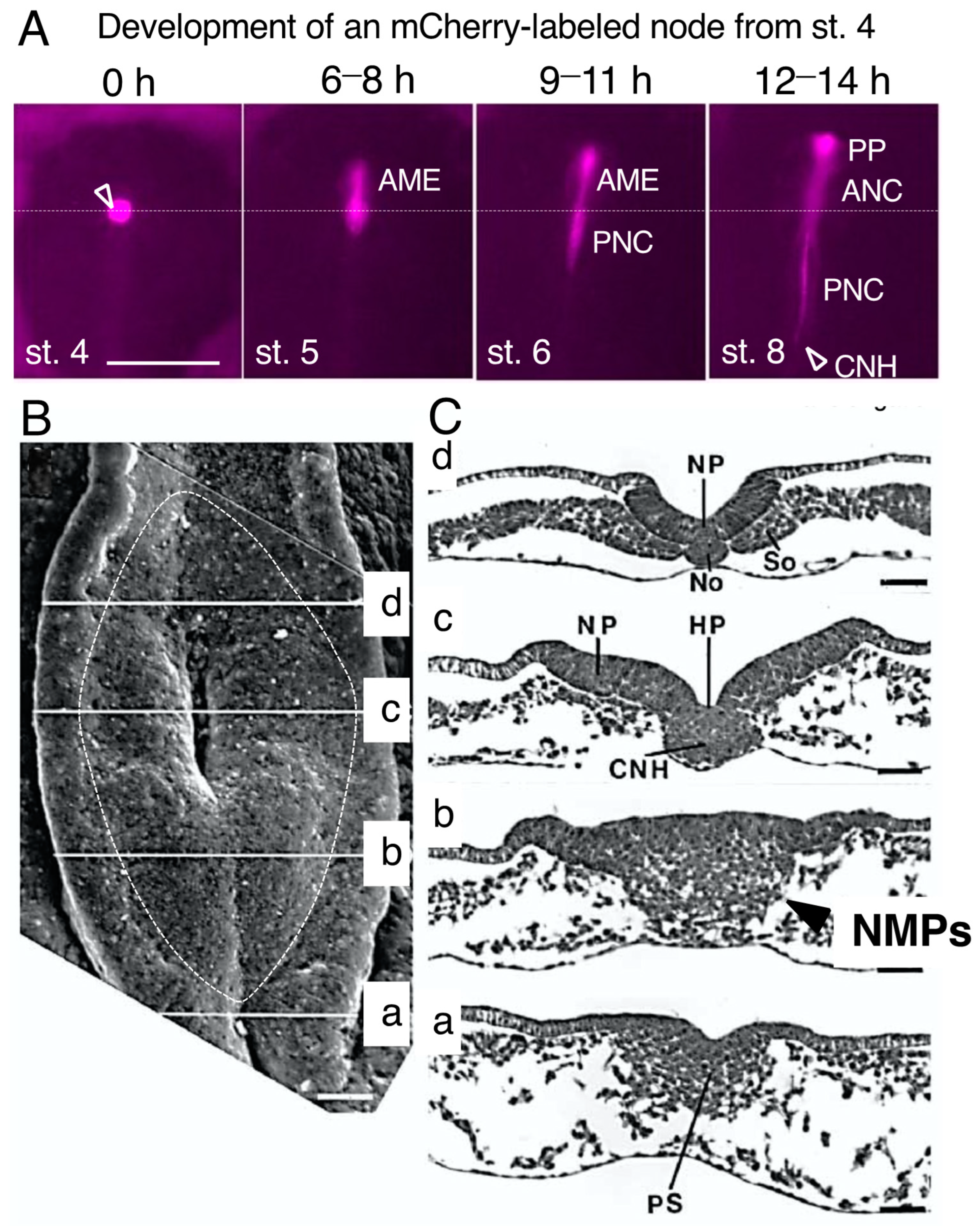
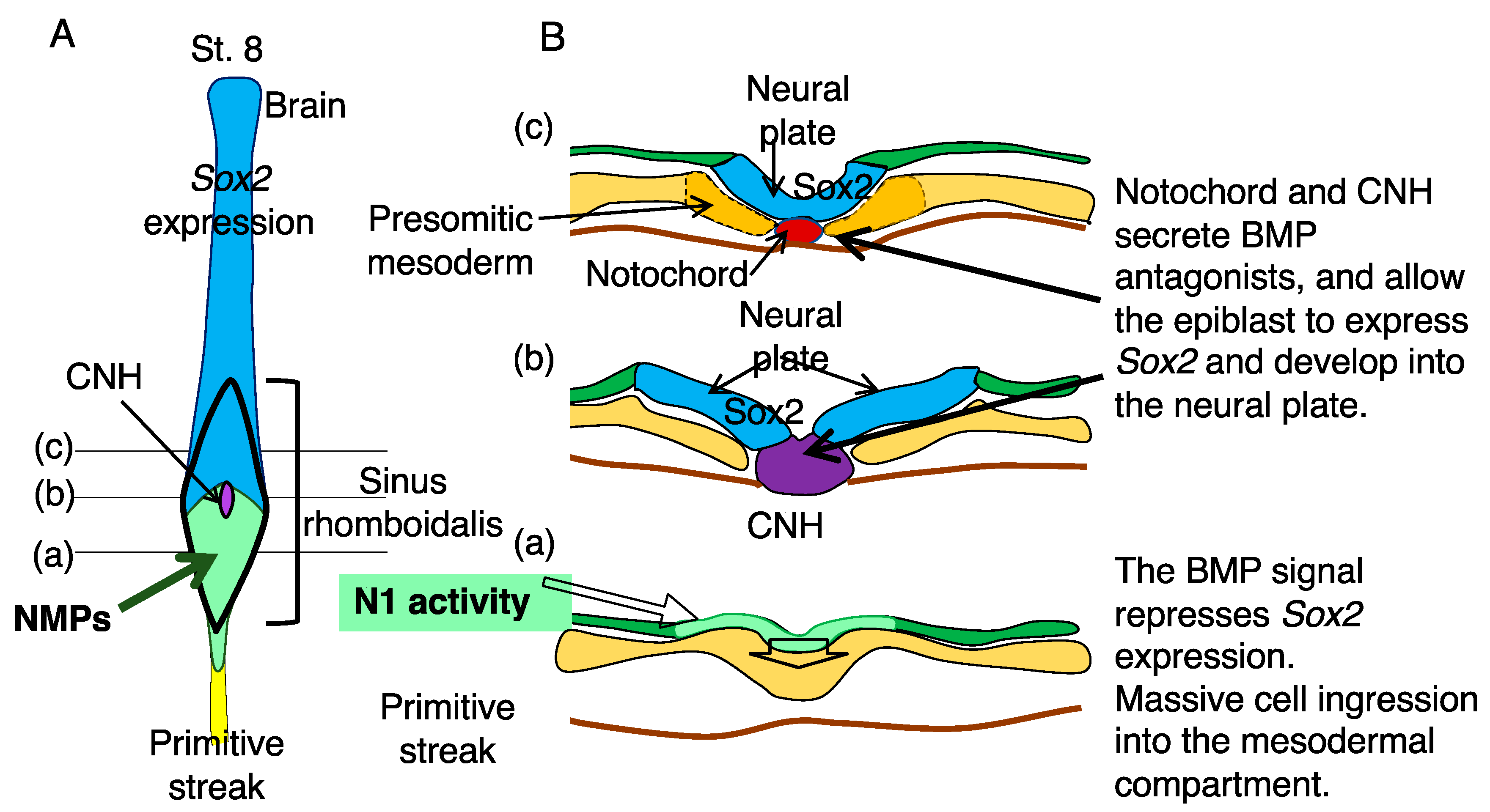
2.1. The Distinction between the Node and the Chordoneural Hinge (CNH)
2.2. Sinus Rhomboidalis Tissue Organization
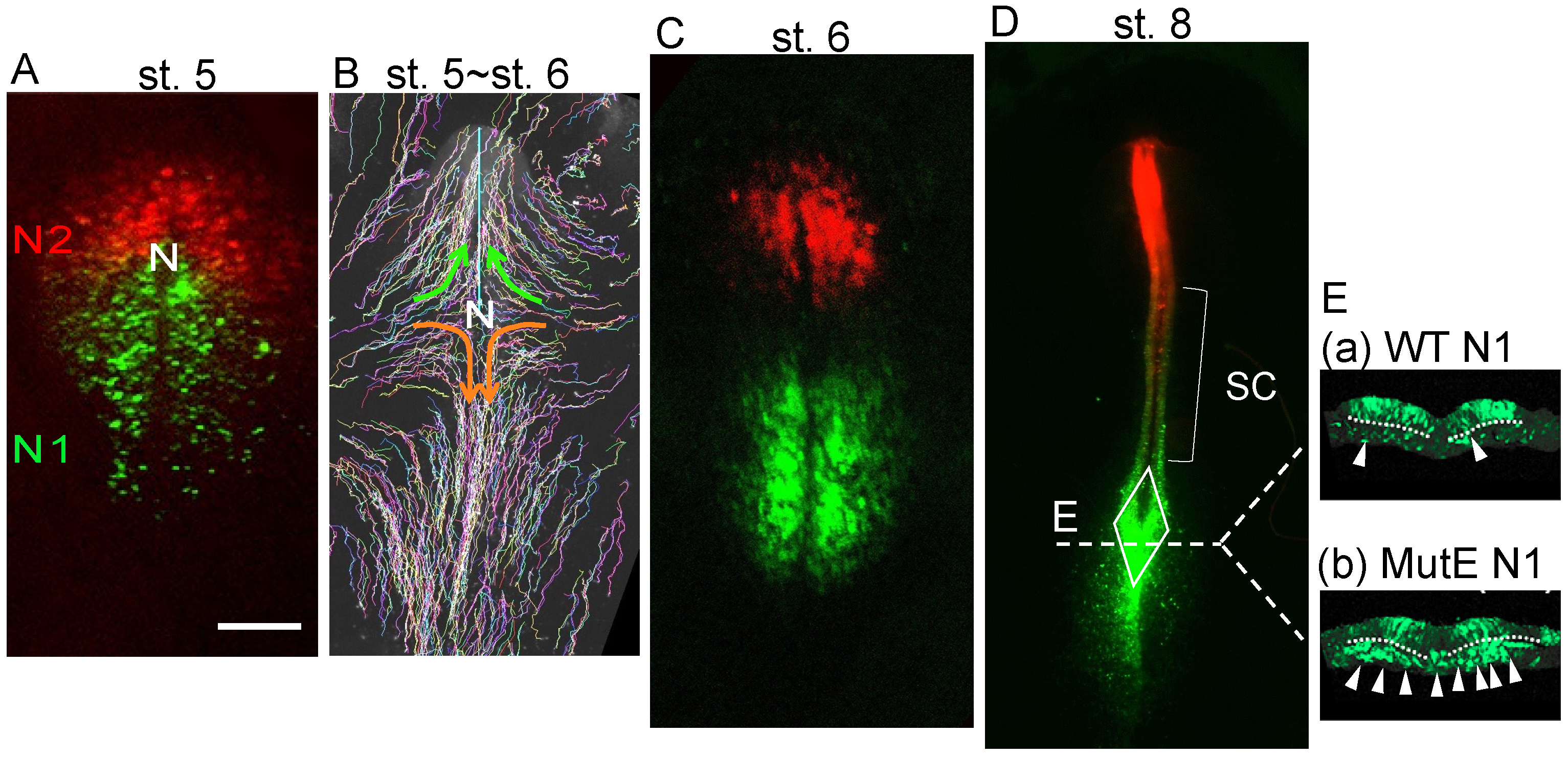
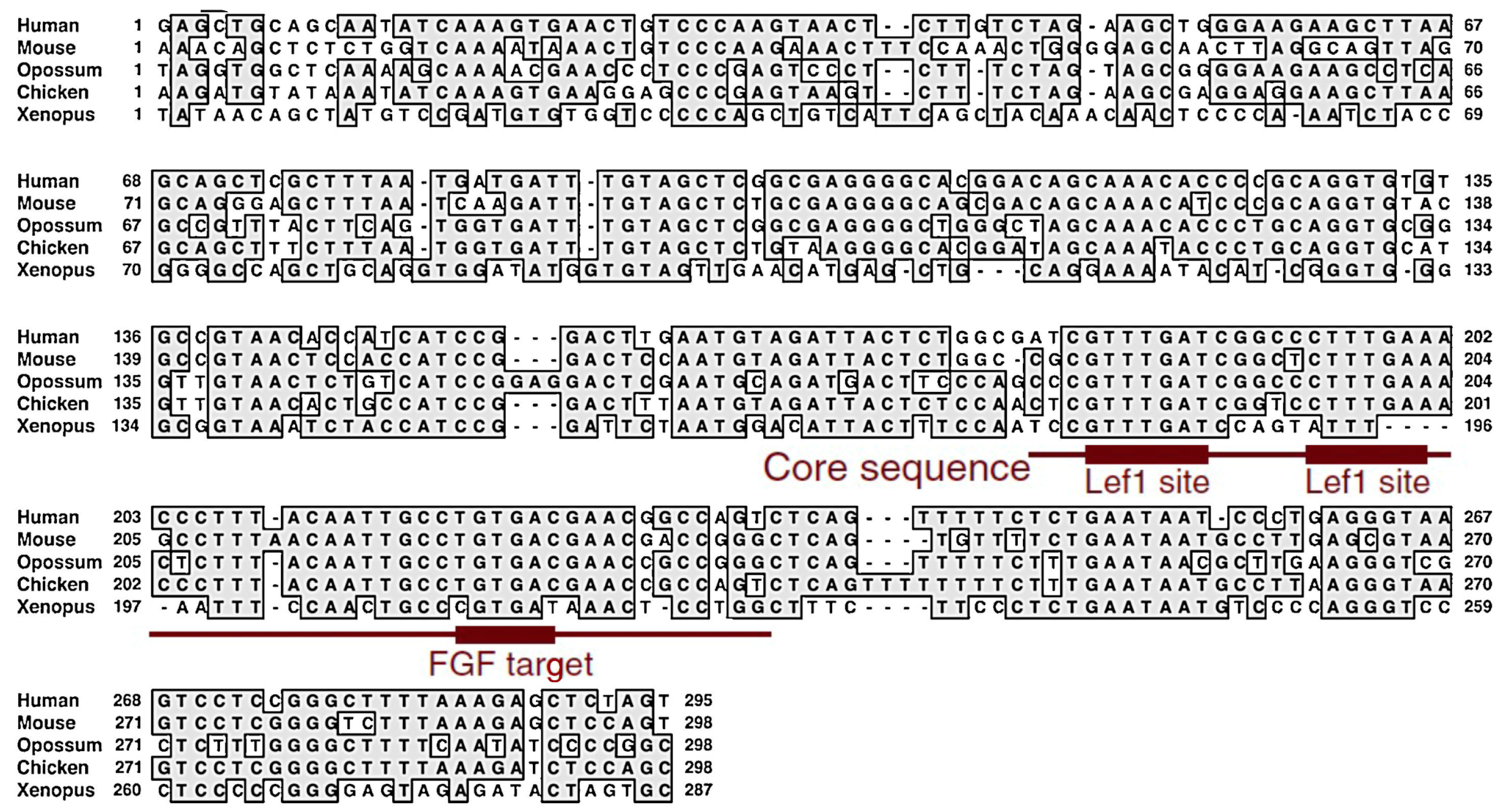
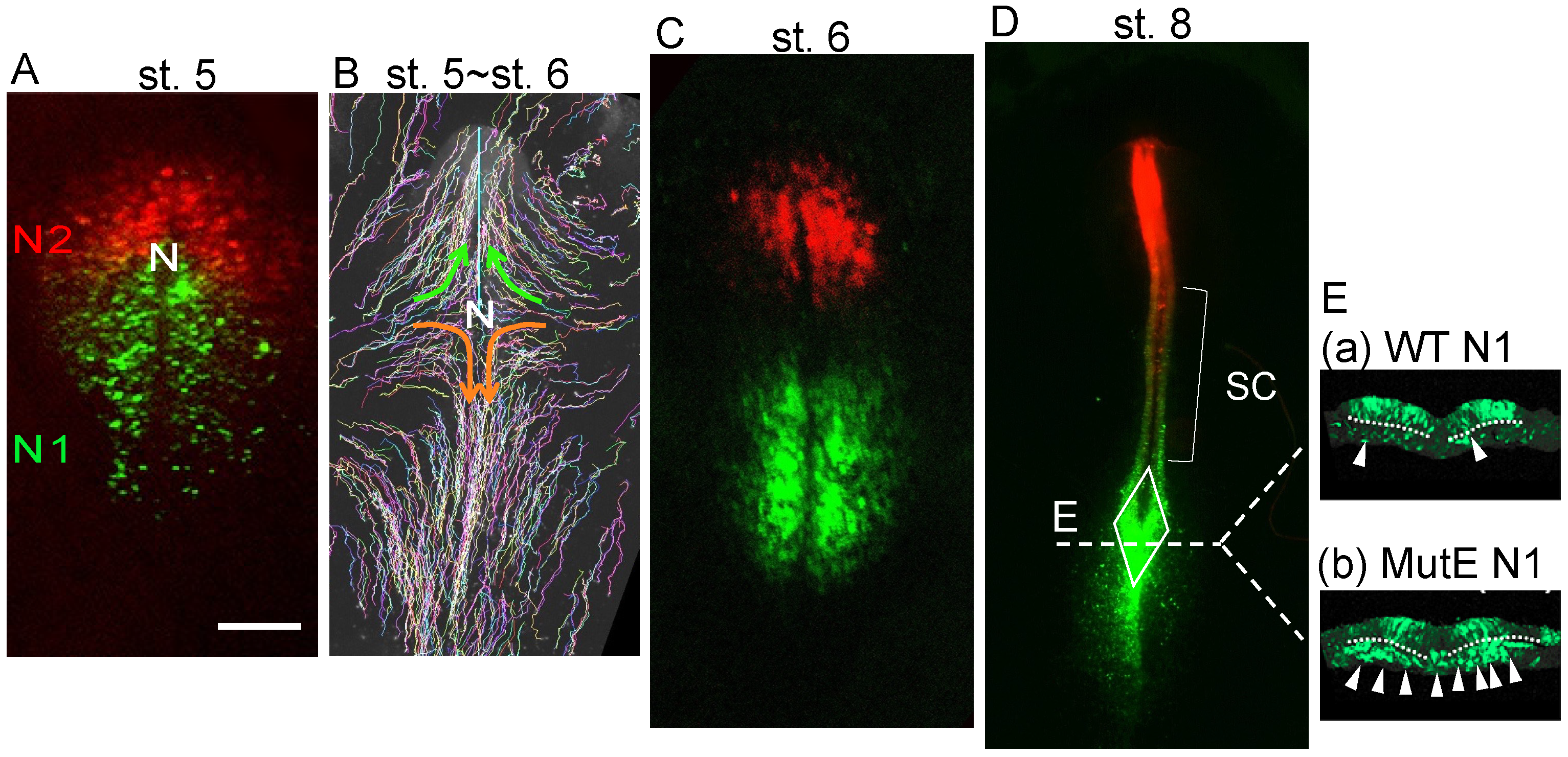
3. The Development of Two Distinct Neural Progenitors in the Epiblast with the Activation of the Sox2 Enhancers N1 or N2, with N1 Marking NMPs
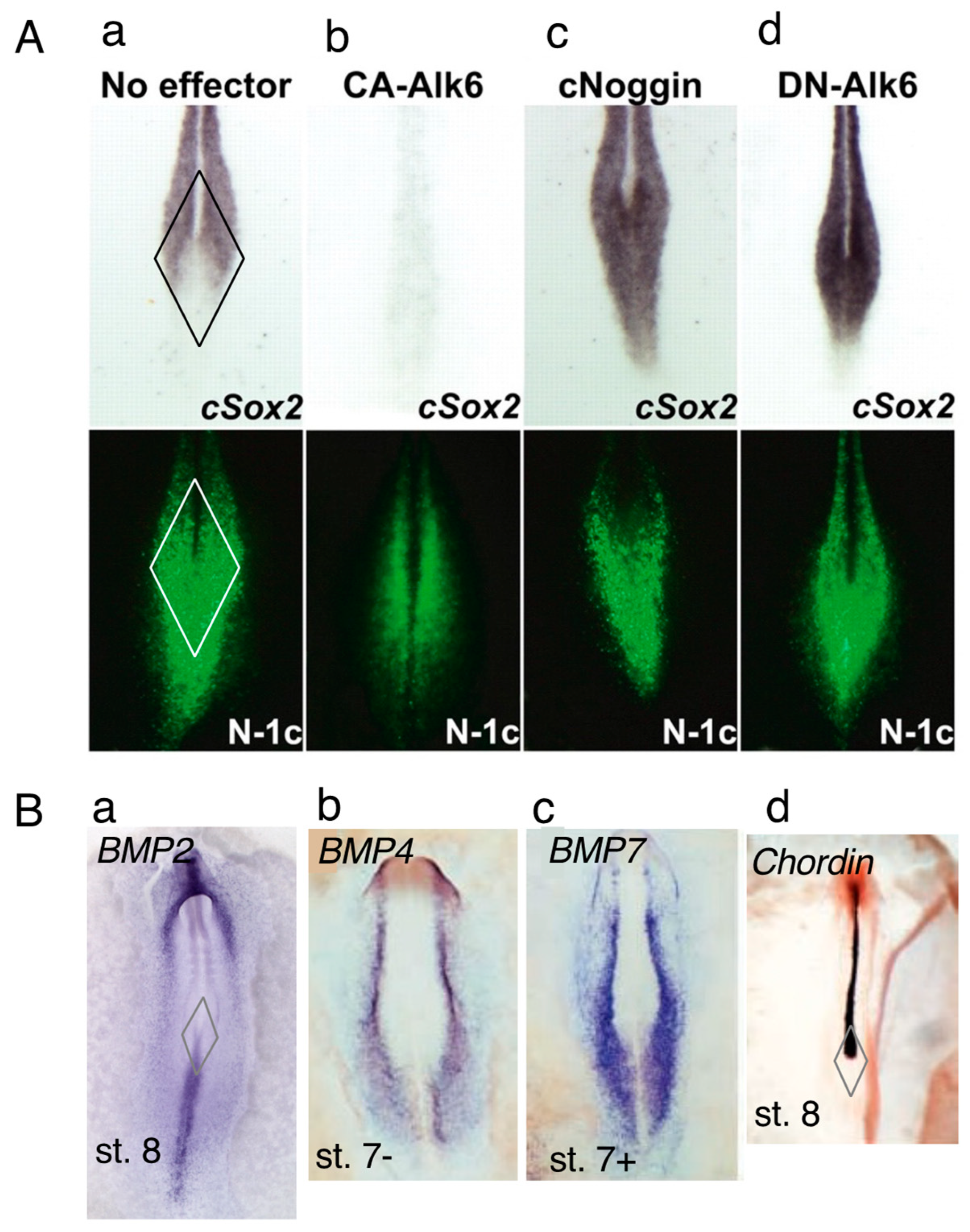
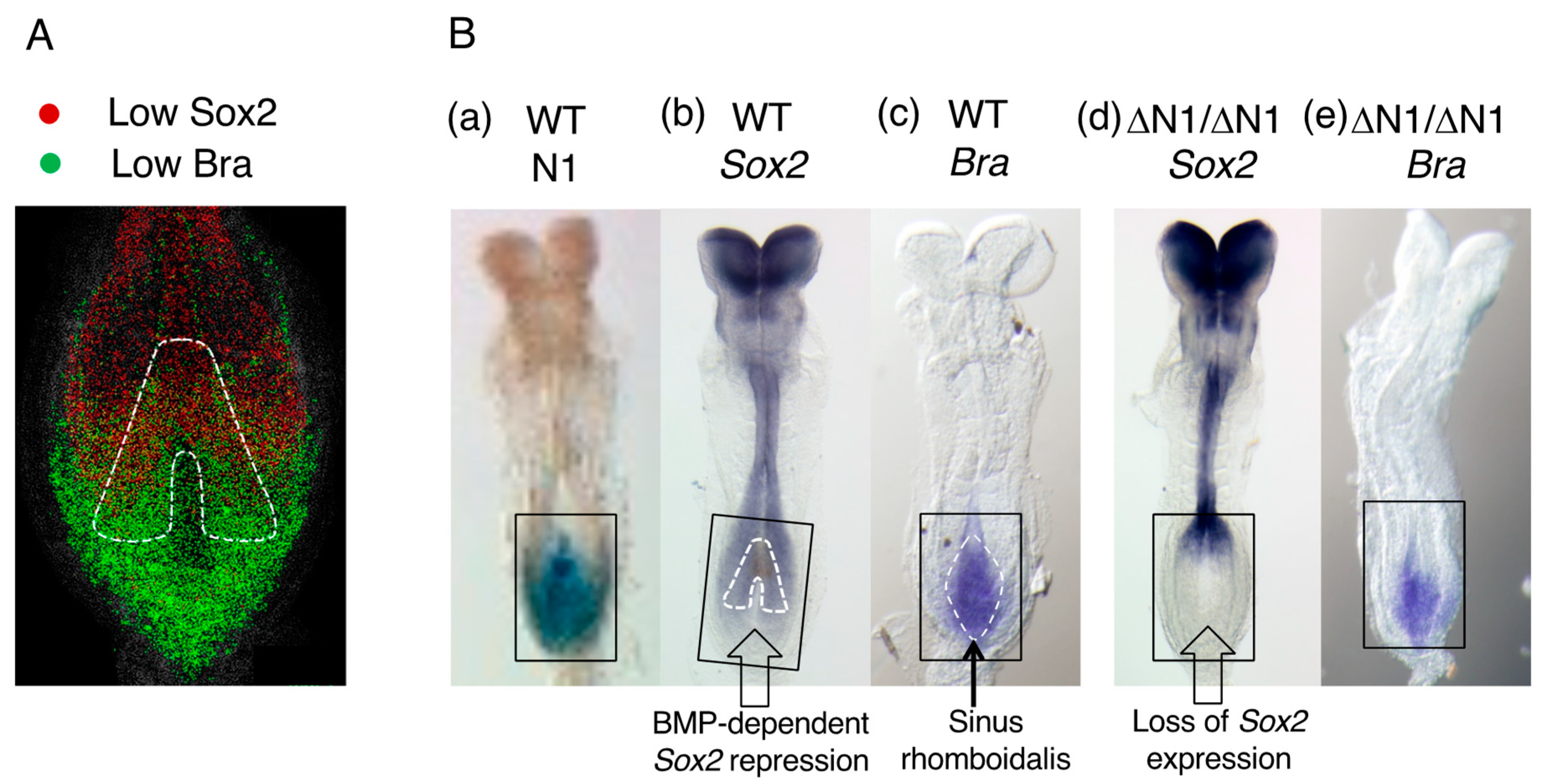
4. BMP-Dependent Sox2 Repression Regulates the Neural or Mesodermal Fates of NMPs
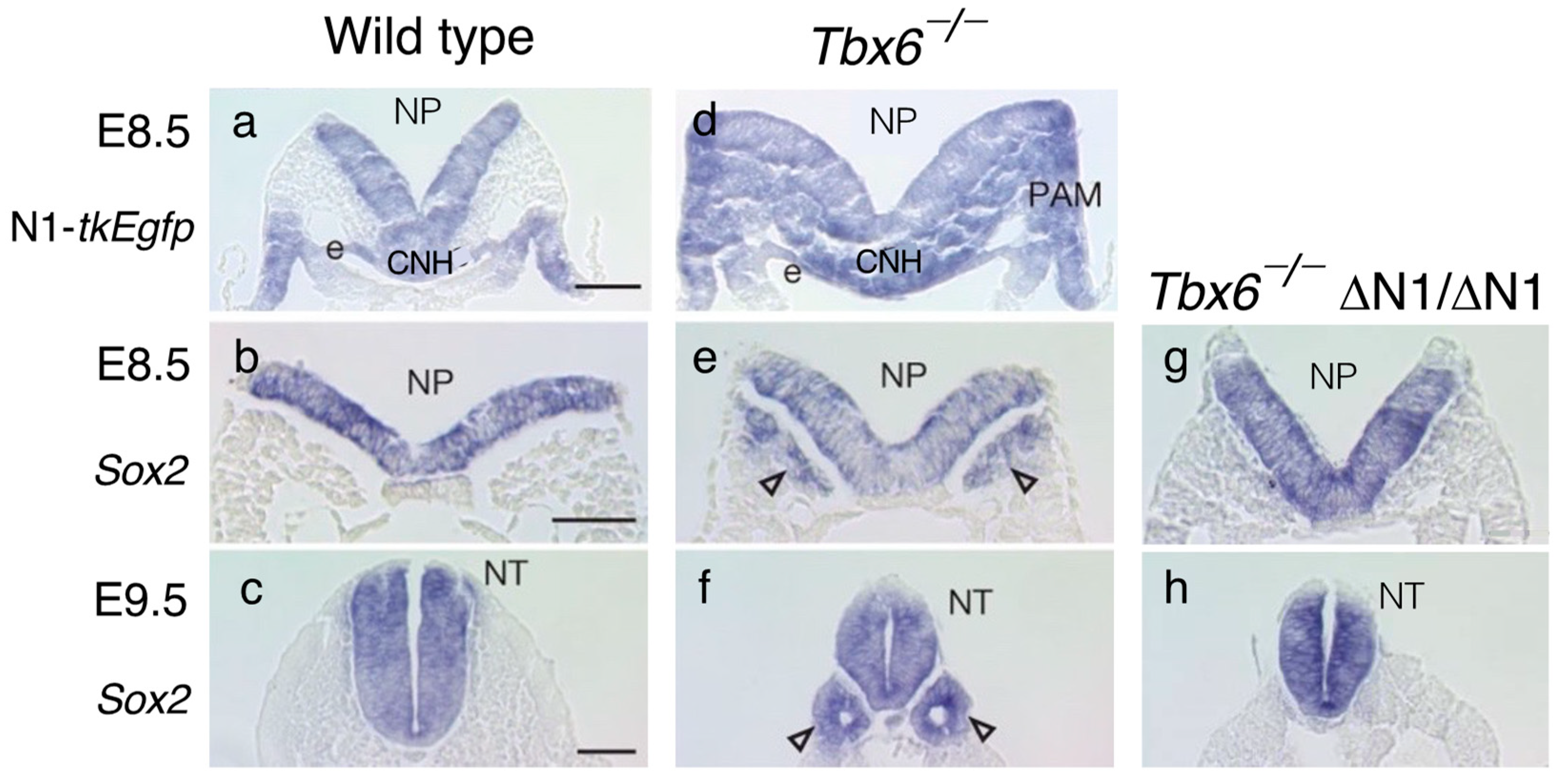
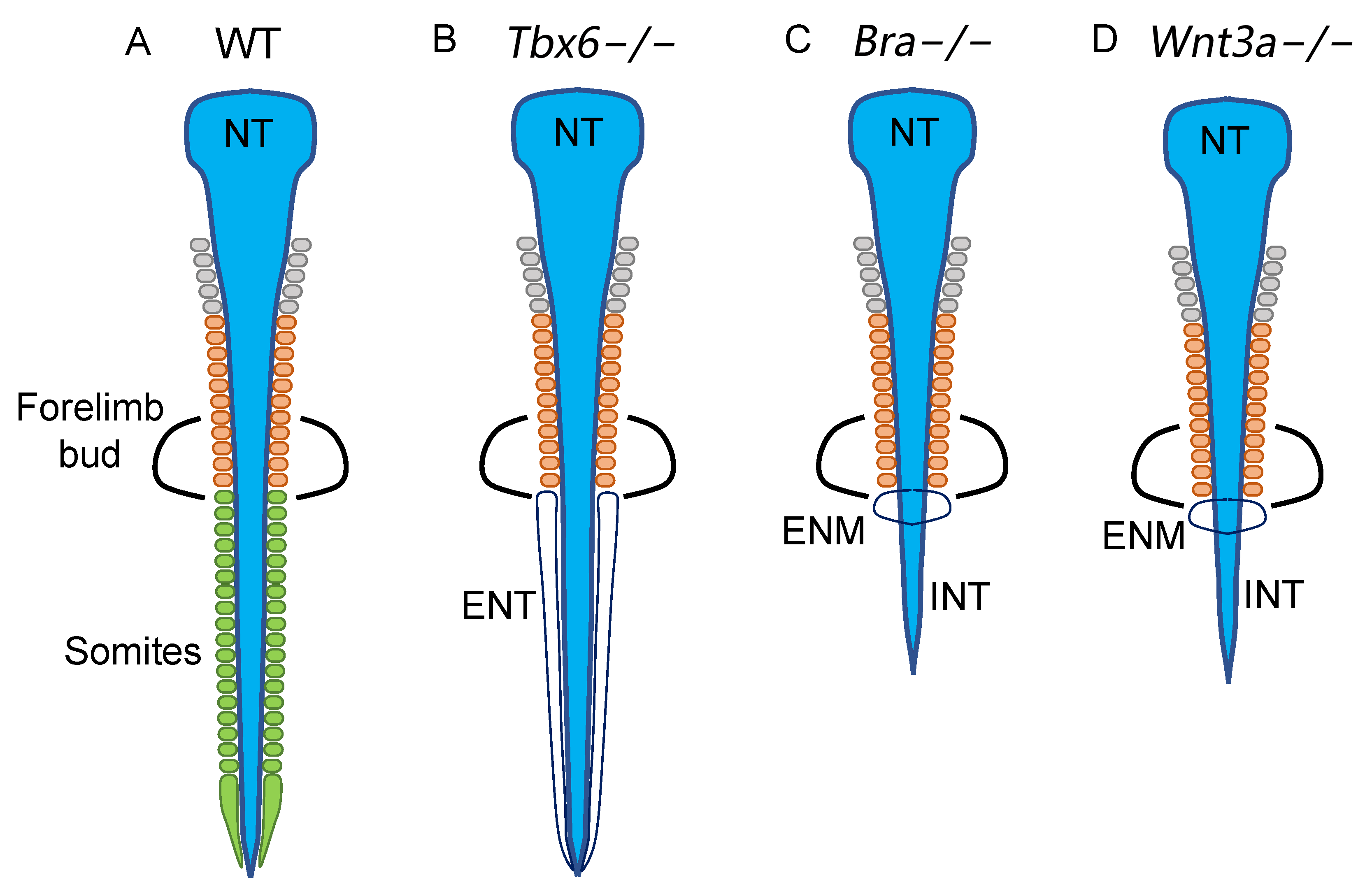
5. Tbx6 Represses N1 Enhancer Activity in the NMP-Derived Mesoderm and Inhibits Neural Development of the Paraxial Mesoderm
6. Nephric Mesenchyme Also Develops from NMPs
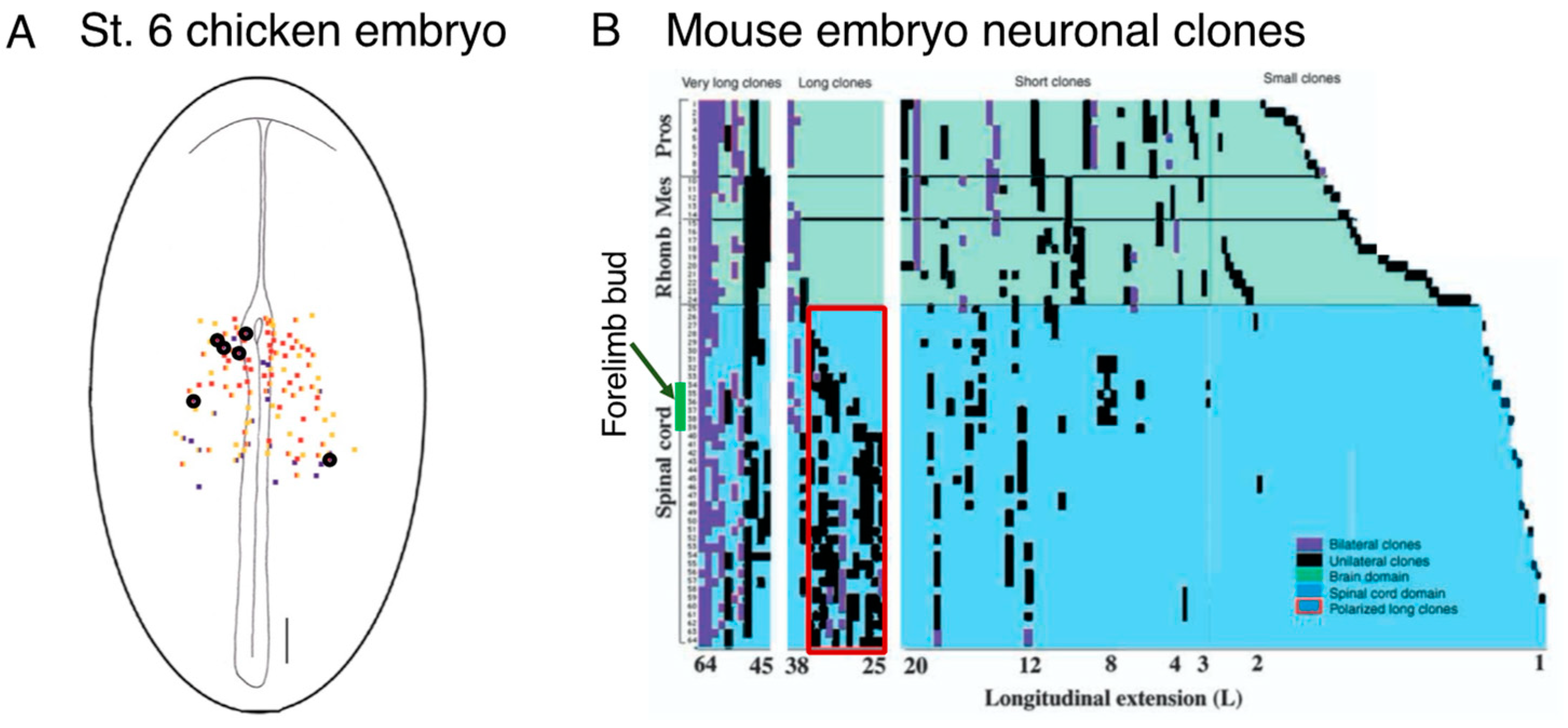
7. The Transition of Spinal Cord and Paraxial Mesoderm Sources from Tissue-Specific Progenitors to NMPs
8. Sox2 and Bra Coexpression Is Not a Hallmark of NMPs
9. NMP Pool Maintenance during Axial Elongation of the Embryo through the Supply of Canonical Wnt Signals
10. In Vitro Models of NMP Development
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Takemoto, T.; Uchikawa, M.; Kamachi, Y.; Kondoh, H. Convergence of Wnt and FGF signals in the genesis of posterior neural plate through activation of the Sox2 enhancer N-1. Development 2006, 133, 297–306. [Google Scholar] [CrossRef]
- Tzouanacou, E.; Wegener, A.; Wymeersch, F.J.; Wilson, V.; Nicolas, J.F. Redefining the progression of lineage segregations during mammalian embryogenesis by clonal analysis. Dev. Cell 2009, 17, 365–376. [Google Scholar] [CrossRef]
- Mathis, L.; Bonnerot, C.; Puelles, L.; Nicolas, J.F. Retrospective clonal analysis of the cerebellum using genetic laacZ/lacZ mouse mosaics. Development 1997, 124, 4089–4104. [Google Scholar] [CrossRef]
- Zambrowicz, B.P.; Imamoto, A.; Fiering, S.; Herzenberg, L.A.; Kerr, W.G.; Soriano, P. Disruption of overlapping transcripts in the ROSA beta geo 26 gene trap strain leads to widespread expression of beta-galactosidase in mouse embryos and hematopoietic cells. Proc. Natl. Acad. Sci. USA 1997, 94, 3789–3794. [Google Scholar] [CrossRef]
- Chapman, D.L.; Papaioannou, V.E. Three neural tubes in mouse embryos with mutations in the T-box gene Tbx6. Nature 1998, 391, 695–697. [Google Scholar] [CrossRef]
- Takemoto, T.; Uchikawa, M.; Yoshida, M.; Bell, D.M.; Lovell-Badge, R.; Papaioannou, V.E.; Kondoh, H. Tbx6-dependent Sox2 regulation determines neural or mesodermal fate in axial stem cells. Nature 2011, 470, 394–398. [Google Scholar] [CrossRef]
- Martin, B.L.; Kimelman, D. Canonical Wnt signaling dynamically controls multiple stem cell fate decisions during vertebrate body formation. Dev. Cell 2012, 22, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Kimelman, D. Hox13 genes are required for mesoderm formation and axis elongation during early zebrafish development. Development 2020, 147, dev185298. [Google Scholar] [CrossRef] [PubMed]
- Serrano Nájera, G.; Weijer, C.J. Cellular processes driving gastrulation in the avian embryo. Mech. Dev. 2020, 163, 103624. [Google Scholar] [CrossRef] [PubMed]
- Voiculescu, O. Movements of chick gastrulation. Curr. Top. Dev. Biol. 2020, 136, 409–428. [Google Scholar] [CrossRef] [PubMed]
- Yoshihi, K.; Kato, K.; Iida, H.; Teramoto, M.; Kawamura, A.; Watanabe, Y.; Nunome, M.; Nakano, M.; Matsuda, Y.; Sato, Y.; et al. Live imaging of avian epiblast and anterior mesendoderm grafting reveals the complexity of cell dynamics during early brain development. Development 2022, 149, dev199999. [Google Scholar] [CrossRef]
- Huss, D.; Benazeraf, B.; Wallingford, A.; Filla, M.; Yang, J.; Fraser, S.E.; Lansford, R. A transgenic quail model that enables dynamic imaging of amniote embryogenesis. Development 2015, 142, 2850–2859. [Google Scholar] [CrossRef] [PubMed]
- Catala, M.; Teillet, M.A.; De Robertis, E.M.; Le Douarin, M.L. A spinal cord fate map in the avian embryo: While regressing, Hensen’s node lays down the notochord and floor plate thus joining the spinal cord lateral walls. Development 1996, 122, 2599–2610. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Epstein, D.J. Distinct regulators of Shh transcription in the floor plate and notochord indicate separate origins for these tissues in the mouse node. Development 2003, 130, 3891–3902. [Google Scholar] [CrossRef] [PubMed]
- Cambray, N.; Wilson, V. Two distinct sources for a population of maturing axial progenitors. Development 2007, 134, 2829–2840. [Google Scholar] [CrossRef] [PubMed]
- Uchikawa, M.; Kondoh, H. Regulation of Sox2 via Many Enhancers of Distinct Specificities. In Sox2: Biology and Role in Development and Disease; Kondoh, H., Lovell-Badge, R., Eds.; Academic Press: San Diego, CA, USA, 2016; Chapter 7; pp. 107–129. [Google Scholar]
- Chapman, S.C.; Brown, R.; Lees, L.; Schoenwolf, G.C.; Lumsden, A. Expression analysis of chick Wnt and frizzled genes and selected inhibitors in early chick patterning. Dev. Dyn. 2004, 229, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Danielian, P.S.; Fritzsch, B.; McMahon, A.P. Evidence that FGF8 signalling from the midbrain-hindbrain junction regulates growth and polarity in the developing midbrain. Development 1997, 124, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Karabagli, H.; Karabagli, P.; Ladher, R.K.; Schoenwolf, G.C. Comparison of the expression patterns of several fibroblast growth factors during chick gastrulation and neurulation. Anat. Embryol. 2002, 205, 365–370. [Google Scholar] [CrossRef]
- Stolte, D.; Huang, R.; Christ, B. Spatial and temporal pattern of Fgf-8 expression during chicken development. Anat. Embryol. 2002, 205, 1–6. [Google Scholar] [CrossRef]
- Kamachi, Y.; Iwafuchi, M.; Okuda, Y.; Takemoto, T.; Uchikawa, M.; Kondoh, H. Evolution of non-coding regulatory sequences involved in the developmental process: Reflection of differential employment of paralogous genes as highlighted by Sox2 and group B1 Sox genes. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 55–68. [Google Scholar] [CrossRef]
- Nakamura, K.; Watanabe, Y.; Boitet, C.; Satake, S.; Iida, H.; Yoshihi, K.; Ishii, Y.; Kato, K.; Kondoh, H. Wnt signal-dependent antero-posterior specification of early-stage CNS primordia modeled in EpiSC-derived neural stem cells. Front. Cell Dev. Biol. 2024, 11, 1260528. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, H.; Luo, W.; Tarusawa, E.; Saito, Y.M.; Sato, T.; Yoshimura, Y.; Itohara, S.; Iwasato, T. NMDAR-regulated dynamics of layer 4 neuronal dendrites during thalamocortical reorganization in neonates. Neuron 2014, 82, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Storey, K.G. A region of the vertebrate neural plate in which neighbouring cells can adopt neural or epidermal fates. Curr. Biol. 2000, 10, 869–872. [Google Scholar] [CrossRef]
- Antin, P.B.; Yatskievych, T.A.; Davey, S.; Darnell, D.K. GEISHA: An evolving gene expression resource for the chicken embryo. Nucleic Acids Res. 2014, 42, D933–D937. [Google Scholar] [CrossRef]
- Streit, A.; Lee, K.J.; Woo, I.; Roberts, C.; Jessell, T.M.; Stern, C.D. Chordin regulates primitive streak development and the stability of induced neural cells, but is not sufficient for neural induction in the chick embryo. Development 1998, 125, 507–519. [Google Scholar] [CrossRef]
- Streit, A.; Stern, C.D. Establishment and maintenance of the border of the neural plate in the chick: Involvement of FGF and BMP activity. Mech. Dev. 1999, 82, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Iimura, T.; Pourquié, O. Collinear activation of Hoxb genes during gastrulation is linked to mesoderm cell ingression. Nature 2006, 442, 568–571. [Google Scholar] [CrossRef]
- Iimura, T.; Yang, X.; Weijer, C.J.; Pourquié, O. Dual mode of paraxial mesoderm formation during chick gastrulation. Proc. Natl. Acad. Sci. USA 2007, 104, 2744–2749. [Google Scholar] [CrossRef]
- Knezevic, V.; De Santo, R.; Mackem, S. Two novel chick T-box genes related to mouse Brachyury are expressed in different, non-overlapping mesodermal domains during gastrulation. Development 1997, 124, 411–419. [Google Scholar] [CrossRef]
- Kondoh, H. Gastrulation: Its principles and variations. In Molecular Basis of Developmental and Stem Cell Regulation; Springer-Nature: Berlin/Heidelberg, Germany, 2024; Chapter 3; ISBN 978-3-031-39026-5. [Google Scholar]
- Dressler, G.R. Advances in early kidney specification, development and patterning. Development 2009, 136, 3863–3874. [Google Scholar] [CrossRef]
- Hayashi, S.; Suzuki, H.; Takemoto, T. The nephric mesenchyme lineage of intermediate mesoderm is derived from Tbx6-expressing derivatives of neuro-mesodermal progenitors via BMP-dependent Osr1 function. Dev. Biol. 2021, 478, 155–162. [Google Scholar] [CrossRef]
- Kanda, S.; Tanigawa, S.; Ohmori, T.; Taguchi, A.; Kudo, K.; Suzuki, Y.; Sato, Y.; Hino, S.; Sander, M.; Perantoni, A.O.; et al. Sall1 maintains nephron progenitors and nascent nephrons by acting as both an activator and a repressor. J. Am. Soc. Nephrol. 2014, 25, 2584–2595. [Google Scholar] [CrossRef]
- Taguchi, A.; Nishinakamura, R. Nephron reconstitution from pluripotent stem cells. Kidney Int. 2015, 87, 894–900. [Google Scholar] [CrossRef]
- Wilm, B.; James, R.G.; Schultheiss, T.M.; Hogan, B.L. The forkhead genes, Foxc1 and Foxc2, regulate paraxial versus intermediate mesoderm cell fate. Dev. Biol. 2004, 271, 176–189. [Google Scholar] [CrossRef]
- Mathis, L.; Nicolas, J.F. Different clonal dispersion in the rostral and caudal mouse central nervous system. Development 2000, 127, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- Chesley, P. Development of the short-tailed mutation in the house mouse. J. Exp. Zool. 1935, 70, 429–459. [Google Scholar] [CrossRef]
- Gruneberg, H. Genetical studies on the skeleton of the mouse. XXIII. The development of brachyury and anury. J. Embryol. Exp. Morphol. 1958, 6, 424–443. [Google Scholar] [PubMed]
- Takada, S.; Stark, K.L.; Shea, M.J.; Vassileva, G.; McMahon, J.A.; McMahon, A.P. Wnt-3a regulates somite and tailbud formation in the mouse embryo. Genes Dev. 1994, 8, 174–189. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, Y.; Fujimori, T.; McMahon, A.P.; Takada, S. Evidence that absence of Wnt-3a signaling promotes neuralization instead of paraxial mesoderm development in the mouse. Dev Biol. 1997, 183, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.P.; Takada, S.; Yoshikawa, Y.; Wu, N.; McMahon, A.P. T (Brachyury) is a direct target of Wnt3a during paraxial mesoderm specification. Genes Dev. 1999, 13, 3185–3190. [Google Scholar] [CrossRef] [PubMed]
- Henrique, D.; Abranches, E.; Verrier, L.; Storey, K.G. Neuromesodermal progenitors and the making of the spinal cord. Development 2015, 142, 2864–2875. [Google Scholar] [CrossRef]
- Delfino-Machín, M.; Lunn, J.S.; Breitkreuz, D.N.; Akai, J.; Storey, K.G. Specification and maintenance of the spinal cord stem zone. Development 2005, 132, 4273–4283. [Google Scholar] [CrossRef]
- Wymeersch, F.J.; Huang, Y.; Blin, G.; Cambray, N.; Wilkie, R.; Wong, F.C.; Wilson, V. Position-dependent plasticity of distinct progenitor types in the primitive streak. eLife 2016, 5, e10042. [Google Scholar] [CrossRef]
- Rodrigo Albors, A.; Halley, P.A.; Storey, K.G. Lineage tracing of axial progenitors using Nkx1-2CreERT2 mice defines their trunk and tail contributions. Development 2018, 145, dev164319. [Google Scholar] [CrossRef] [PubMed]
- Guillot, C.; Djeffal, Y.; Michaut, A.; Rabe, B.; Pourquié, O. Dynamics of primitive streak regression controls the fate of neuromesodermal progenitors in the chicken embryo. eLife 2021, 10, e64819. [Google Scholar] [CrossRef] [PubMed]
- Gouti, M.; Delile, J.; Stamataki, D.; Wymeersch, F.J.; Huang, Y.; Kleinjung, J.; Wilson, V.; Briscoe, J. A Gene Regulatory Network Balances Neural and Mesoderm Specification during Vertebrate Trunk Development. Dev. Cell 2017, 41, 243–261.e7. [Google Scholar] [CrossRef] [PubMed]
- Koch, F.; Scholze, M.; Wittler, L.; Schifferl, D.; Sudheer, S.; Grote, P.; Timmermann, B.; Macura, K.; Herrmann, B.G. Antagonistic Activities of Sox2 and Brachyury Control the Fate Choice of Neuro-Mesodermal Progenitors. Dev. Cell 2017, 42, 514–526.e7. [Google Scholar] [CrossRef] [PubMed]
- Guibentif, C.; Griffiths, J.A.; Imaz-Rosshandler, I.; Ghazanfar, S.; Nichols, J.; Wilson, V.; Göttgens, B.; Marioni, J.C. Diverse Routes toward Early Somites in the Mouse Embryo. Dev. Cell 2021, 56, 141–153.e6. [Google Scholar] [CrossRef] [PubMed]
- Schoenwolf, G.C. Histological and ultrastructural observations of tail bud formation in the chick embryo. Anat. Rec. 1979, 193, 131–147. [Google Scholar] [CrossRef]
- Kawachi, T.; Shimokita, E.; Kudo, R.; Tadokoro, R.; Takahashi, Y. Neural-fated self-renewing cells regulated by Sox2 during secondary neurulation in chicken tail bud. Dev. Biol. 2020, 461, 160–171. [Google Scholar] [CrossRef]
- Wilson, V.; Olivera-Martinez, I.; Storey, K.G. Stem cells, signals and vertebrate body axis extension. Development 2009, 136, 1591–1604. [Google Scholar] [CrossRef]
- Hatakeyama, Y.; Saito, N.; Mii, Y.; Takada, R.; Shinozuka, T.; Takemoto, T.; Naoki, H.; Takada, S. Intercellular exchange of Wnt ligands reduces cell population heterogeneity during embryogenesis. Nat. Commun. 2023, 14, 1924. [Google Scholar] [CrossRef]
- Takemoto, T.; Abe, T.; Kiyonari, H.; Nakao, K.; Furuta, Y.; Suzuki, H.; Takada, S.; Fujimori, T.; Kondoh, H. R26-WntVis reporter mice showing graded response to Wnt signal levels. Genes Cells 2016, 21, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Tahara, N.; Kawakami, H.; Chen, K.Q.; Anderson, A.; Yamashita Peterson, M.; Gong, W.; Shah, P.; Hayashi, S.; Nishinakamura, R.; Nakagawa, Y.; et al. Sall4 regulates neuromesodermal progenitors and their descendants during body elongation in mouse embryos. Development 2019, 146, dev177659. [Google Scholar] [CrossRef] [PubMed]
- Pappas, M.P.; Kawakami, H.; Corcoran, D.; Chen, K.Q.; Scott, E.P.; Wong, J.; Gearhart, M.D.; Nishinakamura, R.; Nakagawa, Y.; Kawakami, Y. Sall4 regulates posterior trunk mesoderm development by promoting mesodermal gene expression and repressing neural genes in the mesoderm. Development 2024, 151, dev202649. [Google Scholar] [CrossRef] [PubMed]
- Edri, S.; Hayward, P.; Jawaid, W.; Martinez Arias, A. Neuro-mesodermal progenitors (NMPs): A comparative study between pluripotent stem cells and embryo-derived populations. Development 2019, 146, dev180190. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.A.; Hayward, P.C.; Baillie-Johnson, P.; Rue, P.; Broome, R.; Faunes, F.; Martinez Arias, A. Wnt/beta-catenin and FGF signalling direct the specification and maintenance of a neuromesodermal axial progenitor in ensembles of mouse embryonic stem cells. Development 2014, 141, 4243–4253. [Google Scholar] [CrossRef] [PubMed]
- Tsakiridis, A.; Wilson, V. Assessing the bipotency of in vitro-derived neuromesodermal progenitors. F1000Research 2015, 4, 100. [Google Scholar] [CrossRef] [PubMed]
- Edri, S.; Hayward, P.; Baillie-Johnson, P.; Steventon, B.J.; Martinez Arias, A. An epiblast stem cell-derived multipotent progenitor population for axial extension. Development 2019, 146, dev168187. [Google Scholar] [CrossRef] [PubMed]
- Denham, M.; Hasegawa, K.; Menheniott, T.; Rollo, B.; Zhang, D.; Hough, S.; Alshawaf, A.; Febbraro, F.; Ighaniyan, S.; Leung, J.; et al. Multipotent caudal neural progenitors derived from human pluripotent stem cells that give rise to lineages of the central and peripheral nervous system. Stem Cells 2015, 33, 1759–1770. [Google Scholar] [CrossRef]
- Verrier, L.; Davidson, L.; Gierliński, M.; Dady, A.; Storey, K.G. Neural differentiation, selection and transcriptomic profiling of human neuromesodermal progenitor-like cells in vitro. Development 2018, 145, dev166215. [Google Scholar] [CrossRef] [PubMed]
- Cooper, F.; Gentsch, G.E.; Mitter, R.; Bouissou, C.; Healy, L.E.; Rodriguez, A.H.; Smith, J.C.; Bernardo, A.S. Rostrocaudal patterning and neural crest differentiation of human pre-neural spinal cord progenitors in vitro. Stem Cell Reports 2022, 17, 894–910. [Google Scholar] [CrossRef] [PubMed]
- Cooper, F.; Tsakiridis, A. Towards clinical applications of in vitro-derived axial progenitors. Dev Biol. 2022, 489, 110–117. [Google Scholar] [CrossRef]
- Fan, Y.; Hackland, J.; Baggiolini, A.; Hung, L.Y.; Zhao, H.; Zumbo, P.; Oberst, P.; Minotti, A.P.; Hergenreder, E.; Najjar, S.; et al. hPSC-derived sacral neural crest enables rescue in a severe model of Hirschsprung’s disease. Cell Stem Cell. 2023, 30, 264–282.e9. [Google Scholar] [CrossRef]
- Iyer, N.R.; Shin, J.; Cuskey, S.; Tian, Y.; Nicol, N.R.; Doersch, T.E.; Seipel, F.; McCalla, S.G.; Roy, S.; Ashton, R.S. Modular derivation of diverse, regionally discrete human posterior CNS neurons enables discovery of transcriptomic patterns. Sci. Adv. 2022, 8, eabn7430. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondoh, H.; Takemoto, T. The Origin and Regulation of Neuromesodermal Progenitors (NMPs) in Embryos. Cells 2024, 13, 549. https://doi.org/10.3390/cells13060549
Kondoh H, Takemoto T. The Origin and Regulation of Neuromesodermal Progenitors (NMPs) in Embryos. Cells. 2024; 13(6):549. https://doi.org/10.3390/cells13060549
Chicago/Turabian StyleKondoh, Hisato, and Tatsuya Takemoto. 2024. "The Origin and Regulation of Neuromesodermal Progenitors (NMPs) in Embryos" Cells 13, no. 6: 549. https://doi.org/10.3390/cells13060549
APA StyleKondoh, H., & Takemoto, T. (2024). The Origin and Regulation of Neuromesodermal Progenitors (NMPs) in Embryos. Cells, 13(6), 549. https://doi.org/10.3390/cells13060549